飼料中添加裂殖壺菌粉對凡納濱對蝦脂質(zhì)沉積和血清生化成分的影響
2016-11-24 05:26:55鐘國防焦建剛華雪銘李玲玉劉嘉航
水生生物學(xué)報 2016年2期
關(guān)鍵詞:血清
鐘國防 焦建剛 華雪銘 李玲玉 劉嘉航
(上海海洋大學(xué)農(nóng)業(yè)部水產(chǎn)種質(zhì)資源與利用重點開放實驗室, 上海 201306)
飼料中添加裂殖壺菌粉對凡納濱對蝦脂質(zhì)沉積和血清生化成分的影響
鐘國防 焦建剛 華雪銘 李玲玉 劉嘉航
(上海海洋大學(xué)農(nóng)業(yè)部水產(chǎn)種質(zhì)資源與利用重點開放實驗室, 上海 201306)
以健康的凡納濱對蝦(Litopenaeus Vannamei)[初重(5.81±0.24) g]為研究對象, 研究裂殖壺菌粉(Extractedand-dried microfungi Schizochytrium sp., EDMS)對凡納濱對蝦脂質(zhì)轉(zhuǎn)運、沉積以及血清生化成分的影響。在飼料中分別添加0、0.5%、1.0%、1.5%和2.0%的裂殖壺菌粉, 制成5組(基礎(chǔ)組、EMDS05、EMDS10、EMDS15、EMDS20)等氮等能飼料。將750尾健康的凡納濱對蝦隨機分成5組(為每組3個平行, 每個平行45尾蝦),養(yǎng)殖時間45d。結(jié)果表明, 隨著飼料中DHA水平的提高, 肌肉中水分、粗脂肪和粗蛋白質(zhì)含量均無顯著改變(P>0.05), 但灰分隨著添加量的增加呈現(xiàn)上升趨勢, 且EMDS20組顯著高于基礎(chǔ)組和EMDS05組(P<0.05);肌肉和肝胰臟中高不飽和脂肪酸(HUFA)含量顯著升高(P<0.05), 血清總甘油三酯(TG)含量顯著升高(P<0.05), 總膽固醇(TC)含量保持穩(wěn)定(P>0.05), EDMS15組高/低密度脂蛋白(HDL/LDL)顯著高于其他各組(P<0.05);超氧化物歧化酶(SOD)呈現(xiàn)先下降后升高的趨勢, 其中EMDS10組(1.0%添加組)顯著低于其他各組(P<0.05);丙二醛(MDA)含量顯著升高(P<0.05)。綜上所述, 飼料中添加EDMS促進了凡納濱對蝦肌肉中n-3 HUFA的沉積,同時也提高了蝦體脂質(zhì)過氧化壓力。
凡納濱對蝦; 裂殖壺菌; 高不飽和脂肪酸; 脂質(zhì)過氧化
脂類作為重要的營養(yǎng)素, 除為魚類提供生長發(fā)育所需的脂肪及必需脂肪酸以外, 還是脂溶性維生素吸收的良好載體[1]。魚油是配合飼料脂類的主要來源, 近年來魚油產(chǎn)量的下降, 減少或者替代魚油的研究成為熱點, 玉米油、亞麻油和椰子油等在魚類飼料中的應(yīng)用研究有見報道[2]。但植物油脂缺乏花生四烯酸 (Arachidonic acid, ARA)、二十碳五烯酸(Eicosapentaenoic acid, EPA)和二十二碳六烯酸(Docosahexaenoic acid, DHA)等長鏈不飽和脂肪酸(Long-chain unsaturated fatty acid, LC-UFA), 長期大量使用會影響魚蝦脂肪酸組成。Kim等[3]用不同的植物油脂替代魚油飼養(yǎng)羅氏沼蝦(Macrobrachium rosenbergii)發(fā)現(xiàn), 植物油實驗組肌肉中DHA沉積顯著低于魚油組。甲殼動物由于缺乏合成長鏈脂肪酸的能力, 只能通過食物獲取[4], 因而飼料中對高不飽和脂肪酸(Highly unsaturated fatty acid, HUFA)有更高的需求, 要通過添加含有HUFA的藻類、菌類等[5—7]或者合成DHA等才能滿足需求。微藻、菌類因具有天然抗脂質(zhì)過氧化體系, 是植物油脂替代魚油時補充高度不飽和脂肪酸的良好選擇[8]。
凡納濱對蝦(Litopenaeus vannamei)是當(dāng)前世界上養(yǎng)殖產(chǎn)量最高的三大優(yōu)良品種之一, 它具有生長快、抗病能力強等優(yōu)點, 養(yǎng)殖經(jīng)濟效益顯著。從2001年以來, 我國養(yǎng)殖南美白對蝦面積和產(chǎn)量一直居于對蝦養(yǎng)殖的首位, 其養(yǎng)殖量已達到我國對蝦養(yǎng)殖量的70%以上, 同時也已成為世界上養(yǎng)殖規(guī)模最大產(chǎn)量最高的對蝦品種, 具有極高的經(jīng)濟價值。裂殖壺菌(Schizochytrium limacinum)是屬于破囊壺菌科(Thraustochytrium)的一類海洋真菌, 菌體含70%的油脂(90%以上為甘油三酯), 油脂中含有30%以上的DHA[9], 是生物發(fā)酵生產(chǎn)DHA的工程菌。裂殖壺菌體具有抗氧化活性, 能防止油脂氧化[10], 飼料中添加裂殖壺菌能促進斑點叉尾(Lctalurus punctatus)[11,12]、海鯛(Sparus aurata)[13]、大西洋鮭(Salmo salar)[14]等魚類的生長。裂殖壺菌作為生物餌料的營養(yǎng)強化劑或直接飼喂幼體[15,16]也能取得良好的效果。本實驗以凡納濱對蝦為研究對象, 研究裂殖壺菌對其脂肪轉(zhuǎn)運及沉積和血清生化成分的影響, 為凡納濱對蝦功能性飼料的開發(fā)提供數(shù)據(jù)和理論支持。
1 材料與方法
1.1 實驗飼料
飼料配方及營養(yǎng)組成見表 1。參考Samocha等[17]對凡納濱對蝦DHA需求量的研究, 分別按0、0.5%、1.0%、1.5%和2.0%的水平加入裂殖壺菌粉(Extracted-and-dried microfungi Schizochytrium sp.,EDMS: 粗蛋白質(zhì)19.26%, 粗脂肪33.71%, 水分0.94%, 灰分8.98%, DHA 18%), 配制成基礎(chǔ)組、EDMS05組、EDMS10組、EDMS15組和EDMS20組5種實驗飼料, 飼料中DHA的含量分別為: 19.5、42.1、59.7、80.1和93.1 mg/kg (實測值)。飼料原料購自上海農(nóng)好飼料有限公司, 原料經(jīng)粉碎后過80目篩, 用絞肉機制粒(粒徑1.2 mm, 粒長5 mm), 風(fēng)干后-20℃儲存?zhèn)溆谩?/p>
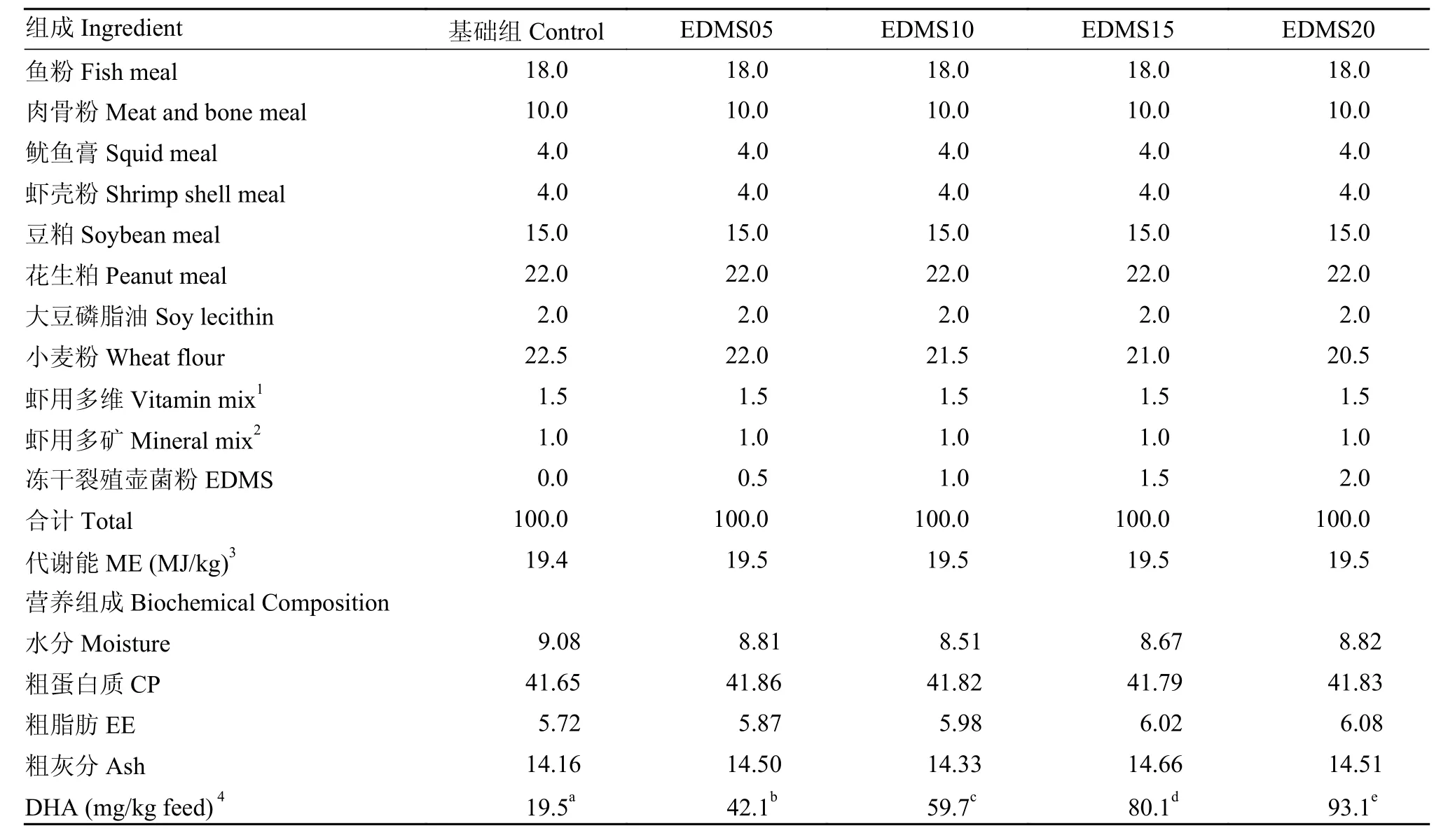
表 1 飼料配方及營養(yǎng)組成表(風(fēng)干基礎(chǔ))Tab. 1 Ingredients and proximate compositions of experimental diets (DM basis, %)
1.2 養(yǎng)殖管理
選擇健壯且體型勻稱的對蝦在上海海洋大學(xué)濱海養(yǎng)殖基地的水泥池(5 m×2 m×1.5 m)中暫養(yǎng)兩周, 稱取750尾蝦[均重: (5.81±0.24) g]隨機放入15個網(wǎng)箱(2 m×1.2 m×1.2 m)中, 每個網(wǎng)箱45尾。實驗共分5個實驗組, 每組3個平行。每天投喂兩次(05:00和18:00), 日投飼量為蝦體重的3%—5%, 每兩周取樣稱重調(diào)整投飼量, 每周測定水質(zhì)指標(biāo)并換水1/3, 養(yǎng)殖期間水溫21—28℃, 鹽度為(4.2±0.2)‰,溶解氧4.5—12 mg/L, 氨氮0—0.2 mg/L, 亞硝酸鹽0—0.1 mg/L。養(yǎng)殖實驗持續(xù)45d[18]。
1.3 樣品采集
養(yǎng)殖試驗結(jié)束后, 停食24h取樣, 每尾蝦稱重后,每平行取10尾蝦的血淋巴液, 用肝素鈉(125 U/mL)防止凝固。同一平行的血淋巴液混合放入兩個2 mL離心管中, 離心(10min, 8000 r/min, 4℃)分離血清備用。取肝胰臟和肌肉置于-20℃冰箱中保存待測。
1.4 測定指標(biāo)和方法
血清生化指標(biāo)測定: 總甘油三酯(Total triglycerides, TG, 氧化酶法)、總膽固醇(Total cholesterol, TC, 氧化酶法)、高/低密度脂蛋白(直接法)使用邁瑞公司(深圳, 中國)BS-300型生化分析儀測定;丙二醛(Methane dicarboxylic aldehyde, MDA, TBA法)、超氧化物歧化酶(Superoxide dismutase, SOD,黃嘌呤氧化酶法)用南京建成生物工程研究所購試劑盒測定, (酶活定義為每毫升反應(yīng)液中SOD抑制率達50%時所對應(yīng)的SOD量為1個亞硝酸鹽單位)。
肌肉營養(yǎng)成分測定: 水分測定用105℃常壓干燥法, 粗灰分測定用550℃灼燒法, 粗蛋白質(zhì)測定用杜馬斯燃燒法[19], 粗脂肪測定用氯仿-甲醇法[20], 每個樣品測3個重復(fù)。
脂肪酸測定: 在粗脂肪樣品加入濃度為0.5 mg/mL的十九烷酸(西格瑪奧德里奇公司, 美國)0.5 mL, 真空干燥后, 用三氟化硼-甲醇法進行甲酯化, 甲酯化樣品用氣質(zhì)聯(lián)用GC-MS測定(7890A GC和5975C GC/MS, 安捷倫公司, 美國)。色譜柱為非極性毛細管柱(L=30 m, ID=0.32 mm, DF=0.25 μm), 載氣為氦氣(純度≥99.99%)。升溫程序: 40—170℃,100℃/min;170—240℃, 20℃/min;進樣量1 μL。通過面積歸一法計算不同脂肪酸在總脂肪酸中的含量來確定脂肪酸的組成;通過不同脂肪酸甲酯的峰面積與十九烷酸內(nèi)標(biāo)物的峰面積比值計算粗脂肪中各脂肪酸在干物質(zhì)中的含量[某種脂肪酸含量m(單位: mg)=(S×M)/(Sc19×Mc19)×mc19/m0, S為某種脂肪酸峰面積, M為某種脂肪酸甲酯摩爾質(zhì)量,Sc19為十九烷酸內(nèi)標(biāo)的峰面積, Mc19為十九烷酸甲酯的摩爾質(zhì)量, m0為樣品質(zhì)量]。
1.5 數(shù)據(jù)處理
用SPSS 17.0處理數(shù)據(jù), 用單因子方差分析(one-way ANOVA, LSD)和Duncan氏多重比較檢驗統(tǒng)計結(jié)果。所有結(jié)果使用平均值±標(biāo)準差(mean±SD)表示。以P<0.05為顯著水平。
2 結(jié)果
2.1 飼料中EDMS水平對凡納濱對蝦肌肉常規(guī)營養(yǎng)成分的影響
凡納濱對蝦肌肉成分如表 2所示, 隨著配合飼料中EDMS水平逐漸升高, 肌肉灰分呈現(xiàn)上升趨勢,且EMDS20組顯著高于基礎(chǔ)組和EMDS05組(P<0.05)。但水分、粗蛋白質(zhì)和粗脂肪各實驗組間無顯著差異(P>0.05)。
2.2 飼料中EDMS水平對凡納濱對蝦肌肉脂肪酸組成的影響

表 2 飼料中EDMS水平對凡納濱對蝦肌肉常規(guī)營養(yǎng)成分的影響Tab. 2 Effects of dietary EDMS on muscle composition of L. vannamei (%)
由表 3可知, 在45d養(yǎng)殖實驗后, 肌肉總脂肪中C18:0在各組中的比例呈現(xiàn)先下降后上升的趨勢,EDMS10組顯著低于EDMS05和EDMS20組(P<0.05), 但與基礎(chǔ)組無顯著差異(P>0.05);C18:2n6含量呈現(xiàn)逐漸下降的趨勢, 且基礎(chǔ)組顯著低于實驗組(P<0.05);EDMS10和EDMS15組的DHA含量顯著高于基礎(chǔ)組。EDMS20組的總飽和脂肪酸(Satura-ted fatty acid, SFA)顯著高于EDMS10組(P<0.05),但總多不飽和脂肪酸(Polyunsaturated fatty acid,PUFA)含量顯著低于EDMS10組(P<0.05);EDMS10組的HUFA顯著高于基礎(chǔ)組(P<0.05);n-3/n-6呈現(xiàn)先上升后下降的趨勢, 且EDMS10、EDMS15和EDMS20組顯著高于基礎(chǔ)組(P<0.05)。

表 3 飼料中EDMS水平對凡納濱對蝦肌肉脂肪酸組成的影響(%,占總脂肪酸)Tab. 3 Effects of dietary EDMS on muscle fatty acids composition of L. vannamei (% of total fatty acids)
2.3 飼料中EDMS水平對凡納濱對蝦肌肉脂肪酸含量的影響
由表 4可知, 在45d養(yǎng)殖實驗后, 在肌肉脂肪酸含量中, EDMS20組的C16:0、C17:0、C18:0、C18:1n9、C20:5n3和C22:6n3含量均顯著高于其他各組(P<0.05);EDMS10和EDMS15實驗組的C18:2n6、C20:5n3和C22:6n3均顯著高于基礎(chǔ)組(P<0.05)。HUFA和n-3/n-6含量隨添加量的增加逐漸增加, 各添加組n-3/n-6均顯著高于基礎(chǔ)組(P<0.05),但EDMS10組、 EDMS15和 EDMS20之間沒有顯著性差異(P>0.05)。
2.4 飼料中EDMS水平對凡納濱對蝦肝胰臟脂肪酸組成的影響
由表 5可知, 肝胰臟脂肪酸隨著飼料中EDMS水平的上升, 呈先升高后降低的有: C17:0、 C18:0、C20:1n9, 峰值均出現(xiàn)在EDMS15組, 并且EDMS15組顯著高于基礎(chǔ)組(P<0.05);呈先下降后上升的脂肪酸有: C18:1n9、C18:2n6、C18:2n3, 且EDMS15組的C18:2n6顯著低于基礎(chǔ)組(P<0.05);呈逐漸上升趨勢的脂肪酸有: C22:5n3, 且EDMS15組和EDMS20組顯著高于基礎(chǔ)組(P<0.05);呈逐漸下降趨勢的脂肪酸有: C20:5n3、C20:4n6, 且EDMS20組顯著低于基礎(chǔ)組(P<0.05);隨著飼料中EDMS水平的上升, SFA呈現(xiàn)逐漸上升的趨勢,EDMS20組顯著高于基礎(chǔ)組(P<0.05), n-3/n-6隨著添加量的增加逐漸升高, 各實驗組均顯著高于基礎(chǔ)組(P<0.05);PUFA和HUFA均呈現(xiàn)逐漸下降的趨勢, 且EDMS20組顯著低于基礎(chǔ)組(P<0.05)。
2.5 飼料中EDMS水平對凡納濱對蝦肝胰臟脂肪酸含量的影響
由表 6可知, 隨著飼料中EDMS添加量的上升,肝胰臟脂肪酸中C16:0、C16:1n7、C17:0、C20:1n9、C20:2n6、C22:5n3呈先下降后上升的趨勢, 谷值均出現(xiàn)在EDMS10組且顯著低于EDMS20(P<0.05);C20:5n3、C20:4n6呈現(xiàn)逐漸上升的趨勢, 但各實驗組與基礎(chǔ)組間沒有顯著性差異(P>0.05)C22:6n3也呈上升趨勢, 且EDMS20的組顯著高于其他各組(P<0.05);EDMS20組的SFA、PUFA、HUFA和TFA均顯著高于其他各組(P<0.05), 而n-3/n-6在各實驗組之間沒有顯著性差異(P>0.05)。
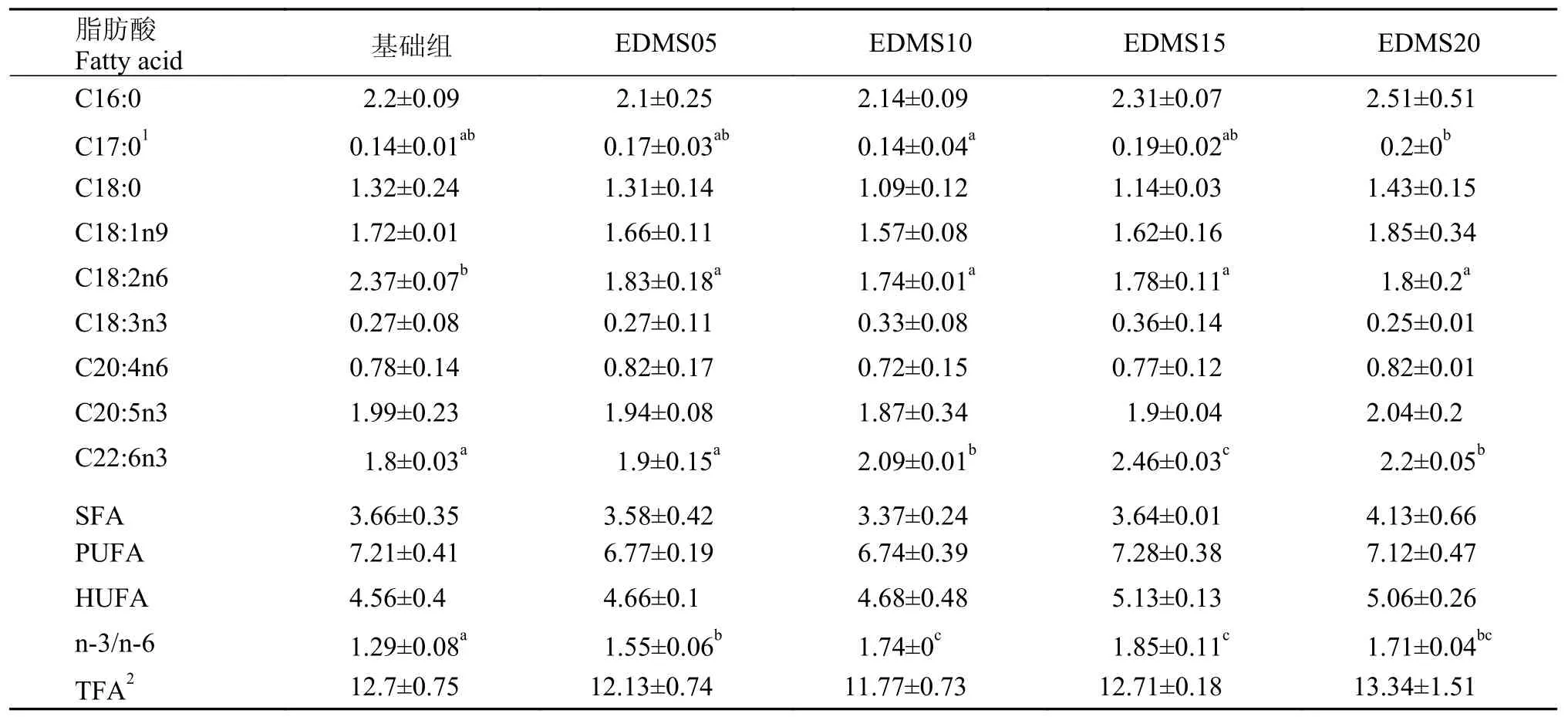
表 4 飼料中EDMS水平對凡納濱對蝦肌肉脂肪酸含量的影響Tab. 4 Effects of dietary EDMS on muscle fatty acids content of L. vannamei (mg/g dried muscle)
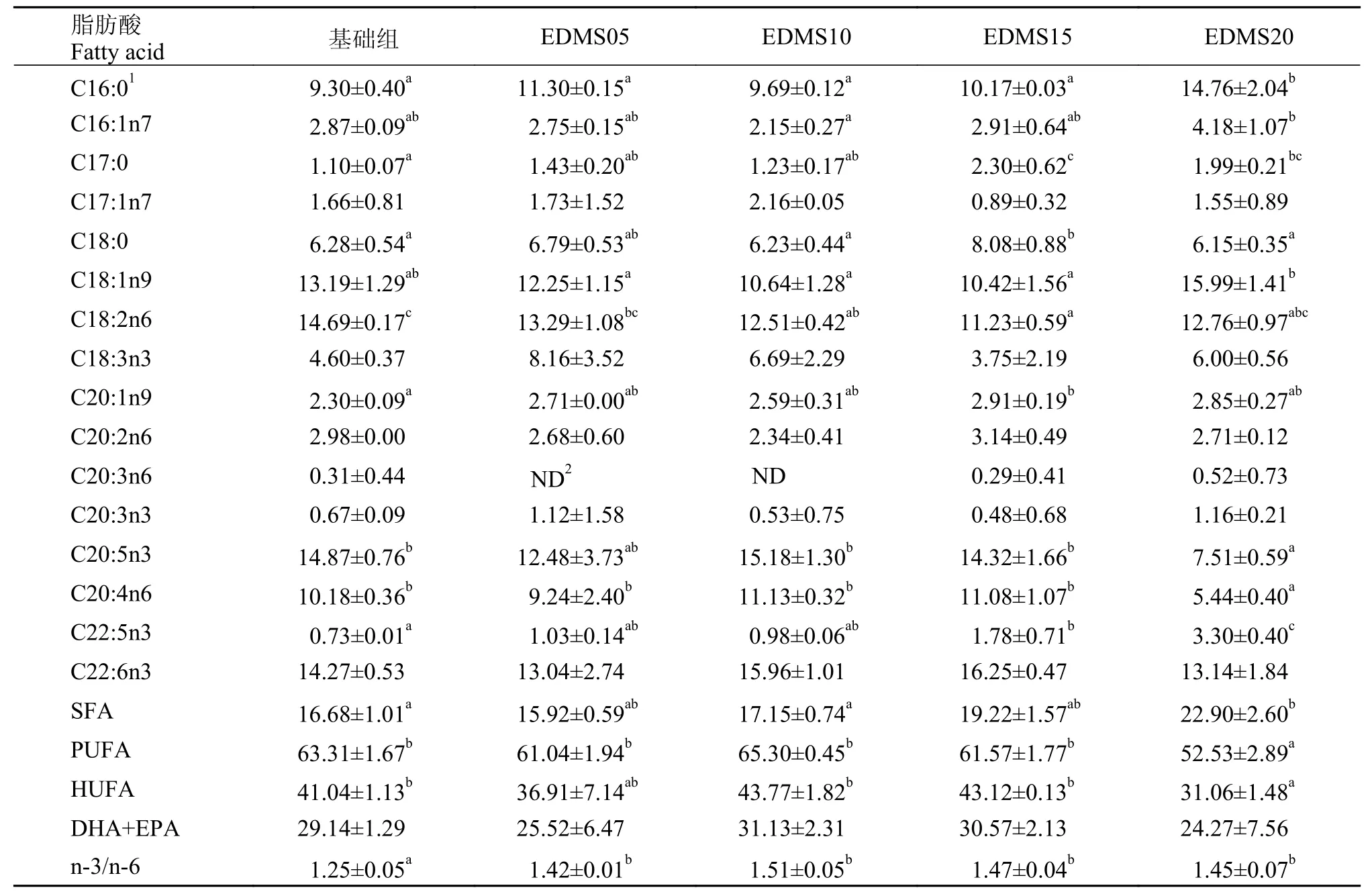
表 5 飼料中EDMS水平對凡納濱對蝦肝胰腺脂肪酸組成的影響(%, 占總脂肪酸)Tab. 5 Effects of dietary EDMS on Hepatopacreas fatty acids composition of L. vannamei (% of total fatty acids)
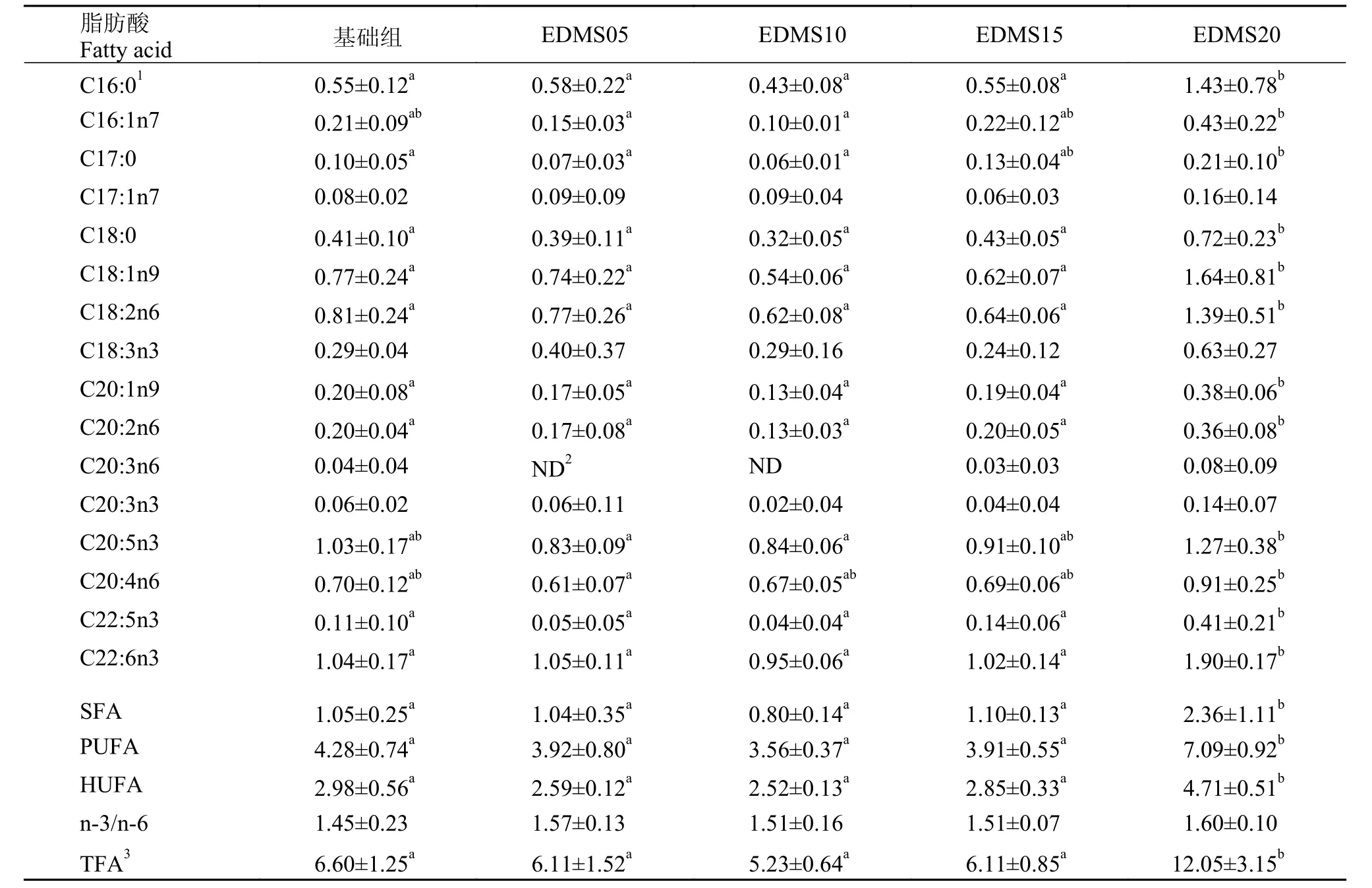
表 6 飼料中EDMS水平對凡納濱對蝦肝胰臟脂肪酸含量的影響(毫克每克肝胰臟,mg/g)Tab. 6 Effects of dietary EDMS on hepotopancreas fatty acids content of L. vannamei (mg/g wet hepatopancreas)
2.6 飼料中EDMS水平對凡納濱對蝦血清生化成分的影響
由表 7可知, 凡納濱對蝦血清中TG含量隨著EDMS水平的升高而升高, EDMS20顯著高于EDMS05(P<0.05), 但各實驗組與基礎(chǔ)組無顯著差異(P>0.05);高/低密度脂蛋白呈現(xiàn)先上升后下降的趨勢, 且EDMS10組和EDMS15組顯著高于基礎(chǔ)組(P<0.05);TC含量各實組之間無顯著差異(P>0.05);血清中MDA含量在EDMS10組出現(xiàn)顯著上升, 在EDMS20達到最大值, 隨后稍有下降, 且EDMS10組、EDMS15組和EDMS20組顯著高于EDMS05組和基礎(chǔ)組(P<0.05);SOD活性呈現(xiàn)先下降后升高的“V”型曲線關(guān)系, 在EDMS15組有一個快速上升過程, 且EDMS20組顯著高于EDMS10組(P<0.05)。
3 討論
3.1 EDMS水平對凡納濱對蝦脂肪沉積的影響
HUFA是甲殼動物的必需脂肪酸, 飼料中不同的脂肪酸比例會導(dǎo)致甲殼動物肌肉中脂肪酸的積累出現(xiàn)差異。Forster等[21]用不同水平的豆粕替代魚粉后, 比較魚油和經(jīng)亞麻油酸強化的豆油對凡納濱對蝦肌肉脂肪酸沉積的影響, 發(fā)現(xiàn)魚油被強化豆油替代后, 低水平魚粉組(19%和0)凡納濱對蝦全蝦和肌肉中的n-3 HUFA含量顯著下降(P<0.05), 但肌肉總脂肪含量并無顯著變化(P>0.05)。González-Félix等[22]研究飼料中不同磷脂水平與不同油脂類型對凡納濱對蝦肌肉脂肪酸組成的影響, 結(jié)果表明肌肉中ARA、EPA和DHA含量與飼料中HUFA含量呈高度正相關(guān)性(P<0.05)。在本實驗中, 隨著裂殖壸菌粉(EDMS)添加量的增加, 肌肉粗脂肪含量無顯著差異, 但HUFA含量逐漸升高, 與González-Félix等[22]研究結(jié)果相似。而在脂肪酸組成中,HUFA的含量呈先升高后下降趨勢, 表明HUFA在肌肉脂肪酸中的相對積累速度與脂肪總量的沉積并不同步。

表 7 飼料中EDMS水平對凡納濱對蝦血清生化成分的影響Tab. 7 Effects of dietary EDMS on serum biochemical indicators of L. vannamei
甲殼動物肝胰臟既是脂肪的主要貯存部位, 又是脂肪代謝的中心器官, 其脂肪酸組成與肌肉中的脂肪酸組成有差異[23]。Sui等[24]用氫化植物油替代魚油養(yǎng)殖中華絨螯蟹(Eriocheir sinensis)發(fā)現(xiàn), 隨著替代量的增加, 中華絨螯蟹肝胰臟中EPA和DHA含量顯著下降(P<0.05), 但對肝胰臟粗脂肪含量沒有顯著影響(P>0.05)。而本實驗中肝胰臟粗脂肪含量隨著EDMS添加量的增加先下降后升高, 與此同時HUFA含量與在脂肪酸中所占比例也同步呈現(xiàn)逐漸下降趨勢, 與在肌肉中HUFA含量與在脂肪酸中所占比例的差異不同的原因可能是肝胰臟的高代謝頻率使脂肪酸在肝胰臟中的積累較為平衡。
PUFA是類二十烷酸合成的底物, n-6 PUFA合成促炎性類二十烷酸, 而n-3 PUFA合成抗炎性類二十烷酸, 兩者具有相反的生理學(xué)功能[25]。甲殼動物中, n-3 HUFA比n-6 HUFA對機體生長的促進作用更為顯著, 但增加ARA在必需脂肪酸中的比例可以提高甲殼動物的存活率[26]。適宜的n-3/n-6值對凡納濱對蝦生長代謝具有重要意義。Hu等[27]發(fā)現(xiàn)通過調(diào)節(jié)魚油、豆油和大豆磷脂油的不同配比來調(diào)整飼料中n-3/n-6的比值, 結(jié)果對養(yǎng)殖的凡納濱對蝦肌肉中的n-3/n-6比值產(chǎn)生了影響, 當(dāng)肌肉中的n-3/n-6比值在1.5—2.0范圍內(nèi)時, 凡納濱對蝦的生長和存活率均無顯著差異。在本實驗中, 肌肉中n-3/n-6為1.20—1.68時, 凡納濱對蝦的生長(相對增重率88%—92%)和存活率(66%—69%)均無顯著差異,且EDMS的添加提高了n-3 PUFA在肌肉中的比例,提升了對蝦的食用價值, 對人類健康有積極作用。
3.2 EDMS水平對凡納濱對蝦脂肪轉(zhuǎn)運的影響
脂肪分為膽固醇、中性脂(甘油三酯)、磷脂和游離脂肪酸。凡納濱對蝦不能在體內(nèi)合成HUFA,需要從飼料中攝取。血淋巴是脂肪吸收和轉(zhuǎn)運的樞紐, 甘油三酯和游離脂肪酸作為能源物質(zhì)通過血淋巴在脂肪組織和肝臟之間進行轉(zhuǎn)運。長鏈脂肪酸在血淋巴內(nèi)的運輸以甘油三酯為主, 且DHA可以促進脂肪分解代謝, 促使甘油三酯從肝臟運至組織被代謝供能。González-Félix研究發(fā)現(xiàn)在添加等量磷脂的條件下, 提高DHA的添加量可以提高凡納濱對蝦幼蝦血清甘油三酯的含量[28];膽固醇作為生物體重要的功能性物質(zhì), 是對蝦生長發(fā)育不可缺少的脂類物質(zhì), 主要通過脂蛋白在血淋巴中進行轉(zhuǎn)運。參加膽固醇轉(zhuǎn)運的高密度脂蛋白和低密度脂蛋白具有相反的生理學(xué)功能, 高密度脂蛋白將血淋巴膽固醇運入肝臟而低密度脂蛋白反之, 兩者比例的升高或降低表示血液內(nèi)的膽固醇是進入肝臟參與代謝活動還是從肝臟運輸?shù)浇M織中沉積下來[29,30]。在本研究中, 血清總甘油三酯含量隨EDMS的增加顯著提高(P<0.05), 表明長鏈脂肪酸在血清中的含量增加;總膽固醇隨著EDMS的增加稍有下降但并不顯著, 而HDL/LDL呈現(xiàn)先上升后下降的趨勢,說明適量的EDMS能促進血液向肝臟輸送膽固醇,避免膽固醇在血管中的堆積, 提高膽固醇的利用效率, 且平衡血淋巴內(nèi)的膽固醇在穩(wěn)定水平。
3.3 EDMS水平對凡納濱對蝦脂質(zhì)過氧化反應(yīng)的影響
HUFA因為含有3個以上的雙鍵, 極易受到超氧陰離子的攻擊而發(fā)生脂質(zhì)過氧化反應(yīng)。MDA是脂質(zhì)過氧化產(chǎn)物, 它在血清中的含量表示體內(nèi)脂質(zhì)過氧化程度。Yang 等[31]研究飼料中添加氧化魚油對凡納濱對蝦生長和氧化應(yīng)激的影響, 認為添加50—100 g/kg的氧化魚油顯著抑制了對蝦增重率和特定生長率, 且血清、肌肉中的MDA含量顯著上升(P<0.05)。生物體內(nèi)有天然的抗氧化機制, 如SOD能將超氧陰離子(O2-)通過歧化反應(yīng)生成O2和
[1]Yue Y F, Peng S M, Shi Z H, et al. The effects of fatty acid nutrition on the growth, lipid metabolism and immune performance of the fish [J]. Modern Fisheries Information, 2012, 26(11): 13—19 [岳彥峰, 彭士明, 施兆鴻, 等. 脂肪酸營養(yǎng)對魚類生長, 脂代謝及免疫性能影響的研究進展. 現(xiàn)代漁業(yè)信息, 2012, 26(11): 13—19]
[2]Chen J L, Han D, Zhu X M, et al. Dietary lipid sources for gibel carp (Carassius auratus gibelio): Growth performance, tissue composition and muscle fatty acid profiles [J]. Acta Hydrobiologica Sinica, 2011, 33(6): 988—997 [陳家林, 韓冬, 朱曉鳴, 等. 不同脂肪源對異育銀鯽的生長、體組成和肌肉脂肪酸的影響. 水生生物學(xué)報, 2011, 33(6): 988—997]
[3]Kim Y C, Romano N, Lee K S, et al. Effects of replacing die- tary fish oil and squid liver oil with vegetable oils on the growth, tissue fatty acid profile and total carotenoids of the giant freshwater prawn, Macrobrachium rosenbergii [J]. Aquaculture Research, 2013, 44(11): 1731—1740
[4]Tocher D R. Fatty acid requirements in ontogeny of marine and freshwater fish [J]. Aquaculture Research, 2010,41(5): 717—732
[5]Alhazzaa R, Bridle A R, Nichols P D, et al. Replacing dietary fish oil with Echium oil enriched barramundi with C18PUFA rather than long-chain PUFA [J]. Aquaculture,2011, 312(1): 162—171
[6]Zhou J S, Bente E. Torstensen, Ingunn Stubhaug. Oleic acid trans-membrane uptake in hepatocytes of Atlantic salmon (Salmo salar L.) and effect of replacing dietary H2O2, 再通過過氧化氫酶分解H2O2, 不斷清除體內(nèi)超氧陰離子(O2-), 減少它攻擊不飽和脂肪酸, 從而減弱了脂質(zhì)過氧化反應(yīng)。Zhang 等[32]研究不同的脂肪添加量對凡納濱對蝦血清SOD活力和MDA含量的影響, 結(jié)果表明隨著飼料中脂肪含量的升高,MDA在血清中的含量顯著上升, 且與飼料脂肪酸中DHA的含量呈顯著正相關(guān)(P<0.05)。在本實驗中, 隨著飼料中DHA含量的上升, 血清甘油三酯和MDA含量顯著升高(P<0.05), 而SOD活力先下降后恢復(fù), 表明DHA含量的增加給機體帶來了顯著的脂質(zhì)過氧化壓力。
動物脂代謝是一個極為復(fù)雜的過程, 也是現(xiàn)階段水產(chǎn)動物營養(yǎng)學(xué)的研究熱點之一[33], 作為一種潛在的高不飽和脂肪酸的添加劑來源, 裂殖壺菌可以改變凡納濱對蝦對脂類的代謝, 并有利于提高其生產(chǎn)價值, 但菌粉作為一種細胞結(jié)構(gòu)其成分是復(fù)雜的,對脂代謝的調(diào)節(jié)作用還需要更多的后續(xù)實驗加以確定, 并為其作為水產(chǎn)動物營養(yǎng)可以使用的一種添加劑提供必要的生產(chǎn)驗證和理論背景。fish oil with vegetable oil [J]. Acta Hydrobiologica Sinica, 2014, 38(1): 121—128 [周繼術(shù), Bente E. Torstensen, Ingunn Stubhaug. 飼料中植物油替代魚油對大西洋鮭肝細胞油酸跨膜吸收的影響. 水生生物學(xué)報,2014, 38(1): 121—128]
[7]Zhu L Y, Zhang X C, Song X J, et al. Advances in n-3 polyunsaturated fatty acids DHA and EPA [J]. Marine Science, 2007, 31(11): 78—85 [朱路英, 張學(xué)成, 宋曉金,等. n-3 多不飽和脂肪酸 DHA, EPA 研究進展. 海洋科學(xué), 2007, 31(11): 78—85]
[8]Sargent J R, McEvoy L A, Bell J G. Requirements,presentation and sources of polyunsaturated fatty acids in marine fish larval feeds [J]. Aquaculture, 1997, 155(1): 117—127
[9]Yaguchi T, Tanaka S, Yokochi T, et al. Production of high yields of docosahexaenoic acid by Schizochytrium sp. strain SR21 [J]. Journal of the American Oil Chemists’ Society, 1997, 74(11): 1431—1434
[10]Pan B F, Li Z Y. Production of a single cell oil which is highly enriched in DHA by marine microalgae [J]. China Biotechnology, 2000, 20(6): 43—45 [潘冰峰, 李祖義. 利用海洋微藻生產(chǎn)富含 DHA 的單細胞油脂. 中國生物工程雜志, 2000, 20(6): 43—45]
[11]Li M H, Robinson E H, Tucker C S, et al. Effects of dried algae Schizochytrium sp., a rich source of docosahexaenoic acid, on growth, fatty acid composition, and sensory quality of channel catfish Ictalurus punctatus [J]. Aquaculture, 2009, 292(3): 232—236
[12]Faukner J, Rawles S D, Proctor A, et al. The effects of diets containing standard soybean oil, soybean oil enhanced with conjugated linoleic acids, menhaden fish oil, or an algal docosahexaenoic acid supplement on channel catfish performance, body composition, sensory evaluation,and storage characteristics [J]. North American Journal of Aquaculture, 2013, 75(2): 252—265
[13]Ganuza E, Benítez-Santana T, Atalah E, et al. Crypthecodinium cohnii and Schizochytrium sp. as potential substitutes to fisheries-derived oils from seabream (Sparus aurata) microdiets [J]. Aquaculture, 2008, 277(1): 109—116
[14]Miller M R, Nichols P D, Carter C G. Replacement of fish oil with thraustochytrid Schizochytrium sp. L oil in Atlantic salmon parr (Salmo salar L) diets [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2007, 148(2): 382—392
[15]Arungamol S, Areechon N, Yongmanitchai W, et al. Application of microalgae Schizochytrium limacinum for the nursing of Pacific white shrimp (Litopenaeus vannamei,Boone) larva and its effects on stress tolerance and disease resistance: Proceedings of the 45th Kasetsart University Annual Conference, Kasetsart, 29 January-1 Feb-ruary, 2008. Subject: Fisheries [Z]. Kasetsart University. 2008, 138—146
[16]Song X, Zhang X, Guo N, et al. Assessment of marine thraustochytrid Schizochytrium limacinum OUC88 for mariculture by enriched feeds [J]. Fisheries Science,2007, 73(3): 565—573
[17]Samocha T M, Davis D A, Roy L A, et al. The effect of non-marine HUFA supplementation with fish oil removal on growth and survival of the Pacific white shrimp,Litopenaeus vannamei [J]. Aquaculture Nutrition, 2011,17(5): 518—525
[18]Zhang L T, Sun Y, Chen A L, et al. Nitrification and denitrification in sediment of shrimp ponds in the late culture period [J]. Progress in Fishery Sciences, 2011, 32(1): 67—74 [張立通, 孫耀, 陳愛玲, 等. 對蝦養(yǎng)殖中后期蝦塘沉積物的硝化與反硝化作用. 漁業(yè)科學(xué)進展, 2011,32(1): 67—74]
[19]Etheridge R D, Pesti G M, Foster E H. A comparison of nitrogen values obtained utilizing the Kjeldahl nitrogen and Dumas combustion methodologies (Leco CNS 2000)on samples typical of an animal nutrition analytical laboratory [J]. Animal Feed Science and Technology, 1998,73(1): 21—28
[20]Huang K, Wu H Y, Zhu D G, et al. Effects of dietary lipid levels on growth and fatty acid composition in hepatopancreas and muscle of pacific white leg shrimp Litopenaeus vannamei [J]. Fisheries Science, 2011, 30(5): 249—255 [黃凱, 吳宏玉, 朱定貴, 等. 飼料脂肪水平對凡納濱對蝦生長, 肌肉和肝胰腺脂肪酸組成的影響. 水產(chǎn)科學(xué), 2011, 30(5): 249—255]
[21]Forster I P, Dominy W G, Obaldo L G, et al. The effect of soybean oil containing stearidonic acid on growth performance, n-3 fatty acid deposition and sensory characteristics of pacific white shrimp (Litopenaeus vannamei)[J]. Aquaculture Nutrition, 2011, 17(2): 200—213
[22]González-Félix M L, Lawrence A L, Gatlin III D M, et al. Growth, survival and fatty acid composition of juvenile Litopenaeus vannamei fed different oils in the presence and absence of phospholipids [J]. Aquaculture, 2002,205(3): 325—343
[23]Dall W. Lipid absorption and utilization in the Norwegian lobster, Nephrops norvegicus (L.) [J]. Journal of Experimental Marine Biology and Ecology, 1981, 50(1): 33—45
[24]Sui L Y, Sun H X, Wu X G, et al. Effect of dietary HUFA on tissue fatty acid composition and reproductive performance of Chinese mitten crab Eriocheir sinensis (H. Milne-Edwards) broodstock [J]. Aquaculture International, 2011, 19(2): 269—282
[25]Williams C D, Whitley B M, Hoyo C, et al. A high ratio of dietary n-6/n-3 polyunsaturated fatty acids is associated with increased risk of prostate cancer [J]. Nutrition Research, 2011, 31(1): 1—8
[26]Nonwachai T, Purivirojkul W, Limsuwan C, et al. Growth, nonspecific immune characteristics, and survival upon challenge with Vibrio harveyi in Pacific white shrimp (Litopenaeus vannamei) raised on diets containing algal meal [J]. Fish and Shellfish Immunology, 2010,29(2): 298—304
[27]Hu Y, Tan B, Mai K S, et al. Effects of dietary menhaden oil, soybean oil and soybean lecithin oil at different ratios on growth, body composition and blood chemistry of juvenile Litopenaeus vannamei [J]. Aquaculture International, 2011, 19(3): 459—473
[28]González-Félix M L, Gatlin III D M, Lawrence A L, et al. Effect of dietary phospholipid on essential fatty acid requirements and tissue lipid composition of Litopenaeus vannamei juveniles [J]. Aquaculture, 2002, 207(1): 151—167
[29]Lin X D, Xie R T, Zhong L X, et al. Effects of different lipids in dietary feed of Litopenaues vannamei juvenile prawn on its growth and composition of liver fatty acids[J]. Journal of Southern Agriculture, 2011, 42(4): 441—445 [林向東, 謝瑞濤, 鐘靈香, 等. 飼料中添加不同油脂對凡納濱對蝦幼蝦生長及肝臟脂肪酸的影響.南方農(nóng)業(yè)學(xué)報, 2011, 42(4): 441—445]
[30]Li X H, Wang B J, Wang L, et al. Biochemical characteration and molecular cloning of penaeid shrimp hemolymph lipoproteins [J]. Marine Science, 2008, 32(11): 80—83 [李曉華, 王寶杰, 王雷, 等. 對蝦血淋巴脂蛋白的生化特征及分子克隆研究進展. 海洋科學(xué), 2008,32(11): 80—83]
[31]Yang S P, Liu H L, Wang C G, et al. Effect of oxidized fish oil on growth performance and oxidative stress of Litopenaeus vannamei [J]. Aquaculture Nutrition, 2015,21(1): 121—127
[32]Zhang S, Li J, Wu X, et al. Effects of different dietary lipid level on the growth, survival and immune-relating genes expression in Pacific white shrimp, Litopenaeus vannamei [J]. Fish and Shellfish Immunology, 2013,34(5): 1131—1138
[33]Wang A M, Han G M, Feng G N, et al. Effects of dietary lipid levels on growth performance, nutrient digestibility and blood biochemical indices of Gift tilapia (Oreochromis niloticus) [J]. Acta Hydrobiologica Sinica, 2011,35(1): 81—87 [王愛民, 韓光明, 封功能, 等. 飼料脂肪水平對吉富羅非魚生產(chǎn)性能、營養(yǎng)物質(zhì)消化及血液生化指標(biāo)的影響. 水生生物學(xué)報, 2011, 35(1): 81—87]
EFFECT OF EXTRACTED AND DRIED MICROFUNGI SCHIZOCHYTRIUM IN THE DIET ON FAT DEPOSITION AND SERUM BIOCHEMICAL INDEXES OF LITOPENAEUS VANNAMEI
ZHONG Guo-Fang, JIAO Jian-Gang, HUA Xue-Ming, LI Ling-Yu and LIU Jia-Hang
(Key Laboratory of Freshwater Fishery Germplasm Resources, Ministry of Agriculture, Shanghai Ocean University, Shanghai 201306, China)
To evaluate the effects of dietary extracted-and-dried microfungi Schizochytrium sp. (EDMS) on muscle nutritional composition, fatty-acid composition of muscle and hepatopancreas, and the biochemical composition of the serum of the Litopenaeus vannamei [initial average weight of (5.81±0.24) g, mean±SE)], five isonitrogenous(41.65%—41.86% CP, crude protein) and isocaloric (17.6 MJ/kg) diets were formulated, containing 0 (control), 0.5%,1.0%, 1.5% and 2.0% EDMS, named as control, EDMS05, EDMS10, EDMS15 and EDMS20, respectively. Each diet was randomly fed to triplicate groups of 45 shrimps per cage for 45d. The results showed that EDMS has no effect on crude lipid and crude protein of muscle (P>0.05). EDMS increased the ash level and EMDS20 has significantly higher ash than EMDS05 (P<0.05). EDMS significantly increased the percentage and content of the essential fatty acids in muscle and hepatopancreas (P<0.05). The total triglyceride of serum was remarkably increased (P<0.05) by adding EDMS and the total cholesterol of serum did not change significantly (P>0.05) by adding EDMS. The HDL/LDL ratio in EDMS15 was significantly higher than that of the other groups (P<0.05). EDMS decreased superoxide dismutase(SOD) and EMDS10 has the strongest effect (P<0.05). EDMS significantly increased the Methane Dicarboxylic Aldehyde (MDA) in serum (P<0.05). These results suggest that EDMS can improve n-3 HUFA deposition in the muscle of the white shrimp and increase a risk of lipid peroxidation.
Litopenaeus vannamei; Schizochytrium limacinum; Highly unsaturated fatty acid; Lipid peroxidation
10.7541/2016.32
S963.73
A
1000-3207(2016)02-0225-10
2015-03-26;
2015-08-18
公益性行業(yè)(農(nóng)業(yè))專項: 淡水池塘工程化改造與環(huán)境修復(fù)技術(shù)研究與示范(201203083); 水產(chǎn)動物遺傳育種中心上海市協(xié)同創(chuàng)新中心(ZF1206); 上海海洋大學(xué)科技發(fā)展專項基金(A2-0209-15-200057); 上海市科技興農(nóng)重點攻關(guān)項目(滬農(nóng)科攻字(2014)第3—8號)資助 [Supported by Special Fund for Agro-scientific Research in the Public Interest, State Agriculture Ministry of China(201203083); Shanghai University Knowledge Service Platform Project (ZF1206); Shanghai Ocean University Funding Program for Development of Science and Technology (A2-0209-15-200057); Shanghai Science and Technology Key Project for Promoting Agriculture]
鐘國防(1974—), 男, 湖南臨武人; 副教授; 研究方向為動物營養(yǎng)與飼料科學(xué)、池塘生態(tài)養(yǎng)殖技術(shù)。E-mail: gfzhong@shou. edu.cn
華雪銘(1974—), 女, 副教授; 研究方向為動物營養(yǎng)與飼料科學(xué)。E-mail: xmhua@shou.edu.cn
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫(yī)科大學(xué)學(xué)報(2021年2期)2021-03-29 05:31:08
昆明醫(yī)科大學(xué)學(xué)報(2021年1期)2021-02-07 01:06:36
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
昆明醫(yī)科大學(xué)學(xué)報(2020年12期)2021-01-26 00:44:04
中華養(yǎng)生保健(2020年4期)2020-11-16 01:31:40
中西醫(yī)結(jié)合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業(yè)科學(xué)(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫(yī)學(xué)院學(xué)報(2015年5期)2015-12-05 08:22:29
