缺氮脅迫下含 Na+葉面肥對花生生長的抑制及補氮后的恢復效應
2016-12-20 13:44:39于天一鄭永美吳正鋒陳殿緒孫學武王才斌鄭亞萍
植物營養與肥料學報 2016年6期
沈 浦,馮 昊,羅 盛,于天一,鄭永美,吳正鋒,陳殿緒,孫學武,王才斌,鄭亞萍
(山東省花生研究所,山東青島 266100)
缺氮脅迫下含 Na+葉面肥對花生生長的抑制及補氮后的恢復效應
沈 浦,馮 昊,羅 盛,于天一,鄭永美,吳正鋒,陳殿緒*,孫學武*,王才斌,鄭亞萍
(山東省花生研究所,山東青島 266100)
【目的】葉面肥中的鈉離子 (Na+) 大多為伴隨離子,其對作物的影響通常被忽視。實踐中,Na+對作物的作用包括促進和抑制兩方面,取決于作物營養生理狀況。研究 Na+對花生的抑制與缺氮 (N) 脅迫的關系,探索補施氮素對上述抑制作用的恢復效應,可為花生高效合理施肥提供理論依據。 【方法】盆栽試驗在山東省花生研究所人工氣候室內進行,設噴施肥料中含 Na+0、2.0、10.0 mg/pot 3 個水平,依次記為 T0、T1 和 T2。測定了花生葉片光合作用、莖葉及根干物質累積、根系形態特性和養分吸收,比較了缺氮脅迫 (T0、T1、T2) 和后期補施尿素態 N (T0-N、T1-N、T2-N) 條件下 Na+對花生影響的差異,并深入分析花生 Na 吸收與干物質累積和氮(N)、磷 (P)、鉀 (K) 養分吸收的相關關系。 【結果】1) 缺氮脅迫條件下,葉面噴施 Na+顯著抑制了花生的光合作用,凈光合速率由 CO211.4 μM/(m2·s) 下降到 CO26.7~7.5 μM/(m2·s)。莖葉和根干重 T0 (6.4 和 2.4 g/pot) > T1 (5.7 和 1.9 g/pot) > T2 (5.3 和 1.5 g/pot)。與 T0 相比,根總長度、總表面積和總體積 T1 處理下降了7.8%~10.5%,T2 處理顯著下降了 27.5%~31.8%。植株 Na 與 N、K 含量呈顯著負相關關系 (P < 0.05),花生Na 吸收量每增加 1.0 mg/pot,植株總的 N、K 吸收量分別下降 26.6 和 20.9 mg/pot。2) 后期補充噴施氮素條件下,施 Na+處理花生莖葉干物重比不施 Na+處理增加了 20.3%~25.8%。補施氮素后,施 Na+促進了莖葉對 N、P、K 的吸收,N 的吸收量增加了 9.2%~19.2%,P 增加了 20.0%~42.3%,K 增加了 21.4%~24.0%。相關性分析表明,花生植株總干物重及 P、K 含量在補施氮素后與 Na 含量呈顯著正相關關系 (P < 0.05)。 【結論】Na+對花生生長及養分吸收的影響與氮素營養有關。在缺氮脅迫下,Na+對花生的生長為抑制作用,抑制程度隨噴施量增加而加重,高濃度 Na+顯著抑制根莖葉干物質的累積和 N、K 養分的吸收。補施氮肥提高了花生植株對Na+的適應性,恢復了光合作用及養分吸收能力,消除了 Na 脅迫的危害作用,產生“N–Na”協同促進效應。
鈉脅迫;生長狀況;補充供氮;恢復效應
Na+常常被認為是造成作物鹽堿危害的主要離子,其可通過滲透脅迫引起營養元素的缺乏,易造成活性氧清除酶 (如 SOD、POD 等) 失活,置換細胞膜中的 Ca2+,引起膜脂、蛋白質、核酸等的降解,破壞細胞組分,進而影響植株生長發育[1–3]。但近年來研究表明,很多植株的生長需要一定量的 Na+以維持滲透壓、促進碳水化合物代謝等,且 Na+能夠提高一些作物的葉面積指數、增加光截獲能力,以及具有替代植株其他營養元素 (如 K+) 的作用[4–7]。前人研究已證實,Na+對作物的抑制作用與營養生理狀況有很大關系,而缺氮脅迫是其中一個重要的因素[8–9]。由于土壤供給能力、氮肥利用效率及品種等方面的差異,作物缺氮脅迫現象時有發生[10–11]。缺氮脅迫下作物生長發育一方面會受到顯著抑制,另一方面作物自身也會產生一系列生理生化的適應性變化[12–13]。由此可見,缺氮脅迫下 Na+對作物生長發育的影響需要研究予以證實。
花生是我國重要的油料作物、經濟作物,在國民生產中有重要的作用[14–15]。生產中,花生可通過肥料及土壤吸收一定量的 Na+,特別是使用葉面施肥過程中,一些含 Na+成分 (EDTA-Na2、氯化鈉、亞硫酸氫鈉、四水八硼酸鈉、復硝酚鈉、硝普鈉和葡萄糖酸鈉等) 隨營養元素一起噴施在葉面而進入植株體內[16–19]。對此,大多數研究僅關注該肥料中營養元素(如 N、P、K 等) 的吸收利用狀況,而 Na+對花生的真實影響卻鮮有報道。Na+對花生的作用可能與其施入量 (或濃度) 有關,且較高 Na+施入量會對花生生長造成不利影響[20–21]。這種不利影響可能首先表現在葉片受損、養分吸收受到抑制、光合產物下降,而地下部根系發育是否受到抑制還不明確。作者前期研究結果發現葉面噴施氮素能夠顯著改善花生的葉面積、SPAD 值及凈光合速率,這主要由于氮素是花生蛋白質和核酸的組成元素,又是光合作用所必需的葉綠素、各種代謝調控酶、激素的構成元素[22–23]。然而,葉面噴施 Na+是否會對花生產生抑制作用以及這種抑制能否通過補施氮素緩解,有待于深入研究。
本文利用人工氣候室盆栽試驗,研究了缺氮素脅迫下含不同濃度 Na+葉面肥對花生光合作用、干物質累積、根系形態及養分吸收的影響差異。同時,比較了補施氮素對各處理的作用效果,以探究葉面噴施氮肥能否消除或緩解 Na+的影響,以及對花生的光合作用和生長發育的恢復效應。本研究的開展可為花生高效合理施肥提供理論依據。
1 材料與方法
1.1 試驗材料
花生品種為山東省花生研究所培育的花育 20 號,該品種為小花生類型。試驗噴施液中鈉來源于氫氧化鈉,氮來源于尿素,磷、鉀來源于磷酸二氫鉀,三種化學試劑均為上海國藥集團生產。供試土壤采自于山東省花生研究所萊西試驗農場,土壤類型為棕壤,pH 值為 5.1、有機質 11.6 g/kg、速效磷 26.5 mg/kg、堿解氮 56.3 mg/kg、速效鉀 70.0 mg/kg。
1.2 試驗設計
盆栽試驗于 2014 年 11 月 27 日到 2015 年 2 月 2 日在山東省花生研究所人工氣候室開展,氣候室內晝夜溫度為 28℃/22℃、相對濕度 65%/70%。試驗盆缽直徑 15 cm、高 20 cm,每盆用土 2.1 kg,盆內土壤不施基肥。每盆播入 3 粒花生種,在齊苗后間苗,保留 2 棵長勢均勻的花生苗。盆栽試驗每 2 天澆水 1 次,按最大田間持水量的 75% 左右保持土壤含水量。
2014 年 12 月 22 日 (四葉期) 開始葉面噴施含氫氧化鈉及磷酸二氫鉀溶液,每周噴 1 次,共噴施 3次;試驗噴施的 Na 及 N、P、K 含量狀況見表 1。試驗處理 T0 噴施液的 pH 值為 5.0;T1 處理以 T0 為基礎,噴施液中加 Na 量為 2.0 mg/pot (每次噴施 Na 8.5 mL、0.08 mg/mL),噴施液 pH 值為 6.0;T2 處理加 Na 量為 10.0 mg/pot (每次噴施 Na 8.5 mL、0.4 mg/mL),噴施液 pH 值為 7.0。其中,T2 處理葉面肥中 Na+濃度依據常見含鈉葉面肥噴施狀況 (0.4‰) 設定。從 2015 年 1 月 12 日 (七葉期) 開始,一半盆栽噴施蒸餾水 (T0、T1 和 T2 處理),一半盆栽噴施氮素 (T0-N、T1-N 和 T2-N 處理),每周 1 次,共噴施 3次,各試驗處理均重復 3 次。

表1 處理編號及葉面噴施元素量 (mg/pot)Table 1 Treatment code and element for spraying solution
1.3 取樣與測定
2015 年 2 月 2 日先利用 CIRAS-3 光合儀 (PP Systems 公司,美國) 測定花生葉片凈光合速率。然后取整株花生,將其分為根部和莖葉部。根部用WinRHIZO 根系分析系統 (Regent Instruments 公司,加拿大) 測定根長度、根表面積、根體積等根系形態參數[24]。植株根和莖葉烘干后,測定干物重;然后樣品粉碎磨細,用 H2SO4–H2O2進行消化,依次測定N、P、K 和 Na 含量[25]。
1.4 數據處理
采用 Sigmaplot10.0 作圖,花生干物重、根系形態特性、NPK 和 Na 含量用 SAS 8.0 軟件進行各處理之間差異顯著性分析 (ANOVA) 和各指標之間相關顯著性分析。
2 結果與分析
2.1 花生地上部干物質累積及凈光合速率變化
缺氮脅迫下,噴施高 Na+葉面肥可顯著抑制花生凈光合速率及干物質累積 (圖 1)。T0 處理凈光合速率為 CO211.4 μM/(m2·s),噴施 Na+葉面肥顯著下降到 CO26.7~7.5 μM/(m2·s)(T1 和 T2 處理)。花生地上部莖葉干物重隨著 Na+增加而逐漸下降,T0、T1和 T2 處理分別為 6.4、5.7 和 5.3 g/pot。然而,補施氮素條件下,加入 Na+處理 T1-N 和 T2-N (8.4 和 8.1 g/pot) 地上部莖葉干物重卻顯著高于不加 Na+處理T0-N (6.7 g/pot)。同時,凈光合速率各處理沒有顯著差異,且以 T2-N 較高。在缺氮脅迫條件下,隨著葉面肥 Na+增加,花生干物質重量受到顯著抑制,而后期補充噴施氮素可緩解 Na+的抑制,甚至表現出促進干物質累積的作用。
2.2 花生根重及形態變化
缺氮脅迫下,花生根重及形態對葉面肥中 Na+的響應非常敏感 (圖 2)。根干重由 T0 的 2.4 g/pot 下降到 T1 的 1.9 g/pot 和 T2 的 1.5 g/pot。補施氮素后,3 個處理根干重 T1-N 高于 T0-N 和 T2-N,T0-N降低到 1.7 g/pot,而 T1-N 處理增加到 2.1 g/pot,T2-N處理維持在 1.5 g/pot。
花生根系形態也有相同的變化特征 (圖 2)。在缺氮脅迫條件下,葉面肥加入 Na+后,根總長度、根總表面積和根總體積都呈現下降趨勢,T1 比 T0 分別下降 7.8%~10.5%,T2 比 T0 分別顯著下降了 27.5%~31.8%。補施氮素后,T0-N 根系形態受到抑制,根總長度、根總表面積和根總體積仍高于 T2-N 處理11.3%~21.4%,卻比 T1-N 處理下降了 14.2%~21.0%。
三個根系形態指標總量對葉面肥 Na+的響應雖然相似,但從不同直徑根系來看,在缺氮脅迫條件下,高 Na+(T2 處理) 對根長的抑制從細根 (< 0.5 mm)開始,直徑超過 1.5 mm 根系受其抑制作用較弱;而根表面積及根體積則從粗根 ( > 1.5 mm) 開始,其受高 Na+的抑制越來越大 (圖 3a1、圖 3b1、圖 3c1)。補施氮素下,低 Na+(T1-N) 對根長的恢復也從細根(< 0.5 mm) 開始,而根表面積和根體積的恢復則從粗根 ( > 1.5mm) 開始 (圖 3a2、圖 3b2、圖 3c2)。
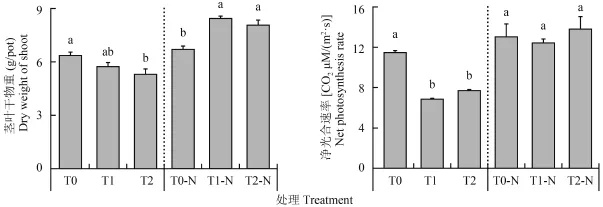
圖1 花生莖葉干重及凈光合速率變化Fig. 1 Changes in dry weight of shoot and net photosynthesis rate

圖2 花生根干重及總根系形態變化Fig. 2 Changes in dry weight of root and root morphological peanut
2.3 花生鈉吸收對干物質累積及氮、磷、鉀營養的影響
葉面噴施 Na+顯著提高了莖葉對鈉的吸收,缺氮脅迫下高 Na+處理 (T2) 莖葉鈉吸收量顯著高于 T1和 T0 處理 (圖 4),補施氮素下植株鈉含量表現為:T2-N > T1-N、T0-N。缺 N 脅迫下 Na+對莖葉 N 的吸收也產生了顯著影響,各處理莖葉氮素吸收量分別為 T0 (144.1 mg/pot) > T1 (132.0 mg/pot) > T2 (115.4 mg/pot)。補施氮素后,三個處理莖葉氮吸收量都顯著增加,而處理間以 T1-N (251.1 mg/pot) 最高,T2-N (230.0 mg/pot) 次之,T0-N (210.7 mg/pot) 最低。對于莖葉磷、鉀,缺氮脅迫下 T1 和 T2 處理與 T0 處理相比有下降趨勢,但沒有達到顯著差異水平。然而,補施氮素下施 Na+處理尤其高 Na+處理顯著高于不噴 Na+處理,T2-N 處理比 T0 處理磷吸收量和鉀吸收量分別顯著增加 42.3% 和 24.0%。

圖3 花生根系在不同直徑范圍的累積動態變化Fig. 3 Dynamics of root accumulations within different diameter range
在缺氮脅迫條件下,花生根系對 Na 的吸收量在噴施 Na+后略有增加,但未達到顯著差異;補施氮素后,各處理根的 Na 吸收量都下降至 0.42 mg/pot左右。根系對 N、P、K 的吸收量與根干重相似,隨著噴施 Na+濃度增加呈下降趨勢,T0 處理根的 N、P、K 吸收量大于 T1 和 T2 處理。補施 N 素促進了低 Na+處理 (T1-N) 根部對 N、P、K 的吸收,其吸收量總體高于其他兩個處理 (T0-N 和 T2-N)。
相關性分析表明,植株 Na 含量 (根與莖葉總吸收量) 與干物重在缺氮脅迫條件下呈顯著負相關關系(r = 0.69*),若每盆花生 Na 吸收量增加 1.0 mg,干物重將下降 1.2 g,同時花生氮、鉀吸收量也顯著下降26.6 mg 和 20.9 mg,而磷素的響應并不明顯 (圖 5)。補施氮素下,花生植株干物重及營養狀況與 Na 吸收卻呈正相關關系,其中 Na 與植株干物重 (r = 0.65*)、磷含量 (r = 0.71*) 和鉀含量 (r = 0.80**) 相關性達顯著水平 (P < 0.05)。
3 討論
含 Na+葉面肥對花生的影響在缺氮脅迫條件下表現為抑制作用,且這種抑制對于高 Na+量處理更為顯著。噴施的 Na+隨氣孔或胞間連絲進入植株體內,易產生滲透脅迫,致使植株體內的一些必需營養元素缺乏[1]。由于 Na+與 K+可競爭結合酶復合物的結合點位,以及 Na+同帶正電荷的 K+、NH4+產生的陽離子易產生一定競爭作用[26],因而本研究中花生鈉含量與氮、鉀含量呈顯著負相關關系,而與磷含量關系不顯著 (H2PO4–、HPO42–帶負電荷)。筆者前期研究證實,噴施液由 pH 5 增加到 pH 7 后,花生磷的吸收濃度增加,而總磷吸收量沒有顯著增加[24]。隨著植株吸收的鈉量增加,花生氮、鉀 (尤其是氮素)吸收受抑制更為顯著,這就使得花生的光合作用受限、凈光合速率顯著下降。事實上,不僅地上部莖葉在噴施含 Na+溶液后干物重顯著下降,地下部根系的發育也受到顯著抑制,根的長度、表面積及體積也顯著下降。莖葉及根系生長受到抑制除了上述原因外,Na 累積能夠導致花生體內蛋白質和核酸的代謝不能正常進行,從而使得一些有毒次生產物 (如游離氨基酸、胺等) 產生,進而影響花生生長發育及干物質累積,且這些抑制作用在缺氮脅迫下表現更為明顯。

圖4 花生莖葉和根中鈉及氮、磷、鉀含量Fig. 4 Contents of Na, N, P and K in shoot and root of peanut

圖5 花生鈉吸收量與干物質累積及氮磷鉀營養的相關關系Fig. 5 Relations between Na accumulation and dry weight of plant and N, P, K contents in peanut
后期補充噴施氮素后,花生受 Na+脅迫現象消除。葉面氮肥可通過增加葉綠素含量,提升光系統PSII 反應中心內部光能轉換效率等,促使葉片光合作用的恢復[23,27]。同時,氮素是蛋白質、氨基酸、核酸等重要組成成分,葉面氮肥也顯著改善了作物營養,從而促進了干物質的累積[28–31]。相關性分析表明,花生對 Na 的吸收在補施氮素下甚至與干物質累積,N、P、K 含量呈正相關關系。對于莖葉的干物重,補施氮素和含 Na+肥料比單獨補施氮素顯著增加,根的干物重也以噴施 Na+溶液和氮素最高。有研究表明,Na+能夠提高植株體中硝酸還原酶的活性及葉綠體對丙酮酸鹽的吸收,推進光合磷酸化過程[6,32–33]。另外,氮肥的施用在促進花生生長發育及干物質累積的同時,對植株體內 Na+的抑制作用也起到一定的稀釋效應。因而,噴施 Na+結合氮肥顯著促進了花生的干物質累積及養分吸收。可見,Na+對花生生長及養分吸收影響與氮素水平有很大關系,后期補施氮肥提高了植株對 Na+的適應性,消除了 Na 脅迫的危害作用,產生“N–Na”協同效應。
4 結論
葉面噴施含 Na+肥料在缺氮脅迫條件下抑制了花生的光合作用、干物質累積及氮、鉀養分吸收,這種抑制隨著噴施 Na+含量增加而更顯著。后期補施氮肥后,這種抑制作用能夠及時消除,花生的光合作用恢復、干物質累積和氮、磷、鉀養分吸收顯著增加。
[1]曲元剛, 趙可夫. NaCl和Na2CO3對鹽地堿蓬脅迫效應的比較[J]. 植物生理與分子生物學報, 2003, 29(5): 387–394. Qu Y G, Zhao K F. Comparision of the stress effects of NaCl and Na2CO3on Suaeda salsa L [J]. Journal of Plant Physiology and Molecular Biology, 2003, 29(5): 387–394.
[2]Surekha K A, Paul V, Zhu J K. Mechanisms of tolerance in plants [M]. Beijing: Tsinghua University Press, 2005: 44–45.
[3]蔡建一, 馬清, 周向睿, 等. Na+在霸王適應滲透脅迫中的生理作用[J]. 草業學報, 2011, 20(1): 89–95. Cai J Y, Ma Q, Zhou X R, et al. Physiological role of Na+in adaption of Zygophyllum xanthoxylum to osmotic stress [J]. Acta Prataculturae Sinica, 2011, 20(1): 89–95.
[4]Sobbarao G V, Ito O, Berry W L, et al. Sodium–a functional plant nutrient [J]. Critical Reviews in Plant Sciences, 2003, 22: 391–416.
[5]鄭小林, 鐘炳輝, 陳榮清. 鉀、鈉離子對富鉀植物離體葉片氣孔運動的影響[J]. 西北植物學報, 2004, 24(2): 320–323. Zheng X L, Zhong B H, Chen R Q. Effect of K+, Na+on isolated leaves’ stomatal movement of rich potassium plants [J]. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(2): 320–323.
[6]李三相, 周向睿, 王鎖民. Na+在植物中的有益作用[J]. 中國沙漠, 2008, 28(3): 485–490. Li S X, Zhou X R, Wang S M. Positive functions of sodium in plants [J]. Journal of Desert Research, 2008, 28(3): 485–490.
[7]雷晶, 郝艷淑, 王典, 等. 鈉鉀替代對不同基因型棉花鉀利用效率的影響[J]. 植物營養與肥料學報, 2015, 21(4): 962–968. Lei J, Hao Y S, Wang D, et al. Substitution effect of sodium and potassium on potassium use efficiency of different cotton genotypes [J]. Journal of Plant Nutrition and Fertilizer, 2015, 21(4): 962–968.
[8]逄煥成, 楊勁松, 謝曉紅. 氯化鈉脅迫下施氮對冬小麥生長發育及體內氯、鈉離子積累的影響[J]. 植物營養與肥料學報, 2005, 11(5): 654–658. Pang H C, Yang J S, Xie X H. Effects of nitrogen fertilization on growth and vivo Cl-and Na+accumulation of winter wheat under NaCl stress [J]. Plant Nutrition and Fertilizer Science, 2005, 11(5): 654–658.
[9]張智猛, 萬書波, 寧堂原, 等. 氮素水平對花生氮素代謝及相關酶活性的影響[J]. 植物生態學報, 2008, 32(6): 1407–1416. Zhang Z M, Wan S B, Ning T Y, et al. Effects of nitrogen on nitrogen metabolism and correlation enzyme activity in peanut [J]. Journal of Plant Ecology, 2008, 32 (6): 1407–1416.
[10]趙平, 孫谷疇, 彭少麟. 植物氮素營養的生理生態學研究[J]. 生態科學, 1998, 17(2): 37–42. Zhao P, Sun G C, Peng S L. Ecophysiological research on nitrogen nutrition of plant [J]. Ecologic Science, 1998, 17(2): 37–42.
[11]曹翠玲, 李生秀, 苗芳. 氮素對植物某些生理生化過程影響的研究進展[J]. 西北農業大學學報, 1999, 27(4): 96–101. Cao C L, Li S X, Miao F. The research situation about effects of nitrogen on certain physiological and biochemical process in plants [J]. Acta Universitatis Agriculturalis Boreali-Occidentalis, 1999, 27(4): 96–101.
[12]林鄭和, 陳榮冰, 陳常頌. 植物對氮脅迫的生理適應機制研究進展[J]. 湖北農業科學, 2011, 50(23): 4761–4764. Lin Z H, Chen R B, Chen C S. Research progress on physiological adaptability of plants to nitrogen deficiency [J]. Hubei Agricultural Sciences, 2011, 50(23): 4761–4764.
[13]李梁, 黃劍華, 陳志偉, 等. 作物耐低氮的相關生物學研究進展[J].上海農業學報, 2012, 28(2): 117–122. Li L, Huang J H, Chen Z W, et al. Progress of biological research on crops tolerant to low nitrogen [J]. Acta Agriculturae Shanghai, 2012, 28(2): 117–122.
[14]萬書波. 中國花生栽培學[M]. 上海: 上海科學技術出版社, 2003: 252–262. Wan S B. China peanut cultivation [M]. Shanghai: Shanghai Scientific and Technical Press, 2003: 252–262.
[15]王才斌, 萬書波. 花生生理生態學[M]. 北京: 中國農業出版社, 2011. Wang C B, Wan S B. Peanut physiological ecology [M]. Beijing: China Agriculture Press, 2011.
[16]胡哲森, 時忠杰, 許長欽. 亞硫酸氫鈉對油茶光合機構的生理效應研究[J]. 林業科學, 2001, 37(專刊1): 68–71. Hu Z S, Shi Z J, Xu C Q. Phosiological effect of sodium bisulfate on photosynthetic apparatus of Camellia oleifera [J]. Scientia Silvae Sinicae, 2001, 37(Special 1): 68–71.
[17]李瑞海. 不同配方葉面肥對作物生長的影響[D]. 南京: 南京農業大學碩士學位論文. 2008. Li R H. Effects of different formulas of foliar fertilizers on cropgrowth [D]. Nanjing: MS Thesis of Nanjing Agricultural University. 2008.
[18]李紅利, 王俊玲, 薛占軍, 等. 硝普鈉對韭菜葉片硝酸鹽累積及營養品質的影響[J]. 河北農業大學學報, 2014, 37(4): 53–57. Li H L, Wang J L, Xue Z J, et al. Effects of sodium nitroprusside (SNP) on nitrate accumulation and nutritional quality in the leaves of Chinese chive [J]. Journal of Agricultural University of Hebei, 2014, 37(4): 53–57.
[19]于會麗, 林治安, 李燕婷, 等. 噴施小分子有機物對小油菜生長發育和養分吸收的影響[J]. 植物營養與肥料學報, 2014, 20(6): 1560–1568. Yu H L, Lin Z A, Li Y T, et al. Effects of spraying low molecular organic compounds on growth and nutrients uptake of rape (Brassica Chinensis L.)[J]. Journal of Plant Nutrition and Fertilizer, 2014, 20(6): 1560–1568.
[20]楊勁松. 作物對不同鹽脅迫和調控條件的響應特征與抗鹽性調控研究[D]. 南京: 南京農業大學博士學位論文. 2006. Yang J S. Characteristics of crop response on different salt stresses/management and the regulation of crop salt resistance [D]. Nanjing: PhD Dissertation of Nanjing Agricultural University. 2006.
[21]慈敦偉, 張智猛, 丁紅, 等. 花生苗期耐鹽性評價及耐鹽指標篩選[J]. 生態學報, 2015, 35(3): 805–814. Ci D W, Zhang Z M, Ding H, et al. Evaluation and selection indices of salinity tolerance in peanut seedling [J]. Acta Ecologica Sinica, 2015, 35(3): 805–814.
[22]萬書波. 花生品種改良與高產優質栽培[M]. 北京: 中國農業出版社, 2008. Wan S B. Peanut variety improvement and high-yield and highquality cultivation [M]. Beijing: China Agriculture Press, 2008.
[23]羅盛, 楊友才, 沈浦, 等. 花生氮素吸收、根系形態及葉片生長對葉面噴施尿素的響應特征[J]. 山東農業科學, 2015, (10): 45–48. Luo S, Yang Y C, Shen P, et al. Characteristics of peanut nitrogen uptake, root morphology and leaf growth to foliar-spraying urea [J]. Shandong Agricultural Sciences, 2015, (10): 45–48.
[24]沈浦, 羅盛, 吳正鋒, 等. 花生磷吸收分配及根系形態對不同酸堿度葉面磷肥的響應特征[J]. 核農學報, 2015, (12): 2418–2424. Shen P, Luo S, Wu Z F, et al. Response of P absorption-allocation rate and root morphology of peanut to P foliar fertilizers with different acidities [J]. Journal of Nuclear Agricultural Sciences, 2015, (12): 2418–2424.
[25]鮑士旦. 土壤農化分析. 第三版[M]. 北京: 中國農業出版社, 2000. Bao S D. Soil and agricultural chemistry analysis (3rd Edition) [M]. Beijing: China Agriculture Press, 2000.
[26]Schachtman D P, Schroeder J I. Structure and transport mechanism of a high-affinity potassium uptake transporter from higher plants [J]. Nature, 1994, 370(6491): 655–658.
[27]許大全. 光合作用效率[M]. 上海: 上海科學技術出版社, 2002. Xu D Q. Photosynthesis efficiency[M]. Shanghai: Shanghai Science and Technology Press, 2002.
[28]李燕婷, 李秀英, 肖艷, 等. 葉面肥的營養機理及應用研究進展[J].中國農業科學, 2009, 42(1): 162–172. Li Y T, Li X Y, Xiao Y, et al. Advances in study on mechanism of foliar nutrition and development of foliar fertilizer application [J]. Scientia Agricultura Sinica, 2009, 42(1): 162–172.
[29]梁雄, 彭克勤, 楊毅. 葉面施肥對花生光合作用和植物激素的影響[J]. 作物研究, 2011, 25(1): 15–18. Liang X, Peng K Q, Yang Y. Effect of foliage spraying on photosynthetic indexes and plant hormones of peanut at blossom stage [J]. Crop Research, 2011, 25(1): 15–18.
[30]吳正鋒. 花生高產高效氮素養分調控研究[D]. 北京: 中國農業大學博士學位論文, 2014. Wu Z F. Nitrogen management for high yield and high efficiency of peanut [D]. Beijing: PhD Dissertation of China Agricultural University, 2014.
[31]李燦東, 郭泰, 王志新, 等. 葉面氮素施量對大豆氮素吸收與分配的影響[J]. 植物營養與肥料學報, 2015, 21 (5): 1361–1365. Li C D, Guo T, Wang Z X, et al. Effects of leaf nitrogen application on soybean nitrogen uptake and distribution [J]. Journal of Plant Nutrition and Fertilizer, 2015, 21 (5): 1361–1365.
[32]Terry N, Ulrich A. Effects of potassium deficiency on the photosynthesis and respiration of leaves of sugar beet [J]. Plant Physiology, 1973, 51: 783–786.
[33]鄒邦基. 鈉的植物營養與生理生態[J]. 植物生理學通訊, 1985, 5: 5–11. Zou B J. Plant nutrition and physiological ecology of sodium [J]. Plant Physiology Communications, 1985, 5: 5–11.
Inhibition of Na+-containing foliar fertilizer on peanut growth under N deficiency stress and recovery effect by supplemental N spraying
SHEN Pu, FENG Hao, LUO Sheng, YU Tian-yi, ZHENG Yong-mei, WU Zheng-feng, CHEN Dian-xu*, SUN Xue-wu*, WANG Cai-bin, ZHENG Ya-ping
( Shandong Peanut Research Institute, Qindao, Shandong 266100, China )
【Objectives】Sodium (Na+) is often contained as counterion in foliar fertilizers, so its effect on crop growth is least considered during fertilization. Na+plays both promotion and inhibition roles on crop growth, depending on the plant nutrition and physiology status. The inhibition role of Na+was studied on peanut exposure to Na+stress, and the recovery effect of supplemental spraying N to the inhibition was also investigated.【Methods】A pot experiment was conducted inside the growth chamber located in Shandong Peanut Research Institute. Three levels of Na+were designed in the foliar fertilizer: Na+0, 2.0 and 10.0 mg/pot, and recorded as T0, T1 and T2 in turn. The peanut photosynthesis, dry matter accumulation, root morphology, nutrient absorption were investigated in both N deficit stresses (T0, T1, T2) and later urea supplement (T0-N, T1-N, T2-N). The effect of Na absorption on the N, P and K absorption was analyzed. 【Results】1) Without N supplement, the net photosynthetic rates were significantly decreased from 11.4 CO2μM/(m2·s) to 6.7–7.5 CO2μM/(m2·s) in Na containing treatments, the dry weights of shoot and root were in order of T0 (6.4 and 2.4 g/pot) > T1 (5.7 and 1.9 g/pot) > T2 (5.3 and 1.5 g/pot). Total root length, surface area, and volume in T1 and T2 treatments were decreased by 7.8%–10.5% and 27.5%–31.8% compared with T0. 2) With later supplement of N, Na+inhibition in photosynthesis was recovered quickly. The shoot dry weights in T1-N and T2-N treatments were increased by 20.3%–25.8% compared with T0-N. The inhibition of Na+became promotion of N, P, K absorption. The shoot N, P, K absorptions in T1-N and T2-N treatments were respectively increased by 9.2%–19.2%, 20.0%–42.3%, 21.4%–24.0% compared with T0-N. The total dry weight of peanut and N, K absorptions were positively correlated with Na+(P < 0.05). 【Conclusions】Under N deficit, Na+shows inhibition effect on the growth and nutrient absorption of peanut and the effect will become exaggerated with the increased Na+contents in fertilizers.This inhibition will be remarkably correlated by later supplement of N, and the inhibition role will change to promotion role of Na+. Therefore, application of Na+containing fertilizer, especially those with high Na+concentration, should consider the N supply in order to relieve the possibly harmful effect of Na+.
sodium stress; growth inhibition; N supplemental supply; recovery effect
S565.2
A
1008–505X(2016)06–1620–08
2016–01–11 接受日期:2016–05–06
山東省農業科學院科技創新重點項目(2014CXZ06-2,2014CXZ11-2);國家自然科學基金項目(41501330,31571617);山東省農業科學院青年科研基金項目(2015YQN05);山東省農業重大應用技術創新課題資助。
沈浦(1986—),男,安徽鳳陽人,博士,助理研究員,主要從事花生栽培與生理研究。E-mail:shenpu109@163.com
* 通信作者 E-mail:chenzhao126@sina.com;E-mail:sunwu50@163.com
