夏季若爾蓋高寒濕地水生生物群落食物網結構特征
2016-11-24 05:27:07斌陳發軍陶敏岳興建王志堅張耀光
水生生物學報 2016年2期
李 斌陳發軍陶 敏岳興建王志堅張耀光
(1. 內江師范學院生命科學學院, 長江上游魚類資源保護與利用四川省重點實驗室, 內江 642000; 2. 西南大學, 淡水魚類資源與生殖發育教育部重點實驗室, 重慶 400715)
夏季若爾蓋高寒濕地水生生物群落食物網結構特征
李 斌1陳發軍1陶 敏1岳興建1王志堅2張耀光2
(1. 內江師范學院生命科學學院, 長江上游魚類資源保護與利用四川省重點實驗室, 內江 642000; 2. 西南大學, 淡水魚類資源與生殖發育教育部重點實驗室, 重慶 400715)
為了解若爾蓋高寒濕地夏季水生生物群落食物網結構及營養關系特征, 應用穩定同位素技術分析了若爾蓋濕地 3個不同區域(ZS1、ZS2和ZS3)水生生物群落碳、氮穩定同位素比值, 并計算了δ13C—δ15N同位素生態位中的6個營養結構量化指標。結果顯示:外源性營養源陸生植物δ13C、δ15N值分別為-28.23‰— -26.07‰,-1.20‰—5.98‰; 內源性營養源顆粒有機物 (POM:主要成分為藻類)和水生植物δ13C、δ15N值分別為-26.39‰— -21.17‰、-25.37‰— -24.15‰, 3.68‰—6.61‰、3.50‰—4.01‰; 其中POM樣品δ13C、δ15N組成存在明顯的區域性差異(P < 0.05, P < 0.001)。食物網營養結構分析顯示若爾蓋濕地外源性碳源輸入(優勢陸生植物)對于維持該水域生態系統結構穩定有著十分重要的作用。若爾蓋濕地水域食物網營養級介于2—3,暗示了其生態系統結構的相對脆弱性。同位素量化指標標記發現若爾蓋濕地水生動物群落生態結構存在明顯的空間異質性, 可能與若爾蓋濕地發達的畜牧產業相關。
食物網結構; 穩定同位素; 同位素生態位; 若爾蓋高寒濕地
自然生態系統中食物網主要用于描述不同生物之間內在錯綜復雜的營養關系[1]。然而不同生物間的營養關系常因為種內與種間[2]、資源可利用性[3]、棲息環境變化[4]等因素使得食物網結構變的十分復雜。所以探討食物網結構與功能, 能夠揭示群落內部不同生物之間的關系, 可以評估和預測自然或人為干擾對生態系統“安全”產生的威脅[1]。
傳統定量研究食物網結構主要采用胃腸含物分析。隨著技術的進步, 穩定性同位素技術的出現已經成為研究水生生態系統食物網結構與功能不可或缺的手段[5—8]。生物體內天然存在的碳(δ13C)、氮(δ15N)穩定同位素比值變化可以反映生物長期消化吸收的食物來源、營養級位置和食物網結構[9]。δ15N在營養級的傳遞過程中有大約3.4‰的富集度,通常用來指示生物所處的營養級水平[10]。消費者δ13C值與其食物來源相似, 平均富集度約為0—1‰,是食物來源良好的示蹤指標[11,12]。近年來, 碳、氮穩定同位素技術的應用在我國得到了快速發展, 研究領域主要集中在湖泊[9]、海洋[13]、河流及河口[14—16]食物網結構、碳源貢獻及營養關系等方面。
若爾蓋是世界上面積最大的高原沼澤濕地, 被譽為“高原之腎”、“黃河蓄水池”, 對于保護生物多樣性、維持區域乃至全球C循環平衡以及保障地區社會經濟可持續發展具有重要作用[17]。隨著20世紀70年代初的大規模挖溝排水工程及現代旅游業的快速崛起, 若爾蓋濕地出現了一系列嚴重的環境問題, 如植被退化、水土流失、沼澤干涸、濕地面積萎縮等[18], 亟需開展人為干擾對若爾蓋濕地水生生物群落食物網結構和功能的影響研究。本研究應用碳、氮穩定同位素技術, 并結合Layman等[19]提出的形態生態學方法, 以若爾蓋濕地水域生物群落為研究對象, 討論生物群落食物能量流動和營養級構成特征, 以期為若爾蓋濕地管理和生態修復及保護提供理論支持。
1 材料與方法
1.1 研究區域概況
若爾蓋濕地位于青藏高原東北部, 地理位置101°36′—103°30′E, 32°20′—34°00′N。若爾蓋濕地海拔3600—3800 m, 區域內地貌以低山、丘陵、閉流寬谷地和湖群洼地為主[20]。降水多、濕度大、霜凍期長, 屬于高原寒溫帶濕潤氣候, 年平均降水量為660—750 mm, 5—10月為雨季, 降水占全年的90%; 年平均氣溫為1.2℃, 7—8月份平均氣溫為10.9℃(圖 1、表 1)。

圖 1 若爾蓋濕地采樣圖Fig. 1 Locations of sampling areas in Zoige wetland
1.2 樣品的采集與處理
徒手采集若爾蓋濕地水域沿岸優勢陸生植物,用清水除去表面附著物。POM(水中顆粒有機物,經過鑒定主要成分為浮游藻類)采集方法如下:取上層水, 先通過浮游動物網過濾后水樣經預燒的玻璃纖維濾膜(GF/C Whatman)過濾獲得含有顆粒有機物樣品濾膜用錫箔紙包裹, 存放于封口袋中(每一個POM樣品至少由3個相應的樣混合而成)。利用三角形手抄網(100目)采集龍虱Cybister sp.、鉤蝦Gammarid和小劃蝽Sigara substriata sp.等大型底棲無脊椎動物。徒手采集軟體動物泉膀胱螺Physa fontinalis, 鑒定后將每個類群分別放置于充分曝氣的蒸餾水中過夜, 使其腸含物排空。每個采樣點采集的泉膀胱螺樣品至少由100個個體混合而成, 去除其外殼獲取軟組織, 其他幾種底棲無脊椎動物取其整體作為實驗材料(每個類群的數量均能滿足同位素測試需要的量)。魚類樣品通過手抄網獲得,取其背側肌肉; 所有樣品通過移動冰箱運回到實驗室于-20℃低溫保存備用。用于碳穩定性同位素分析的濾膜先用1 mol/L的鹽酸處理, 然后用蒸餾水沖洗干凈。所有樣品在60℃烘干至恒重, 用研缽研磨成均一粉末, 放入干燥器保存待測。
1.3 同位素測試
所有樣品的穩定C和N同位素比值均在北京林業科學院同位素實驗室測定, 穩定同位素質譜儀為菲尼根Flash EA112 HT元素分析儀與菲尼根DELTA V Advantage同位素比率質譜儀相連而成。穩定C、N 同位素的自然豐度表示為:
δX=([R樣品/R標準]-1)×1000‰
式中X是13C或15N, R是13C/12C或15N/14N。δ13C值是相對于國際PDB標準, δ15N值是相對于空氣氮氣的豐度。分析精度δ13C < 0.10‰, δ15N < 0.20‰。
1.4 數據統計與分析
營養級計算公式[1]:
TP (Trophic position)=[ (δ15Nconsumer-δ15Nbaseline)/ Δδ15N‰]+λ 式中, Δδ15N=3.4‰; 當baseline是生產者時, λ=1; 當baseline是初級消費者時, λ=2。
參考了Layman等[20]的形態生態學方法中用于量化營養結構的6個指標:氮值范圍NR(δ15N rang),碳值范圍CR(δ13C rang), 總面積TA(Total area), 平均離心距離CD (Mean distance to centroid), 平均最鄰近距離NND (Mean nearest neighbor distance), 最鄰近距離標準差SDNND(Standard deviation of nearest neighbor distance)。其中TA計算采用ArcGIS 9.3軟件完成, 其余指標計算均在Excel 2003軟件中完成。方差分析在STATISTICA.6.0軟件中完成, 其他數據統計分析在SPSS16.0軟件中完成。

表 1 采樣點簡介Tab. 1 The characteristics of each sampling site in Zoige wetland
2 結果
2.1 碳(δ13C)、氮(δ15N)同位素值組成
表 2顯示了若爾蓋濕地3個樣區(ZS1、ZS2和ZS3)不同生物樣品δ13C、δ15N值(表 2、表 3)。研究區域外源性碳源主要由7種優勢陸生植物組成(表2), 其δ13C值范圍為-28.23‰— -26.07‰, 平均值為(-27.11±0.58)‰, 其中以ZS1的鵝絨委陵菜Potentilla anserina δ13C值最大, ZS3的舌狀花Cichorioideae δ13C值最小; 內源性碳源主要由POM(主要成分為浮游藻類)和水生植物(水毛茛)構成, 方差(ANOVAs)
分析顯示不同樣區之間POM樣品δ13C值差異顯著(F=14.56, P=0.045), 最大值出現在ZS1, 平均值為(-22.43±0.85)‰, ZS2最小, 平均值為(-26.31±0.04)‰。在消費者中不同種類δ13C值變幅較寬, 從-30.86‰(高原鰍 Triplophysa sp)至-22.30‰(泉膀胱螺)差異達到了8.56‰。浮游動物枝角類Cladocera δ13C值在若爾蓋濕地不同樣區間差異不明顯(ANOVAs,F=3.40, P=0.10), 但軟體動物泉膀胱螺δ13C值差異極為顯著(F=73.45, P<0.000), 其大小順序依次為ZS3>ZS1>ZS2(表 2)。
外源性碳源δ15N值為-1.2‰(ZS1, 鵝絨委陵菜)至6.61‰(ZS2, POM), 差值達到了7.18‰。POM樣品δ15N值在不同樣區間變化趨勢與其 δ13C值不同(ANOVAs, F=171.1, P<0.001), 其大小依次為ZS2>ZS3>ZS1。消費者中以高級捕食者黃河裸裂尻Schizopygopsis pylzovi δ15N值最高, 平均值為(10.62±0.19)‰(ZS3), 浮游動物最低, 平均值為(4.08±0.31)‰(ZS3)。不同樣樣區間浮游動物和軟體動物δ15N值差異均極顯著(ANOVAs, F=63.29,P<0.000; F=44.18, P<0.001), 其大小順序依次是ZS2>ZS3>ZS1, ZS3>ZS1>ZS2 (表 3)。總體而言魚類的δ15N值高于無脊椎動物。
2.2 營養結構

表 2 若爾蓋濕地不同生物類群δ13C值Tab. 2 The values of carbon isotope of the different organisms communities in Zoige wetland
依據不同生物類群δ13C、δ15N值構建了ZS1、ZS2和ZS3樣區的二維營養結構模型圖(圖 2—4)。圖 2和圖 4中大部分消費者δ13C介于外源性碳源禾本科、扁蓄蓼Polygonum aviculare 、鵝絨委陵菜、披堿草Elymus dahuricus和茼蒿Chrysanthemum coronarium sp.等δ13C范圍內。圖 3顯示(ZS2樣區)多數消費者δ13C值都超出了內源性碳源(POM和水生植物)和外源性碳源(沿岸陸生植物)δ13C值的范疇。

表 3 若爾蓋濕地不同生物類群δ15N值Tab. 3 The values of nitrogen of the different organisms communities in Zoige wetland
營養關系是了解生態系統能量流動的核心, 也是群落賴以生存的基礎。消費者和食物之間的氮穩定性同位素分餾值伴隨著營養級增加δ15N值不斷增加。由于初級生產者δ15N的變化可以影響到對食物網中、高級生物營養級的判斷, 通常采用生態系統中食性相對簡單、常年存在的初級消費者作為基線生物[1]。本研究中泉膀胱螺符合此要求,故采用泉膀胱螺的δ15N值作為計算營養級的基線值, 分別計算了若爾蓋濕地不同樣區水生動物群落食物網中的營養級水平(表 4), 其中ZS1食物網營養級長度達到了3級, ZS2接近3級, 而ZS3僅有2.4個營養級(圖 4)。若爾蓋濕地中, ZS2的浮游動物2.93± 0.09和高原鰍2.79±0.32的營養級均高于ZS1和ZS3。
2.3 消費者生態位空間結構

圖 2 ZS1中不同生物類群δ13C-δ15N雙坐標圖Fig. 2 The bi-plots of stable isotope based on species collected from ZS1
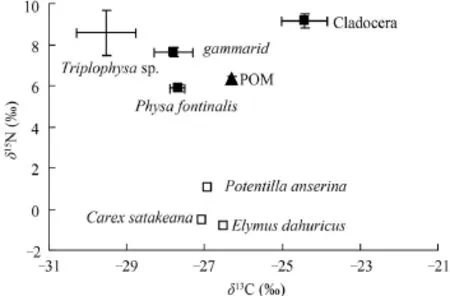
圖 3 ZS2中不同生物類群δ13C-δ15N雙坐標圖Fig. 3 The bi-plots of stable isotope based on species collected from ZS2
本研究采用的“δ13C-δ15N”組成的二維生態位,并結合Layman[20]的同位素營養結構量化指標比較了若爾蓋濕地水域3個不同樣點(ZS1、ZS2和ZS3)消費者生態位空間結構(圖 5—7)。ZS3動物δ13C-δ15N總面積(TA值)較ZS1和ZS2為大(TA=13.54,圖 7), 表明了在ZS3TA值所代表的消費者群落的生態位總空間較ZS1和ZS2大。ZS1的氮值范圍NR值>ZS2>ZS3, 但其碳值范圍 CR值<ZS2<ZS3, 說明了ZS1的營養級長度較ZS2和ZS3長, 但在食物網基礎食物源多樣性方面卻低于它們。ZS3樣區的平均離心距離(CD值)最大(CD=2.67, 圖7), 顯示了其營養多樣性程度最高。通過兩個樣點間平均最近鄰距離(NND值)的大小可知, ZS3消費者營養位置差異最大, 而ZS1消費者的最近鄰距離標準差(SDNND)最大, 說明在該樣點消費者營養級分化程度相對于ZS2和ZS3而言更為均勻。
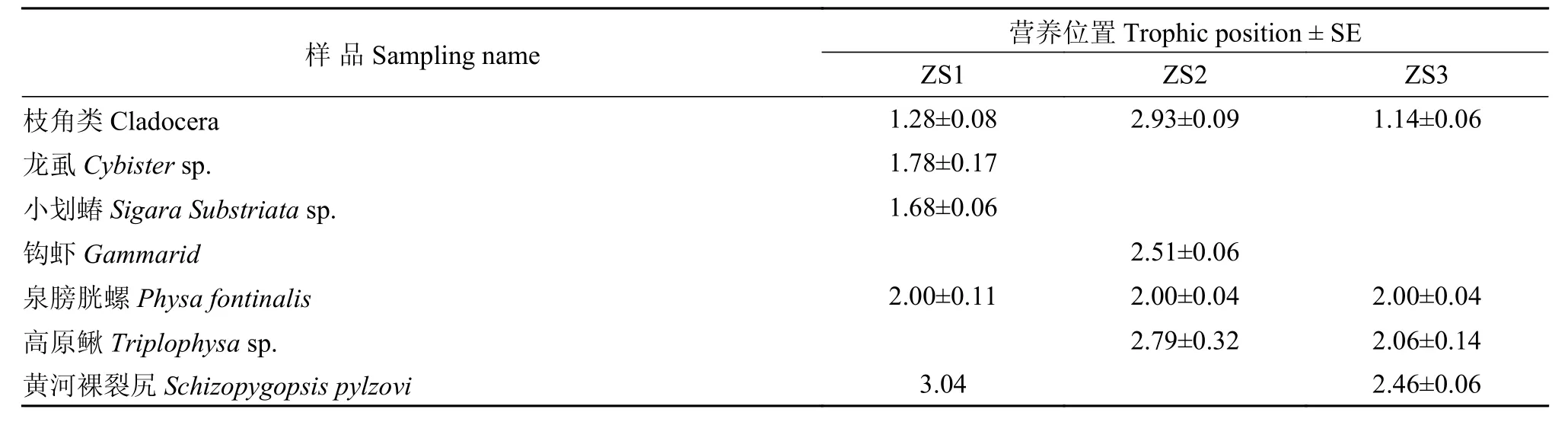
表 4 若爾蓋濕地物種多樣性生物相對營養級Tab. 4 The relative trophic position of selective organism
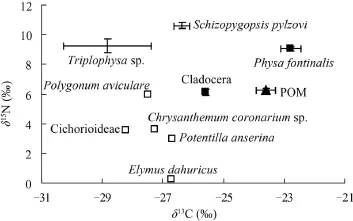
圖 4 ZS3中不同生物類群δ13C-δ15N雙坐標圖Fig. 4 The bi-plots of stable isotope based on species collected from ZS3
3 討論
3.1 若爾蓋濕地初級生產者同位素特征
若爾蓋濕地外源性碳源的碳同位素δ13C值分布在一個相對狹窄的范圍-28.23‰— -26.07‰, 介于李明財[21]已報道的青藏高原東部高寒草甸陸生植物δ13C值范圍內(-29.2‰— -24.9‰)。水生植物和POM(主要成為為藻類)δ13C值與其他水生生態系統研究中的相關報道相近[22,23]。與碳穩定同位素信號不同, 若爾蓋濕地所有初級生產者氮穩定同位素信號均存在明顯的空間異質性可能主要與該區域畜牧業發達有關[24]。
3.2 若爾蓋濕地食物網能流途徑

圖 5 若爾蓋濕地ZS1點主要消費者營養結構Fig. 5 The trophic structure of the main consumers in ZS1 from Zoige wetland
在食物網的研究中, 既要關注食物網資源與消費者穩定性同位素值的特征, 又要重點理解消費者營養生態位的組成, 尤其是消費者種類組成變化在維持水生生態系統穩定性中扮演的重要角色, 決定了食物鏈長度和物質能量的傳遞效率[25]。若爾蓋濕地水域生態環境安全問題嚴重, 生態健康狀況呈現逐年退化趨勢[18], 關注若爾蓋濕地水域生物群落種類、食物網能量流動及營養級結構組成特征有重要意義。
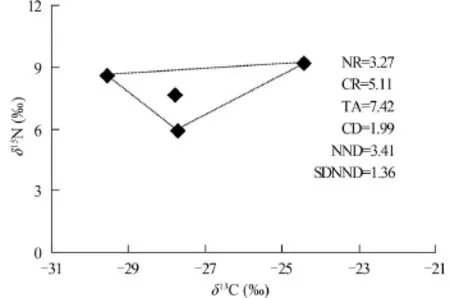
圖 6 若爾蓋濕地ZS2點主要消費者營養結構Fig. 6 The trophic structure of the main consumers in ZS2 from Zoige wetland
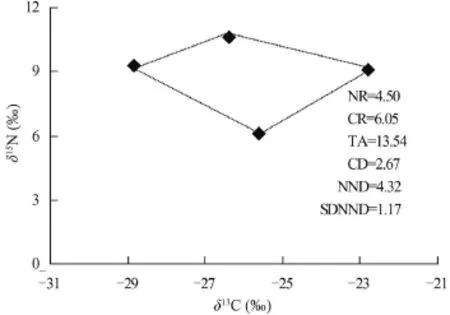
圖 7 若爾蓋濕地ZS3點主要消費者營養結構Fig. 7 The trophic structure of the main consumers in ZS3 from Zoige wetland
在若爾蓋濕地3個樣區中, ZS1和ZS3營養結構模型圖(圖 2、圖 4)表明了外源性營養源(陸生植物)是它們食物網能量的重要基礎碳源。而ZS2(圖3)大多數消費者能量可能來源于δ13C值更為貧乏的營養源。上述結果表明若爾蓋濕地外源性碳源(沿岸優勢陸生植物)對于維持該區域水域生態系統食物網結構的穩定性有著十分重要的作用, 這一結果與眾多湖泊和河流食物網基礎食物源主要來源于內源性碳源(藻類)的結論明顯不同[26—28], 這一現象可能與若爾蓋高寒濕地生態系統地理位置和氣候條件有關。分析其原因, 首先分布在若爾蓋濕地水域沿岸帶優勢陸生植物, 在每年若爾蓋雨(6—8月份)季洪水淹沒后可以為水生動物提供了多樣的生境; 其次長時間浸泡在水中的陸生植物慢慢腐爛分解形成豐富的營養源。
若爾蓋濕地水生動物種類組成的多樣性相對較為單一。此次調查僅采集到無脊椎動物和魚類種類均較少, 尤其是魚類(黃河裸裂尻和高原鰍)。研究中魚類在不同樣區(ZS1、ZS2和ZS3)的食物網中均位于食物鏈的頂端(圖 2—4)。從營養級水平來看, 若爾蓋濕地水域食物鏈長度介于2—3級, 與湖泊平均4.0個營養級[29]相比少了1—2個營養級, 這也暗示了若爾蓋濕地水域生態系統食物網結構的相對脆弱性。
3.3 基于Layman指標評價效果分析
理論上物種的生態位會受到人為干擾、生物多樣性、種群大小及捕食關系等多種因素的影響[30]。當生態系統中食物源缺乏時, 物種的生態位幅度降低, 反之則增大[31]。應用穩定同位素研究群落水平的營養關系是近年來研究的一個焦點, 尤其是定量評價技術手段在分析群落數據中的應用[32]。本研究采用了營養結構量化的6個指標數據評價了若爾蓋濕地不同樣區之間消費者生態位空間結構。總體而言, 若爾蓋濕地不同樣區間水生動物群落生態結構存在明顯的空間異質性, 可能與當地發達的畜牧業有關。研究結果表明了該方法在分析若爾蓋濕地不同水域食物網水生動物生態結構空間異質性方面有明顯優勢。與δ13C-δ15N直觀圖(圖 2—4)的趨勢特征相比, Layman指標給出了定量評價若爾蓋濕地生態系統中不同動物生態位空間差異的結果, 也為進一步探究若爾蓋濕地水域生態系統的穩定性和抵抗力提供了一種定量判別方法。
4 結論
夏季若爾蓋高寒濕地生態系統中外源性碳源(沿岸陸生優勢植物)輸入對于維持該區域食物網結構的穩定性有著十分重要的作用。從營養級水平來看, 若爾蓋濕地水域食物鏈長度介于2—3級, 與湖泊平均4.0個營養級相比少了1—2個營養級, 表明若爾蓋濕地水域生態系統食物網結構的相對脆弱性。比較發現, Layman的6個量化指標方法可以為進一步探究若爾蓋濕地水域生態系統的穩定性和抵抗力提供一種定量判別方法。
致謝:
感謝內江師范學院生命科學學院本科生解崇友、倪露蕓、張靜、吳迪、楊燕等同學在樣品的采集和處理過程中給予的幫助,內江師院許斌老
[1]Post D M. Using stable isotopes to estimate trophic position: models, methods, and assumptions [J]. Ecology,2002, 83(3): 703—718
[2]Zimmerman M S, Schmidt S N, Krueger C C, et al. Ontogenetic niche shifts and resource partitioning of lake trout morphotypes [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2009, 66(6): 1007—1018
[3]López-Bao J, Palomares F, Rodríguez A, et al. Intraspecific interference influences the use of prey hotspots [J]. Oikos, 2011, 120(10): 1489—1496
[4]Schindlere D E, Scheuerell M D. Habitat coupling in lake ecosystems [J]. Oikos, 2002, 98(2): 177—189
[5]Boecklen W JM, Yarnes C T, Cook B A, et al. On the use of stable isotopes in trophic ecology [J]. Annual Review of Ecology Evolution and Systematics, 2011, 42(1): 411—440
[6]Monroy M, Maceda Veiga A, Caiola N, et al. Trophic interactions between native and introduced fish species in a littoral fish community [J]. Journal of Fish Biology, 2014,85(5): 1693—1704
[7]Rao W R, Ning J J, Zhong P, et al. Size-dependent feeding of omnivorous Nile tilapia in a macrophyte-doninated lake: implications for lake management [J]. Hydrobiologia, 2015, 749(1): 125—134
[8]Costas N, Pardo I. Isotopic variability in a stream longitudinal gradient: implications for trophic ecology [J]. Aquatic Sciences, 2015, 77(2): 231—260
[9]Xu J. Ecological studies on the food web structures and trophic relationships of freshwater lakes in China using stable carbon and nitrogen isotopes [D]. Institute of Hydrobiology, Chinese Academy of Sciences. 2005 [徐軍.應用碳、氮穩定性同位素探討淡水湖泊的食物網結構與營養級關系. 博士論文, 中國科學院生物研究水生所. 2005]
[10]DeNiro M J, Epstein S. Influence of diet on the distribution of nitrogen in animals [J]. Geochimica et Cosmochimica Acta, 1978, 42(5): 341—355
[11]Gu B, Schelske C L, Hoyer M V. Stable isotopes of carbon and nitrogen as indicators of diet and trophic structure of the fish community in a shallow hypereutrophic lake [J]. Journal of Fish Biology, 1996, 49(6): 1233—1243
[12]Persic A, Roche H, Ramade F. Stable carbon and nitrogen isotope quantitative structural assessment of dominant species from the Vaccares lagoon trophic web (Cammargu Biosphere Reserve, France) [J]. Estuarine, Coastal and Shelf Science, 2004, 60(2): 261—272師在數據計算方面給予的幫助,中國林業科學院穩定性同位素實驗室柯淵老師對同位素測定提供的幫助。
[13]Yang G H, Hou X Q, Sun X L, et al. Constructi on food web model of Liusha Bay-using stable isotope analysis results [J]. Acta Hydrobiologica Sinica, 2013, 37(1): 150—156 [楊國歡, 侯秀瓊, 孫省利, 等. 流沙灣食物網結構的初探——基于穩定同位素方法的分析結果. 水生生物學報, 2013, 37(1): 150—156]
[14]Deng H T, Ba J W, Duan X B, et al. The analysis of the trophic levels of the major fish species in the Daning River using stable isotope technology [J]. Acta Hydrobiologica Sinica, 2015, 39(5): 893—901 [鄧華堂, 巴家文,段辛斌, 等. 運用穩定同位素技術分析大寧河主要魚類營養層級. 水生生物學報, 2015, 39(5): 893—901]
[15]Mao Z G, Gu X H, Zeng Q F, et al. Food web structure of a shallow eutrophic lake (Lake Taihu, China) assessed by stable isotope analysis [J]. Hydrobiologica, 2012, 683(1): 173—183
[16]Quan W M. Food web analysis of salt marshes of the Yangtze River Estuary by using stable isotopes [D]. Fudan University, Shanghai. 2007 [全為民. 長江口鹽沼濕地食物網的初步研究:穩定同位素分析. 上海: 復旦大學. 2007]
[17]Han D Y, Yang Y X, Yang Y, et al. Species composition and succession of swamp vegetatin along grazing gradients in the Zoige Plateau, China [J]. Acta Ecology Sinica,2011, 31(20): 5946—5955 [韓大勇, 楊永興, 楊楊, 等. 放牧干擾下若爾蓋高原沼澤濕地植被種類組成及演替模式. 生態學報, 2011, 31(20): 5946—5955]
[18]Yang Y X, Wang S Y. Human disturbances on mire and peat soils in the Zoige Plateau [J]. Resources Science,2001, 23(2): 37—41 [楊永興, 王世巖. 人類活動干擾對若爾蓋高原沼澤土、泥炭土資源影響的研究. 資源科學, 2001, 23(2): 37—41]
[19]Layman C A M, Arrington D A, Montafia C G, et al. Can stable isotope ratios provide for community-wide measures of trophic structure [J]? Ecology, 2007, 88(1): 42—48
[20]Wang C K, Wang Y S, Zhang A D, et al. Wetland resources and its protection in Zoige Plateau [J]. Bulletin of Soil and Water Conservation, 2001, 21(5): 20—22 [王長科, 王躍思, 張安定, 等. 若爾蓋高原濕地資源及其保護對稱. 水土保持通報, 2001, 21(5): 20—22]
[21]Li M C, Li Z F, Yi X F, et al. Annual variation of plant δ13C and its environmental analysis in alpine meadow in the east of Tibetan Plateau [J]. Ecology and Environment,2007, 16(4): 1205—1210 [李明財, 黎貞發, 易現峰, 等.青藏高原東部高寒草甸植物δ13C年間變化及環境分析.生態環境, 2007, 16(4): 1205—1210]
[22]Raven J A, Walker A D I, Johson M, et al. Implications of δ13C natural abundance measurements of photosynthetic performance by marine macrophytes in their natural environment [J]. Marine Ecology Progress Series, 1995,123(1—3): 193—205
[23]Yoshioka T, Wada E, Hayashi H. A stable isotope studyon seasonal food web dynamics in a eutrophic lake [J]. Ecology, 1994, 75(3): 835—846
[24]Anderson C, Cabana G. Does δ15N in river food webs reflect the intensity and origin of N loads from the watershed [J]. Science of the Total Environment, 2006,367(2—3): 968—978
[25]Vander Zanden M J, Fetzer W W. Global patterns of aquatic food chain length [J]. Oikos, 2007, 116(8): 1378—1388
[26]Lewis W MJr, Hamilton S K, Rodríguez M A. Foodweb analysis of the Orinoco floodplain based on production estimates and stable isotope data [J]. Journal of the North American Benthological Society, 2001, 20: 241—254
[27]Hamilton S K, Lewis W M Jr, Sippel S J. Energy sources for aquatic animals in the Orinoco River floodplain: evidence from stable isotopes [J]. Oecologia, 1992, 89(3): 324—330
[28]Bunn S E, Davies P M, Mosisch T D. Ecosystem measures of river health and their response to riparian and catchment degradation [J]. Freshwater Biology, 1999,41(2): 333—345
[29]Wangtzen K M, Machado F A, Voss M. Seasonal isotopic shiftis of the Pantanal wetland, Brazil [J]. Aquatic Sciences, 2002, 64(3): 239—251
[30]Fox B J. Niche parameters and species richness [J]. Ecology, 1981, 62(6): 1415—1425.
[31]Winemiller K O, Planka E R, Vitt L J Food web laws or niche theory? Six independent empirical tests [J]. The American Naturalist, 2001, 158(2): 193—199
[32]Solomon C T, Carpenter S R, Rusak J A, et al. Long-term variation in isotopic baselines and implications for estimating consumer trophic niches [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2008, 65(10): 2191—2200
THE CHARACTERISTIC OF THE FOOD WEB STRUCTURE AT AQUATIC ORGANISMS COMMUNITY IN SUMMER OVER ZOIGE APLINE WETLAND
LI Bin1, CHEN Fa-Jun1, TAO Min1, YUE Xing-Jian1, WANG Zhi-Jian2and ZHANG Yao-Guang2
(1. Key Laboratory of Sichuan Province for Fishes Conservation and Utilization in the Upper Reaches of the Yangtze River, School of Life Sciences, Neijiang Normal University, Neijiang 642000, China; 2. Key Laboratory of Freshwater Fish Reproduction and Development (Ministry of Education), Southwest University, Chongqing 400715, China)
Zoige Apline wetland is the largest plateau in the world and plays important roles in the biodiversity conservation, regional and global carbon circulation balance and social sustainable development. In order to understand the relationship of food web structure and the trophic characteristics of aquatic organisms community in Zoige Apline wetland, stable isotopes techniques were used to analyze the carbon and nitrogen stable isotope ratios of different aquatic organisms community at 3 sampling areas in Zoige Apline wetland and calculate 6 trophic structure measurable index in δ13C—δ15N isotopic niche. The result showed that the δ13C and δ15N values of exogenous nutrition source (land plant)were -28.23‰ — -26.07‰ and -1.20‰ — 5.98‰ respectively. The δ13C and δ15N values of endogenous nutrition source POM (POM is short for particulate organic matter and it mainly consisted of algae) were -26.39‰ — -21.17‰and -25.37‰ — -24.15‰ respectively. The δ13C and δ15N values of aquatic plants were 3.68‰ — 6.61‰ and 3.50‰ —4.01‰ respectively. The δ13C and δ15N values of POM had a spatial dependent pattern (P<0.05, P<0.001). The result of food web structure analysis revealed that the carbon (advantaged land plant around Zoige Apline wetland) played a key role in the foundation of the food web structure for the aquatic ecosystem. The food web trophic level between 2 and 3 implied the instability of the ecosystem structure. The result of isotope measurable indexes demonstrated significant spatial differences among the animal community ecosystem structure of the different sampling areas, which might associated with the developed graziery of Zoige Apline wetland.
Food web structure; Stable isotope; Isotopic niche; Zoige Apline wetland
10.7541/2016.42
S932.2
A
1000-3207(2016)02-0313-08
2015-04-21;
2015-09-14
四川省教育廳重點項目(14ZA0251); 內江師范學院校級重點項目(13ZA02)資助 [Supported by the Key Fund Project of Sichuan Provincial Department of Education (14ZA0251); the Key Project of Neijiang Normal University (13ZA02)]
李斌(1981—), 男, 河南信陽人; 博士, 副教授; 主要從事水域生態學研究。E-mail: libin2004xinyang@126.com
張耀光(1955—), 男, 四川蓬溪人; 教授博士生導師; 主要從事水生生物學研究。E-mail: zhangyg@swu.edu.cn
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
今日農業(2021年11期)2021-08-13 08:53:34
今日農業(2020年20期)2020-12-15 15:53:19
中華詩詞(2019年7期)2019-11-25 01:43:04
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
瞭望東方周刊(2017年34期)2017-09-13 17:13:26
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
發明與創新(2016年16期)2016-08-21 13:56:16
發明與創新(2016年21期)2016-05-17 03:57:29
