川西高山林線三種灌木凋落葉分解中的無脊椎動物多樣性
2016-11-28 07:18:14和潤蓮陳亞梅鄧長春劉軍偉楊萬勤
生態學報 2016年17期
和潤蓮, 陳亞梅, 鄧長春, 楊 林, 劉軍偉, 楊萬勤, 張 健, 劉 洋
四川農業大學生態林業研究所,高山森林生態系統定位研究站, 長江上游林業生態工程重點實驗室, 長江上游生態安全協同創新中心, 成都 611130
?
川西高山林線三種灌木凋落葉分解中的無脊椎動物多樣性
和潤蓮, 陳亞梅, 鄧長春, 楊 林, 劉軍偉, 楊萬勤, 張 健, 劉 洋*
四川農業大學生態林業研究所,高山森林生態系統定位研究站, 長江上游林業生態工程重點實驗室, 長江上游生態安全協同創新中心, 成都 611130
以無脊椎動物為主體的土壤動物是影響凋落物分解的重要生物因素,對維持陸地生態系統物質循環和能量流動具有重要作用。高山林線交錯帶是高山植被垂直帶譜中重要的過渡區域,擁有比相鄰生態系統更高的生境復雜性和物種多樣性。林線上溫度波動和凍融循環頻率顯著高于針葉林,為了了解林線交錯帶上環境差異對凋落物分解過程中的土壤動物群落結構和多樣性的影響,采用凋落物分解袋的方法,于高山生態系統的兩個主要時期,即雪被末期和生長季末期,研究了林線主要代表性灌木——高山柳(Salixcupularis)、高山杜鵑(Rhododendronlapponicum)和紅毛花楸(Sorbusrufopilosa)凋落葉分解的土壤動物多樣性特征。結果表明:凋落物中的無脊椎動物群落多樣性及個體、類群密度隨物種、海拔梯度和季節而變化,且季節差異對無脊椎動物多樣性的影響比物種和海拔梯度更顯著。3個因子的交互作用不僅影響土壤動物群落多樣性和均勻度,而且影響群落個體密度和類群密度。雪被末期,凋落物中的無脊椎動物多樣性指數H、均勻度指數J及豐富度指數D以針葉林最高,優勢度指數C以林線最高;生長季節末期的無脊椎動物類群密度和個體密度顯著高于雪被末期。總體上,凋落物中的無脊椎動物群落豐富度以生長季末期最高,林線較針葉林豐富。這意味著,未來氣候變暖情景下,灌叢密度增加,凋落物輸入量增大,可能導致無脊椎動物多樣性增加。
無脊椎動物;高山林線;灌木;凋落葉;雪被
以無脊椎動物為主體的土壤動物群落是影響凋落物分解的重要生物因素,對維持陸地生態系統物質循環和能量流動具有重要作用[1]。伴隨氣候變化對高山森林生態系統的影響,土壤動物對高山/亞高山森林凋落物分解的作用逐漸受到廣泛關注[2- 10],研究主要集中于高山森林中岷江冷杉(Abiesfaxoniana)、紅樺(Betulaalbo-sinensis)、方枝柏(Sabinasaltuaria)等優勢樹種凋落葉的分解及其與土壤動物群落結構、多樣性的關系,而對于交錯帶區域的土壤動物多樣性缺乏必要的關注。高山林線交錯帶(Alpine timberline)是高山植被垂直帶譜中重要的過渡區域[11],擁有比相鄰生態系統更高的生境復雜性和物種多樣性,對環境變化較為敏感,是研究高山生態系統物種多樣性及其對氣候變化的理想研究區域[12- 13]。林線上溫度驅動的凍融循環顯著高于針葉林[14],嚴酷的環境條件極大地限制了雪被下土壤動物的活動,這對于凋落物分解過程中的無脊椎動物群落結構和多樣性將會產生什么影響?關于高山林線交錯帶無脊椎動物與凋落物分解的相互作用及其對氣候變化的響應還有許多不確定因素,亟待深入研究。
氣候變暖可能導致氣候帶的移動、植被格局等變化[15],如林線位置的上移及樹木密度的增加等[16]。植被的遷移通常會導致植物多樣性的改變,而無脊椎動物主要以分解碎屑及腐敗植物殘體為食,因此植物種類的變化將直接影響食物網中無脊椎動物群落的分布[17]。團隊前期研究結果表明,川西高山生態系統不同植被類型下無脊椎動物群落結構存在顯著差異,灌木林無脊椎動物群落多樣性較針葉林和高山草甸高[5]。高山林線交錯帶的植被類型從暗針葉林逐漸過渡到灌叢草甸,灌木替代岷江冷杉成為林線的優勢物種,但林線上灌木凋落物分解中的無脊椎動物群落組成和多樣性仍缺乏必要的關注,這極大的限制了對交錯帶區域上無脊椎動物群落結構特征的認識。因此,在前期研究[5,13- 14]的基礎上,采用凋落物分解袋法,以高山林線交錯帶灌木代表性植物高山柳(Salixcupularis)、高山杜鵑(Rhododendronlapponicum)和紅毛花楸(Sorbusrufopilosa)凋落葉為研究對象,對比研究高山針葉林和林線3種灌木凋落葉分解過程中的無脊椎動物群落結構和多樣性,以期為深入了解高山林線交錯帶無脊椎動物群落結構,以及生長季節和雪被期無脊椎動物群落之間的相互聯系提供一定的基礎數據。
1 試驗地概括
研究區區域位于四川省阿壩藏族、羌族自治州理縣米亞羅鷓鴣山(31°50′—31°53′E, 102°40′—102°44′N) 處于青藏高原東緣褶皺帶最外緣部分,岷江上游支流雜谷腦河上游地區。鷓鴣山山體海拔3 200—4 800 m,山地垂直地帶性明顯,自河谷至山頂依次分布的植被類型有針闊混交林、暗針葉林、高山疏林灌叢、高山草甸,4 500m以上為高山荒漠和積雪帶。氣候屬于冬寒夏涼的高寒氣候,年平均氣溫6—12 ℃,1月平均氣溫-8 ℃,7月平均氣溫12.6 ℃,年積溫1 200—1 400 ℃,年降水量600—1 100 mm,年蒸發量1 000—1 900 mm。2012年10月底到2013年11月初一年的日均溫變化情況如圖1。由圖1知,一年內林線上溫度變化波動很大,而針葉林上溫度變化平緩。從2012年10月底到2013年4月底,日均溫在0℃以下,從5月份開始逐漸回溫,6月份達到峰值,7、8月份的日均溫保持較高。地形以石質山地為主,土壤具有粗骨、石礫含量高和薄層等特征。從低海拔到高海拔,土壤類型依次為山地暗棕壤、棕色針葉林土、高山草甸土。主要灌木樹種有大理杜鵑(Rhododendrontaliense)、皺皮杜鵑(R.wiltonii)、高山杜鵑(R.lapponicum)、紅花花楸(Sorbusrufopilosa)、柳葉忍冬(Loniceralanceolata)、越桔葉忍冬(L.myrtillus)、窄葉鮮卑花(Sibiraeaangustata)和高山柳(Salixcupularis)等。每年從4月到10月為生長季末期,冬季雪被末期明顯,從10月到次年4月,長達6—7個月。
2 研究方法
2.1 實驗設計
2012年10月在鷓鴣山針葉林和林線上采集優勢灌木物種的凋落葉,包括高山柳、紅毛花楸和高山杜鵑3個物種,帶回室內將其風干,每個物種稱取10 g裝入20 cm × 20 cm(孔徑為1 mm)的分解袋中,每個物種稱取12袋,共36個凋落袋。10月底分別將凋落物袋平鋪埋設在海拔3900m的針葉林和海拔4000m的林線地表,每個海拔每個物種放置6袋,并用釘子固定位置。同時每個海拔隨機選取一個凋落袋,袋內埋設一個紐扣式溫度記錄儀(iButton DS1923-F5, Maxim/Dallas Semiconductor, Sunnyvale, CA, USA),設定為每3h記錄1次溫度數據,計算凍融頻次。同時,在放置凋落袋的附近用15竹竿插入土壤中,用作雪被厚度測定的標記,在雪被末期每月觀測記錄雪被的厚度情況。
2.2 樣品處理
2013年5月1日和2013年11月7日分別代表雪被末期末和生長季末期,分別采集每個處理的凋落葉分解袋各3袋,將所采的分解袋裝入密封而透氣的黑布袋里面低溫保存,迅速帶回室內,先將袋外的泥土殘落物輕輕去除,然后將袋內的殘落物放置于改良的Tullgren干漏斗中分離提取中小型無脊椎動物,采用體式解剖鏡和生物顯微鏡鏡檢計數和分類,并參照《中國土壤動物檢索圖鑒》[18]鑒定到科。分離結束后,清除殘落物表面的土壤顆粒和混入的雜物,60℃下烘干至恒重,稱量殘落物干重。
2.3 數據處理與分析
采用Shannon指數H′、Simpson優勢度指數C、Pielou均勻度指數J和密度-類群指數DG計算各海拔各無脊椎動物群落多樣性。各指數計算公式如下[8]:
(1)Shannon多樣性指數
(2)Simpson優勢度指數
C=∑(Pi)2
(3)Pielou均勻度指數
J=H′/lnS
(4)Margalef 豐富度指數
D= (S- 1)/lnN
式中,ni為第i個物種的個體數,S為類群數,N為無脊椎動物總個體數,Pi為第i類群的百分比。
各類群優勢度的劃分:個體數量占捕獲總數10.0% 以上的為優勢類群,1.0%—10.0% 的為常見類群,1.0%以下的為稀有類群[1,5]。
數據分析在SPSS17.0中進行,采樣方差分析和獨立樣本T檢驗比較環境因子和無脊椎動物群落結構間的差異,Pearson相關性分析環境因子與無脊椎動物群落結構特征的相關性,顯著性水平設定為a=0.05。相關圖表制作在Excel中完成。
3 結果與分析
3.1 凋落葉分解的環境因子
2012 年10月30日—2013年11月7日針葉林和林線上的日平均溫度變化如圖1所示。針葉林和林線上溫度變化趨勢一致,但針葉林日均溫變化平穩,而林線日均溫波動幅度較大。從10月底到次年1月,氣溫逐漸下降,12月底1月初溫度最低,從1月中下旬開始,氣溫逐漸回升,到5月份日均溫達到0℃以上,在7月份達到峰值,到次年10月份溫度又開始降到0℃以下。
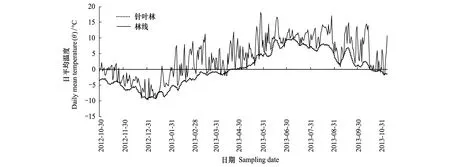
圖1 針葉林和林線凋落袋內年日均溫度動態Fig.1 Dynamics of daily mean temperature in litter bag in the coniferous forest and timberline within a year
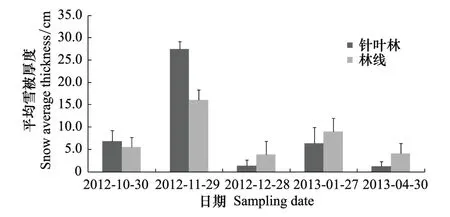
圖2 雪被期針葉林和林線的雪被厚度 Fig.2 Snow depth of the alpine forest and timberline during snow-covered season
雪被期針葉林和林線上雪被厚度如圖2所示。針葉林和林線上的雪被均在11月份最厚,3月初針葉林和林線上沒有監測到雪被厚度。T檢驗發現,雪被期針葉林和林線上的雪被厚度均差異不顯著。兩個時期針葉林和林線上的平均溫度、雪被厚度、凍融頻次、正積溫和負積溫如表1所示。雪被期日均溫在0℃以下,生長季節日均溫在0℃以上,針葉林內平均雪被厚度高于林線上,且兩個時期林線上的日均溫、凍融頻次、正積溫和負積溫均高于針葉林上,進一步說明林線上溫度變化劇烈。
3.2 無脊椎動物群落結構
雪被末期和生長季末期2次采樣凋落物中中小型無脊椎動物共714只,隸屬1門3綱12目32類(科)(表2),不同季節林線交錯帶3種灌木凋落葉中獲得的無脊椎動物群落結構有所變化。雪被末期3種凋落葉中獲得無脊椎動物18科,共111只,其中針葉林中獲得15科,28只,林線上獲得10科,83只。生長季末期3種凋落葉中獲得無脊椎動物25科,共603只,其中針葉林中獲得無脊椎動物14科,335只,林線上獲得無脊椎動物21科,268只。若甲螨科和隱翅甲科僅出現在雪被末期針葉林上的高山杜鵑凋落葉中,大蚊科和管薊馬亞科僅出現在雪被末期針葉林上紅毛花楸的凋落葉中,羅甲螨科僅出現在雪被末期林線上高山杜鵑凋落葉中,而寄螨科和氣肢蟲科僅出現在生長季末期林線上的紅毛花楸凋落葉中,纓甲科和卷甲蟲科僅出現在生長季末期針葉林上高山杜鵑凋落葉中,同翅目僅出現在生長季末期林線上高山杜鵑凋落葉中。兩個季節針葉林和林線上無脊椎動物優勢類群如表3。雪被末期,針葉林中彈尾目、中氣門亞目、甲螨亞目和雙翅目幼蟲為優勢類群,前氣門亞目、鞘翅目和纓翅目為常見類群;林線上以前氣門亞目和中氣門亞目為優勢類群,以彈尾目、甲螨亞目和雙翅目幼蟲為常見類群。生長季末期,針葉林中以彈尾目和中氣門亞目為優勢類群,以中氣門亞目、甲螨亞目和鱗翅目幼蟲為常見類群,以雙翅目幼蟲、鞘翅目、等足目和嚙目為稀有類群;林線上以彈尾目、前氣門亞目和中氣門亞目為優勢類群,以甲螨亞目、鉗亞目和同翅目為常見類群,以雙翅目幼蟲、鱗翅目幼蟲、等足目和嚙目為稀有類群。纓翅目僅出現在雪被末期,鱗翅目幼蟲、雙尾目、等足目、嚙目和同翅目僅在生長季末期出現。無脊椎動物群落結構同時受到凋落葉物種和季節的影響。
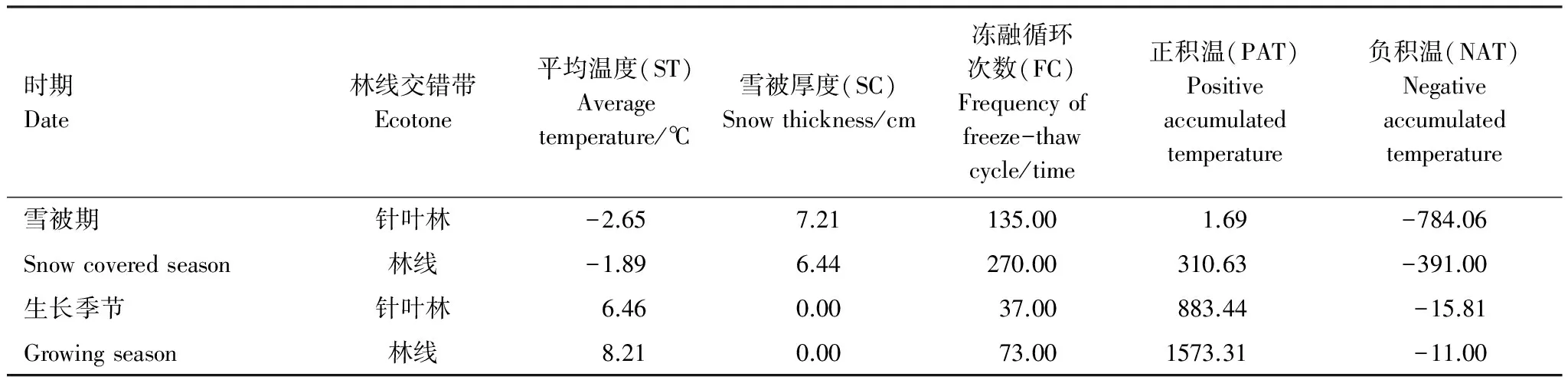
表1 雪被末期和生長季末期環境因子
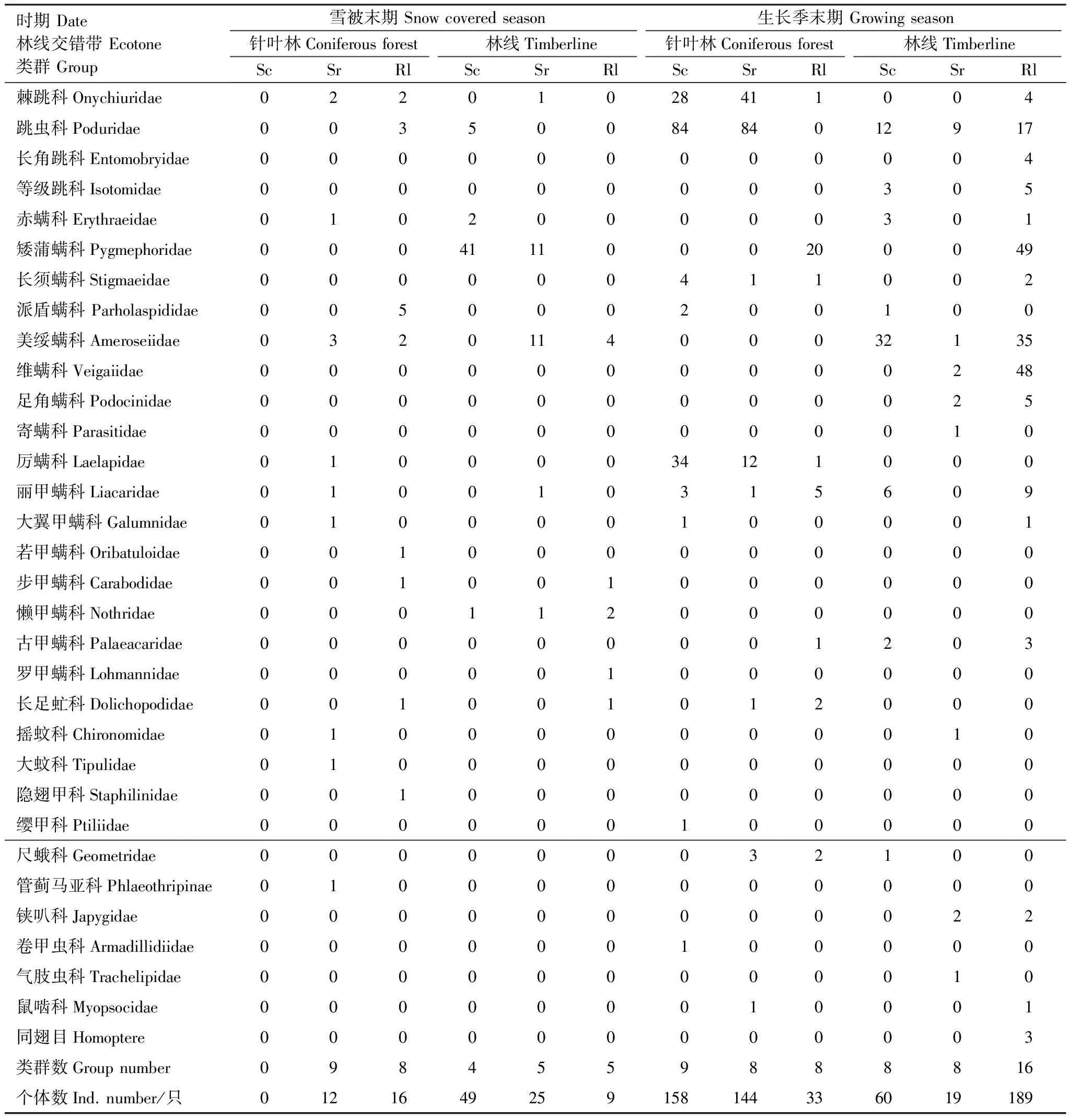
表2 雪被末期和生長季節末期凋落物無脊椎動物群落的種類和數量組成
Sc:高山柳Salixcupularis; Sr:紅毛花楸Sorbusrufopilosa; Rl:高山杜鵑Rhododendronlapponicum
兩個時期針葉林和林線上灌木凋落葉中無脊椎動物個體密度和類群密度如圖3所示。個體密度和類群密度在生長季末期均高于雪被末期,并且個體密度在針葉林上高山柳凋落葉中(F=7.682,P<0.05)和林線上高山杜鵑凋落葉中差異顯著(F=13.609,P<0.05),類群密度在林線上高山杜鵑凋落葉中差異顯著(F=12.914,P<0.05)。說明生長季更適合不同類群無脊椎動物的生存,獲得的個體數量也比雪被期高。
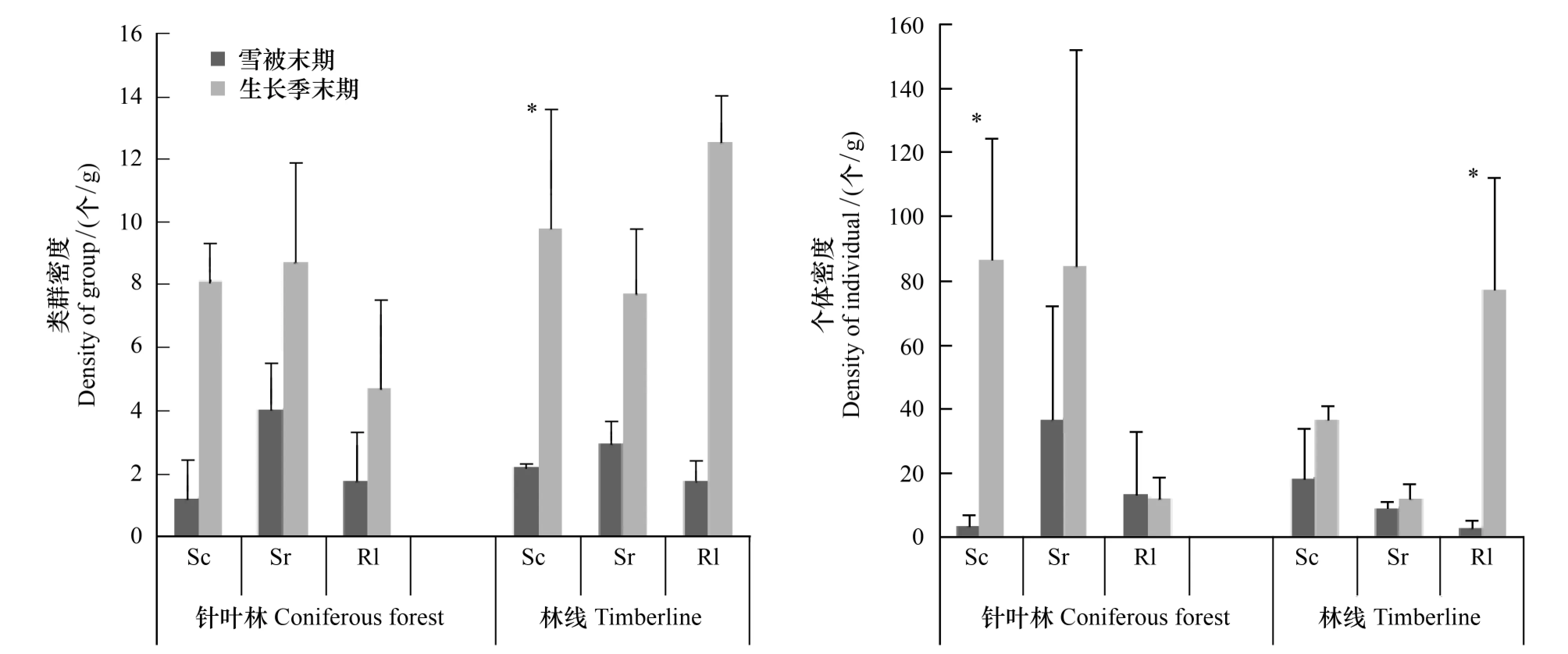
圖3 無脊椎動物個體密度和類群密度(平均值±標準誤,*表示季節間差異顯著) Fig.3 Individual density and group density of invertebrate (Mean ± SE,*indicate significant difference between two seasons, * P < 0.05)
3.3 無脊椎動物群落多樣性特征
采用T檢驗,比較雪被末期和生長季末期林線和針葉林的各物種凋落葉的無脊椎動物多樣性(圖4)。在雪被末期,高山柳凋落葉內沒有無脊椎動物外,Shannon指數H、Pielou指數J和Margalef指數D在針葉林中高于林線上,Simpson指數C相反,并且高山柳凋落葉中Shannon指數H(F=10.048,P<0.05)、Pielou指數J(F=10.047,P<0.05)和Simpson指數C(F=10.305,P<0.05)在針葉林和林線上差異顯著,紅毛花楸凋落葉中Pielou指數J(F=9.844,P<0.05)在針葉林和林線上差異顯著;在生長季末期,高山柳凋落葉中的Pielou指數J、紅毛花楸和高山杜鵑凋落葉中的Simpson指數C針葉林上高于林線上,各凋落葉中的Shannon指數H、Margalef指數D、高山柳和紅毛花楸凋落葉中的Pielou指數J和高山柳凋落葉中的Simpson指數C在林線上高于針葉林上,并且差異不顯著(P>0.05)。總體來說,生長季末期無脊椎動物較雪被末期豐富,林線上較針葉林豐富。
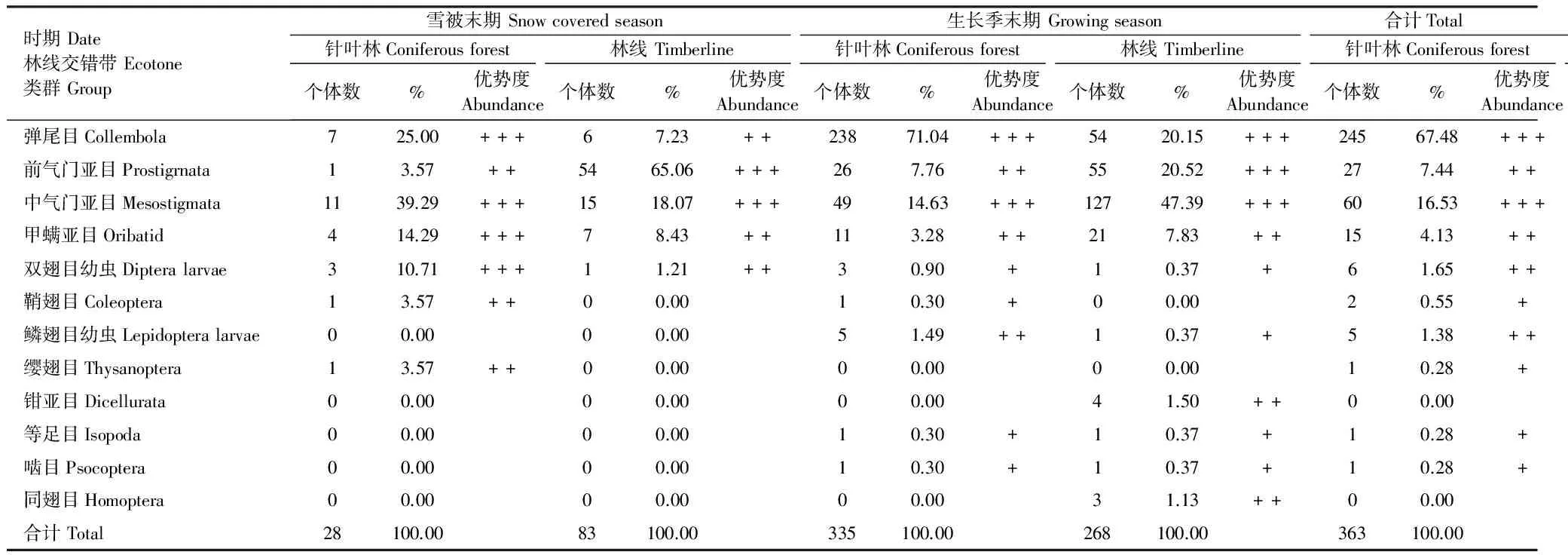
表3 雪被末期和生長季節末期凋落物無脊椎動物群落的種類和數量組成
優勢類群:密度占總密度的10%以上 Average density accounted for more than 10% of the total; 常見類群:密度占總密度的1%—10% Average density accounted for 1%—10% of the total; 稀有類群:密度占總密度的1%以下 Average density accounted for less than 1% of the total
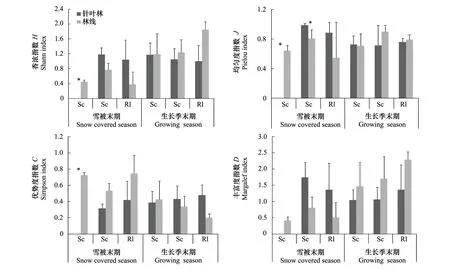
圖4 雪被末期和生長季節末期凋落物無脊椎動物群落多樣性季節動態(平均值±標準誤,*表示季節間差異顯著)Fig.4 Seasonal dynamics of invertebrate community diversity of snow covered season and growing season (Mean ± SE,*indicate significant difference between two seasons, * P < 0.05, ** P < 0.01)
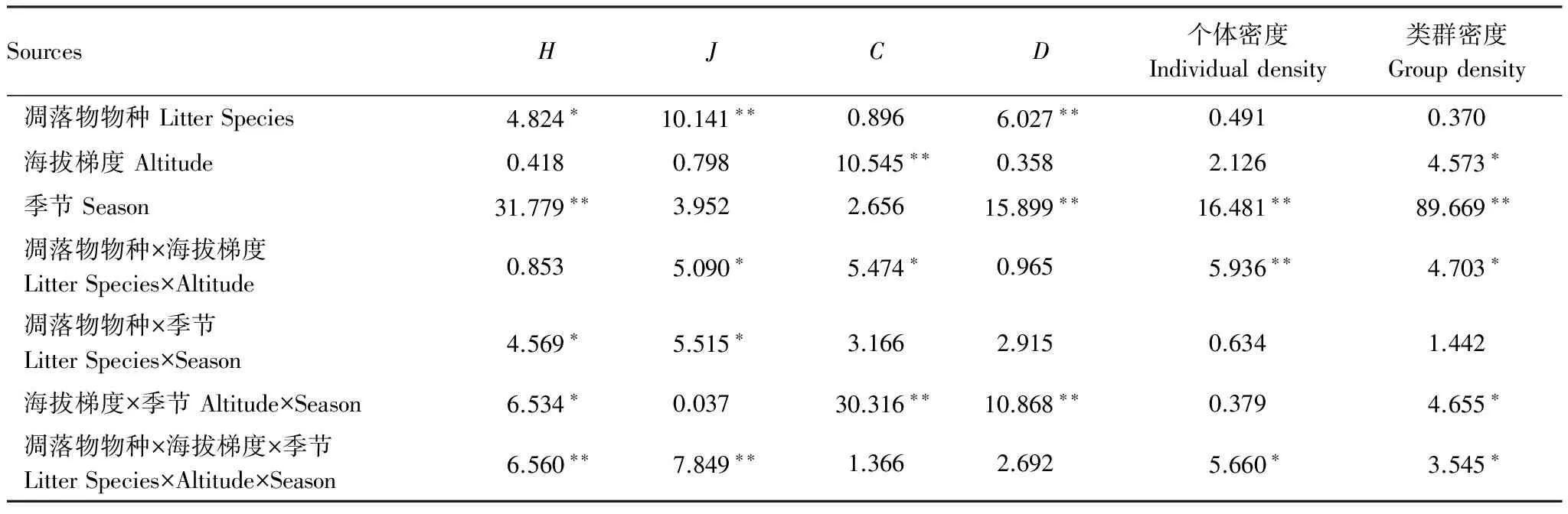
SourcesHJCD個體密度Individualdensity類群密度Groupdensity凋落物物種LitterSpecies4.824?10.141??0.8966.027??0.4910.370海拔梯度Altitude0.4180.79810.545??0.3582.1264.573?季節Season31.779??3.9522.65615.899??16.481??89.669??凋落物物種×海拔梯度LitterSpecies×Altitude0.8535.090?5.474?0.9655.936??4.703?凋落物物種×季節LitterSpecies×Season4.569?5.515?3.1662.9150.6341.442海拔梯度×季節Altitude×Season6.534?0.03730.316??10.868??0.3794.655?凋落物物種×海拔梯度×季節LitterSpecies×Altitude×Season6.560??7.849??1.3662.6925.660?3.545?
H:香濃多樣性指數 Shannon index;J:均勻度指數 Pielou index;C:優勢度指數 Simpson index;D:豐富度指數Margalef index; *P< 0.05, **P< 0.01
凋落物種類、海拔梯度和季節對無脊椎動物群落結構均有一定的影響(表4)。凋落物物種對多樣性指數H(P<0.05)、J和D(P<0.01)有顯著的影響;海拔梯度對C指數(P<0.01)和無脊椎動物類群密度(P<0.05)有顯著影響;季節差異對無脊椎動物多樣性指數H、D、個體密度和類群密度均有極顯著的影響(P<0.01)。凋落物種類和海拔梯度的交互作用對多樣性指數J、C、類群密度(P<0.05)和個體密度(P<0.01)均有顯著影響;凋落物種類和季節的交互作用對多樣性指數H和J影響顯著;海拔梯度和季節的交互作用對多樣性指數H、C、D以及對類群密度均有顯著影響;凋落物種類、海拔梯度和季節差異3個因子的交互對無脊椎動物個體密度和類群密度、多樣性指數H、J有顯著影響。
3.4 無脊椎動物與環境因子的關系
川西高山針葉林和林線上灌木凋落葉中無脊椎動物群落結構與環境因子的相關分析結果如表5。無脊椎動物群落的多樣性指數H、D、個體密度和類群密度均與雪被厚度和凍融頻次極顯著負相關,與正積溫顯著正相關,多樣性指數J與日均溫和正、負積溫正相關,與雪被厚度和凍融頻次負相關,多樣性指數C與凍融頻次極顯著正相關。相關分析結果說明,溫度、雪被和凍融循環頻率對無脊椎動物群落結構和多樣性的影響顯著。
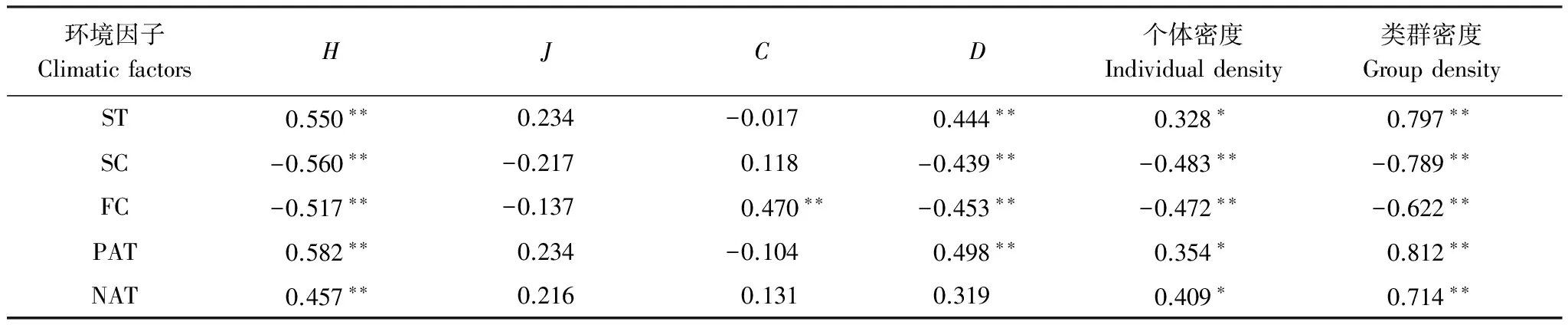
表5 雪被末期和生長季節末期凋落物無脊椎動物多樣性與環境因子的相關性分析
**在0.01水平上顯著相關;*在0.05水平上顯著相關
4 討論
在全球尺度上,氣候因素對凋落物的分解起主導作用,溫度和濕度被認為是影響凋落物分解主要的氣候因子[19],其次是凋落物質量和土壤生物作用。最近的一些分解模型研究發現,氣候和凋落物質量可以解釋全球凋落物分解速率的60%—70%,另外的30%—40%主要是生物因素的作用[20]。從全球無脊椎動物對凋落物分解的貢獻來看,如果排除無脊椎動物的影響,凋落物分解速率將顯著降低35%[21]。凋落物為土壤生物提供食物來源和能量,而土壤生物的活動對凋落物的分解起促進作用[19]。無脊椎動物群落結構和多樣性受到氣候、植被類型、土壤、凋落物種類及小氣候的影響[2,8,14,22]。研究發現,川西高山森林和林線由于郁閉度和植被類型變化,溫度、雪被厚度和凍融頻次等環境因子差異明顯。因此,導致海拔梯度上無脊椎動物群落結構和多樣性的差異,尤其是季節變化對無脊椎動物群落結構和多樣性的影響最為顯著,生長季末期無脊椎動物個體密度和類群密度、多樣性指數H、D均高于雪被末期,這與團隊前期研究結果類似[7- 9,23],與David和Gillon[24]與wang[2]等認為溫濕度和無脊椎動物多樣性密切相關的研究結果一致,進一步證實了季節變化造成的溫度、雪被等環境因子影響著無脊椎動物群落組成及多樣性[14,23,19]。氣候是影響無脊椎動物群落結構和多樣性的決定性因素[21,25],生長季節更高的溫度更適合無脊椎動物的生存繁衍[8]。川西高山地區冬季環境嚴酷,低溫不僅嚴重制約了無脊椎動物活動,導致無脊椎動物群落表現出較低的活性甚至休眠狀態[26-28],同時還凍結了凋落物的養分資源,使得可利用的食物來源嚴重匱乏[23],無脊椎動物難以取食。然而,雪被末期無脊椎動物數量仍占到總數的15.5%,說明在冬季惡劣環境條件下,仍有部分類群和數量的無脊椎動物生存活動[8]。伴隨著生長季的回暖,部分休眠或蟄伏無脊椎動物開始復蘇[29],個體數量和類群數顯著上升。同時,生長季節水熱同季,植被生長旺盛,微生物活性較高,林線交錯帶土壤微生物數量和生物量表現為生長季末期高于雪被末期[30],這為無脊椎動物的生存和繁衍提供了良好的食物來源[31-32]。
海拔差異導致無脊椎動物所處的微環境發生變化,對高山森林和林線上無脊椎動物多樣性及類群密度有重要影響。本研究發現,海拔梯度對凋落物中無脊椎動物優勢度指數C和類群密度影響顯著。雪被期林線上獲得的個體數和類群數較針葉林豐富,生長季節針葉林內獲得的個體數較多,但類群數在林線上更豐富(表2),進一步佐證了林線交錯帶具有較高的物種豐富度[5,12]。兩個海拔獲得的部分優勢類群和常見類群相同,但稀有類群差異較大,如鞘翅目、纓翅目僅在針葉林出現,同翅目僅出現在林線上,可能是因為高山林線處于森林向灌叢轉變的過渡帶上,從暗針葉林過渡到灌叢草甸,植被類型與土壤環境因子發生了顯著的變化,導致林線交錯區的光強、溫濕度、雪被、凍融循環等微環境產生很大的差異,這些變化對無脊椎動物的食物源和棲息環境產生很大的影響,它們直接或間接作用于無脊椎動物群落,從而導致無脊椎動物群落結構和多樣性發生了顯著變化[2,5,14]。在高緯度和高海拔地區,雪被能對這些控制凋落物分解的氣候因子產生重要影響[33]。Templer等[34]研究發現,美國北方闊葉林冬季雪被厚度減少和持續時間縮短,降低了森林地面的節肢動物的豐度和多樣性,改變了群落結構和組成。在未來氣候變暖條件下,隨著氣溫升高,冬季雪線位置的上移[35],高山生態系統中無脊椎動物群落結構勢必發生改變,可能使高寒物種、地方特有種或其中的優勢種因為生境的改變而瀕臨滅絕或被其他物種替代[36]。
凋落物的種類、基質質量影響無脊椎動物的數量分布和類群組成[37]。本研究發現,物種對凋落葉中無脊椎動物多樣性指數H、J和D影響顯著,3種灌木凋落葉中無脊椎動物捕獲量以高山杜鵑最多,紅毛花楸次之,高山柳最少。長角跳科、若甲螨科、步甲螨科、羅甲螨科和隱翅甲科僅出現在高山杜鵑凋落葉中,寄螨科、搖蚊科、大蚊科、管薊馬亞科和氣肢蟲科僅出現在紅毛花楸凋落葉中,纓甲科和卷甲蟲科僅出現在高山柳凋落葉中。值得注意的是,雪被期針葉林內高山柳凋落葉中沒有分離到無脊椎動物,可能是無脊椎動物遷移和活動的偶然性造成的。方差分析結果表明,季節、海拔梯度、凋落物種類兩兩交互或3個因子的交互對無脊椎動物個體密度和類群密度、多樣性指數有顯著影響,且季節和海拔梯度的交互作用更加顯著,說明季節對各海拔的環境因子影響程度不同,對不同凋落物種類的影響也有差異,3個因子綜合作用對土壤動物群落結構造成影響。由于高山植物對氣候變暖的響應最為敏感[38],氣候變暖可能導致林線上移以及灌叢分布密度增加[13,38],從而影響凋落物種類和基質質量[15-16],勢必改變無脊椎動物群落的分布格局和多樣性[25],如灌木凋落葉中的特有種羅甲螨科、纓甲科等可能隨灌木密度的增加和凋落物輸入量增大而增加。同時,未來氣候變化將改變無脊椎動物對凋落物分解的作用方式,尤其是高寒地區的氣候變化對凋落物分解的影響將更為顯著[19],隨著降水的增多和最低溫的升高,無脊椎動物對凋落物分解的作用增大[21],表現在直接加速凋落物分解和有機碳庫的釋放,間接地提高無脊椎動物對凋落物分解的貢獻率。
[1] Swift M J, Heal O W, Anderson J M. Decomposition in Terrestrial Ecosystems. Oxford: Blackwell Scientific Publications, 1979.
[2] Wang S J, Ruan H H, Wang B. Effects of soil microarthropods on plant litter decomposition across an elevation gradient in the Wuyi Mountains. Soil Biology and Biochemistry, 2009, 41(5): 891- 897.
[3] Donoso D A, Johnston M K, Kaspari M. Trees as templates for tropical litter arthropod diversity. Oecologia, 2010, 164(1): 201- 211.
[4] Gavazov K S. Dynamics of alpine plant litter decomposition in a changing climate. Plant and Soil, 2010, 337(1/2): 19- 32.
[5] 黃旭, 文維全, 張健, 楊萬勤, 劉洋, 閆幫國, 黃玉梅. 川西高山典型自然植被土壤動物多樣性. 應用生態學報, 2010, 21(1): 181- 190.
[6] 夏磊, 吳福忠, 楊萬勤. 季節性凍融期間土壤動物對岷江冷杉凋落葉質量損失的貢獻. 植物生態學報, 2011, 35(11): 1127- 1135.
[7] 夏磊, 吳福忠, 楊萬勤, 譚波. 川西亞高山森林凋落物分解初期土壤動物對紅樺凋落葉質量損失的貢獻. 應用生態學報, 2012, 23(2): 301- 306.
[8] 譚波, 吳福忠, 楊萬勤, 夏磊, 楊玉蓮, 王奧. 川西亞高山/高山森林大型土壤動物群落多樣性及其對季節性凍融的響應. 生物多樣性, 2012, 20(2): 215- 223.
[9] 劉瑞龍, 李維民, 楊萬勤, 譚波, 王文君, 徐振鋒, 吳福忠. 土壤動物對川西高山/亞高山森林凋落物分解的貢獻. 應用生態學報, 2013, 24(12): 3354- 3360.
[10] Wu P F, Liu X L, Liu S R, Wang J X, Wang Y. Composition and spatio-temporal variation of soil microarthropods in the biodiversity hotspot of northern Hengduan Mountains, China. European Journal of Soil Biology, 2014, 62: 30- 38.
[11] Holtmeier F K. Ecological aspects of climatically caused timberline fluctuations // Benist M, ed. Mountain Environments in Changing Climates. Routledge. New York: London, 1994, 220- 233.
[12] 石培禮, 李文華, 王金錫, 劉興良. 四川臥龍亞高山林線生態交錯帶群落的種多度關系. 生態學報, 2000, 20(3): 384- 389.
[13] 劉洋, 張健, 楊萬勤. 高山生物多樣性對氣候變化響應的研究進展. 生物多樣性, 2009, 17(1): 88- 96.
[14] 和潤蓮, 陳亞梅, 鄧長春, 楊萬勤, 張健, 劉洋. 雪被期川西高山林線交錯帶兩種地被物凋落物分解與土壤動物多樣性. 應用生態學報, 2015, 26(3): 723- 731.
[15] Walther G R, Bei?ner S, Burga C A. Trends in the upward shift of alpine plants. Journal of Vegetation Science, 2005, 16(5): 541- 548.
[16] Klasner F L, Fagre D B. A half century of change in alpine treeline patterns at Glacier National Park, Montana, U.S.A. Arctic, Antarctic, and Alpine Research, 2002, 34(1): 49- 56.
[17] Wardle D A, Yeates G W, Barker G M, Bonner K I. The influence of plant litter diversity on decomposer abundance and diversity. Soil Biology and Biochemistry, 2006, 38(5): 1052- 1062.
[18] 尹文英, 胡圣豪, 沈韞芬, 寧應之, 孫希達, 吳紀華, 諸葛燕, 張云美, 王敏, 陳建英, 徐成綱, 梁彥齡, 王洪鑄, 楊潼, 陳德牛, 張國慶, 宋大祥, 陳軍, 梁來榮, 胡成業, 王慧芙, 張崇洲, 匡溥人, 陳國孝, 趙立軍, 謝榮棟, 張駿, 劉憲偉, 韓美貞, 畢道英, 肖寧年, 楊大榮. 中國土壤動物檢索圖鑒. 北京: 科學出版社, 1998.
[19] Aerts R. The freezer defrosting: global warming and litter decomposition rates in cold biomes. Journal of Ecology, 2006, 94(4): 713- 724.
[20] Parton W, Silver W L, Burke I C, Grassens L, Harmon M E, Currie W S, King J Y, Adair E C, Brandt L A, Hart S C, Fasth B. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science, 2007, 315(5810): 361- 364.
[21] García-Palacios P, Maestre F T, Kattge J, Wall D H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecology Letters, 2013, 16(8): 1045- 1053.
[22] Maharning A R, Mills A A A, Adl S M. Soil community changes during secondary succession to naturalized grasslands. Applied Soil Ecology, 2009, 41(2): 137- 147.
[23] Tan B, Wu F Z, Yang W Q, Liu L, Yu S. Characteristics of soil animal community in the subalpine/alpine forests of western Sichuan during onset of freezing. Acta Ecologica Sinica, 2010, 30(2): 93- 99.
[24] David J F, Gillon D. Combined effects of elevated temperatures and reduced leaf litter quality on the life-history parameters of a saprophagous macroarthropod. Global Change Biology, 2009, 15(1): 156- 165.
[25] Wall D H, Bradford M A, John M G S T, Trofymow J A, Behan-Pelletier V, Bignell D E, Dangerfield J M, Parton W J, Rusek J, Voigt W, Wolters V, Gardel H Z, Ayuke F O, Bashford R, Beljakova O I, Bohlen P J, Brauman A, Flemming S, Henschel J R, Johnson D L, Jones T H, Kovarova M, Kranabetter J M, Kutny L, Lin K C, Maryati M, Masse D, Pokarzhevskii A, Rahman H, Sabará M G, Salamon J A, Swift M J, Varela A, Vasconcelos H L, White D, Zou X M. Global decomposition experiment shows soil animal impacts on decomposition are climate-dependent. Global Change Biology, 2008, 14(11): 2661- 2677.
[26] Wu F Z, Yang W Q, Zhang J, Deng R J. Litter decomposition in two subalpine forests during the freeze-thaw season. Acta Oecologica, 2010, 36(1): 135- 140.
[27] 王振中, 張友梅, 邢協加. 土壤環境變化對土壤動物群落影響的研究. 土壤學報, 2002, 39(6): 892- 897.
[28] Briones M J I, Ostle N J, McNamara N P, Poskitt J. Functional shifts of grassland soil communities in response to soil warming. Soil Biology and Biochemistry, 2009, 41(2): 315- 322.
[29] Campbell J L, Mitchell M J, Groffman P M, Christenson L M, Hardy J P. Winter in northeastern North America: a critical period for ecological processes. Frontiers in Ecology and the Environment, 2005, 3(6): 314- 322.
[30] 劉洋, 張健, 閆幫國, 黃旭, 徐振鋒, 吳福忠. 青藏高原東緣高山森林-苔原交錯帶土壤微生物生物量碳、氮和可培養微生物數量的季節動態. 植物生態學報, 2012, 36(5): 382- 392.
[31] 苗雅杰, 殷秀琴. 小興安嶺紅松闊葉混交林土壤動物群落研究. 林業科學, 2005, 41(2): 204- 209.
[32] Gongalsky K B, Persson T, Pokarzhevskii A D. Effects of soil temperature and moisture on the feeding activity of soil animals as determined by the bait-lamina test. Applied Soil Ecology, 2008, 39(1): 84- 90.
[33] Baptist F, Yoccoz N G, Choler P. Direct and indirect control by snow cover over decomposition in alpine tundra along a snowmelt gradient. Plant and Soil, 2010, 328(1/2): 397- 410.
[34] Templer P H, Schiller A F, Fuller N W, Socci A M, Campbell J L, Drake J E, Kunz T H. Impact of a reduced winter snowpack on litter arthropod abundance and diversity in a northern hardwood forest ecosystem. Biology and Fertility of Soils, 2012, 48(4): 413- 424.
[35] IPCC. Climate Change 2007: the physical science basis // Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 2007.
[36] Klanderud K, Totland ?. Simulated climate change altered dominance hierarchies and diversity of an alpine biodiversity hotspot. Ecology, 2005, 86(8): 2047- 2054.
[37] Siemann E, Tilman D, Haarstad J, Ritchie M. Experimental tests of the dependence of arthropod diversity on plant diversity. The American Naturalist, 1998, 152(5): 738- 750.
[38] 陳建國, 楊揚, 孫航. 高山植物對全球氣候變暖的響應研究進展. 應用與環境生物學報, 2011, 17(3): 435- 446.
Invertebrate diversity in foliar litter of three shrubs in the alpine timberline of western Sichuan
HE Runlian, CHEN Yamei, DENG Changchun, YANG Lin, LIU Junwei, YANG Wanqin, ZHANG Jian, LIU Yang*
Collaborative innovation center of ecological security in the upper reaches of Yangtze river, Long-term Research Station of Alpine Forest Ecosystems, Key Laboratory of Ecological Forestry Engineering in Sichuan Province, Institute of Ecology & Forestry, Sichuan Agricultural University, Chengdu 611130, China
Invertebrate fauna as the main soil fauna is one of the important biological factors affecting litter decomposition, which plays an essential role in maintaining material circulation and energy flow of terrestrial ecosystems. Alpine timberline ecotone is an important transition region of mountain vegetation vertical band with more habitat complexity and species diversity than the adjacent ecosystems. Temperature fluctuation and freeze-thaw cycle frequency in timberline was significantly higher than that in coniferous forest, in order to understand the environmental difference in timberline ecotone on the influence of invertebrate community structure and diversity in the process of litter decomposition, a field experiment using litterbag method was carried out at the end of two main periods (snow cover season and growing season), the invertebrate community structure and diversity was studied in the foliar litter ofSalixcupularis,RhododendronlapponicumandSorbusrufopilosa, three representative shrubs in the alpine timberline ecotone. The results showed that the invertebrate community diversity and density of individual and groups varied with the altitude, seasons and litter species, and invertebrate diversity influenced by seasonal variation was more significant than the altitude and litter species. The interaction of three factors(altitude, seasons and litter species) affected not only the invertebrate community diversity and evenness, but also the individual density and group density. At the end of snow cover season, Shannon indexH, Pielou indexJand Margalef indexDwas highest in the coniferous forest, and Simpson indexCwas highest in the timberline, while group density and individual density at the end of growing season was significant higher than that of snow cover season. In general, invertebrate fauna was the most abundant at the end of growing season, and relatively rich in timberline than in coniferous forest. This means that with increasing of shrub density and litter input quantity, invertebrate diversity is likely to increase in the future climate change scenarios.
invertebrate fauna; alpine ecotone; shrub; foliar litter; snow cover
國家自然科學基金資助項目(31200345, 31570605);國家科技支撐計劃資助項目(2011BAC09B05);教育部博士點基金資助項目(20115103120003);四川省教育廳重點資助項目(11ZA079)
2015- 02- 25;
日期:2015- 12- 14
10.5846/stxb201502250380
*通訊作者Corresponding author.E-mail: sicauliuyang@163.com
和潤蓮, 陳亞梅, 鄧長春, 楊林, 劉軍偉, 楊萬勤, 張健, 劉洋.川西高山林線三種灌木凋落葉分解中的無脊椎動物多樣性.生態學報,2016,36(17):5497- 5507.
He R L, Chen Y M, Deng C C, Yang L, Liu J W, Yang W Q, Zhang J, Liu Y.Invertebrate diversity in foliar litter of three shrubs in the alpine timberline of western Sichuan.Acta Ecologica Sinica,2016,36(17):5497- 5507.
