氨基酸的內源性合成限制了動物的生長、泌乳和繁殖(Ⅰ)
2016-11-07 06:46:50侯永清姚康印遇龍伍國耀
中國飼料 2016年9期
侯永清,姚康,印遇龍,,伍國耀,3
(1.動物營養與飼料科學湖北省重點實驗室,動物營養與飼料安全湖北省協同創新中心,武漢輕工大學,湖北武漢43000;2中國科學院亞熱帶農業生態研究所,湖南長沙410000;3.德克薩斯農工大學動物科學系,美國德克薩斯州001210)
氨基酸的內源性合成限制了動物的生長、泌乳和繁殖(Ⅰ)
侯永清1,姚康2,印遇龍1,2,伍國耀1,3
(1.動物營養與飼料科學湖北省重點實驗室,動物營養與飼料安全湖北省協同創新中心,武漢輕工大學,湖北武漢43000;2中國科學院亞熱帶農業生態研究所,湖南長沙410000;3.德克薩斯農工大學動物科學系,美國德克薩斯州001210)
中國豬營養國際論壇是由美國動物科學學會、上海亙泰實業集團和上海優久生物科技有限公司聯合主辦,以“凝聚全球科研力量,驅動豬業創新思維”為宗旨,力邀全球一流的機構、專家和學者,傾力打造一個動物營養領域具有國際性、前沿性和權威性的論壇。該論壇每兩年舉辦一屆,聚焦行業發展中的熱點、難點,通過專家學者和企業領導者之間進行開放建設性的學術探討、理論研究和實踐經驗交流,整合全球動物營養領域最新的技術和研究成果,推動行業發展,創造和提升產業價值。
www.asaschina.org
氨基酸(AAs)是蛋白質的組成部分。丙氨酸、天冬酰胺、天冬氨酸、谷氨酸、谷氨酰胺、甘氨酸、脯氨酸、絲氨酸等八種氨基酸是所有動物都能合成的,而精氨酸的從頭合成是大多數哺乳動物的種屬特異性行為(如人類、豬和大鼠)。動物體內可合成的氨基酸傳統上被歸類為動物的營養性非必需氨基酸,因為它們的合成量足夠。然而,以下證據不支持這個假設,這些證據顯示:(1)當日糧中不含精氨酸、谷氨酸、谷氨酰胺時,盡管提供了足夠的其他氨基酸,但大鼠的生長緩慢;(2)當飼喂NRC推薦的滿足蛋白質和氨基酸要求的玉米-豆粕型日糧,而不添加精氨酸、谷氨酸、谷氨酰胺、甘氨酸、脯氨酸時,豬不能達到最大的生長、泌乳或繁殖性能;(3)當在日糧中添加谷氨酸、谷氨酰胺、甘氨酸、脯氨酸時,雞的瘦肉組織增加,飼料利用率提高;(4)沒有后瘤胃提供谷氨酰胺或脯氨酸時,泌乳奶牛不能達到最大的乳蛋白產量;(5)當日糧中不含有谷氨酰胺和脯氨酸時,魚不能達到最大生長;(6)精氨酸缺乏導致男子生精功能受損。氮代謝的定量分析表明,前體可用性和酶活性限制了動物體內氨基酸的合成。總之,這些研究結果支持的結論是,氨基酸的內源性合成限制動物的生長、泌乳和繁殖。
蛋白質;腸道;營養;代謝;健康
氨基酸(AAs)是合成蛋白質和低分子量物質(如一氧化氮、多胺、肌酸、5-羥色胺、多巴胺、和還原型谷胱甘肽)的前體,在生理學上有重要意義。動物以細胞和組織特異性的形式合成丙氨酸、天冬酰胺、天冬氨酸、谷氨酸、谷氨酰胺、甘氨酸、脯氨酸、絲氨酸的碳骨架和完整分子,但不能合成胱氨酸、組氨酸、異亮氨酸、亮氨酸、賴氨酸、蛋氨酸、苯丙氨酸、蘇氨酸、色氨酸、酪氨酸、纈氨酸的碳骨架。精氨酸的從頭合成有種屬特異性,大多數哺乳動物(如人、豬、牛、羊、鼠和大鼠)通過腸腎軸由谷氨酸、谷氨酰胺合成精氨酸,然而由于腸道中酶(如吡咯啉-5-羧酸合成酶等)的表達缺乏,鳥類和一些哺乳動物(如貓和白鼬)不能合成精氨酸(Wu和Jr,1998)。與哺乳動物相比,脯氨酸在雞和某些魚類中的合成有限。這突出了氨基酸營養在動物界里的復雜性(Baker,2008;Li等,2008)。
根據以往的觀點,不能由動物細胞合成碳骨架的氨基酸被稱為“營養性必需氨基酸”(EAAs),必須由日糧提供,然而可由動物細胞合成碳骨架的氨基酸習慣上被認為在日糧中不是必要的,被歸類為“營養性非必需氨基酸”(NEAAs)(Wu,2010)。與這種長期的營養觀念不一致,最近越來越多的數據表明,動物日糧需要添加NEAAs來實現最大生長、泌乳和繁殖的遺傳潛力(Wu等,2014a、b)。本文的主要目的是定量分析由幼小、泌乳、妊娠動物日糧蛋白合成的氨基酸,以豬為主要的動物模型。
1 研究現狀
氨基酸營養在醫藥和畜牧生產上是一個迅速發展的領域。研究人員的一個主要目標是促進蛋白質合成,并減少氨的產生,生物體內高濃度的氨會產生劇毒,并會造成環境污染(Stoll和Burrin,2006)。盡管我們對必需氨基酸在營養中的重要性知之甚多,但缺乏一些關于由生物體合成氨基酸的日糧需要的信息。對豬、鼠和人類的研究表明,日糧氨基酸在小腸中經歷了廣泛的首過分解代謝,并且以細胞、組織特異性的方式進行代謝(Chen等,2007)。預計這些代謝變化可以在生理和病理條件下影響氨基酸的內源性合成以及日糧中氨基酸合成的需要。
2 日糧氨基酸在小腸及其腔內微生物中的利用
胃腸道中的蛋白酶和肽酶,由胰腺局部生產,使得日糧蛋白質水解成游離氨基酸、二肽和三肽,以供小腸內腸腔細菌的吸收。在細胞內,小肽被迅速水解為游離氨基酸。對豬的研究表明,哺乳動物腸道細胞降解日糧中大量的丙氨酸、天冬氨酸、谷氨酸、谷氨酰胺、脯氨酸、異亮氨酸、亮氨酸和纈氨酸,但不能分解天冬酰胺、甘氨酸,絲氨酸、半胱氨酸、組氨酸、賴氨酸、蛋氨酸、苯丙氨酸、蘇氨酸、色氨酸和酪氨酸(Dai等,2011;Chen等,2009)。所有這些氨基酸都以不同速率被小腸中的細菌用于進行的蛋白質合成與分解代謝(Dai等,2013、2012)。
未能被腸上皮細胞和腸道細菌分解的日糧氨基酸在運輸到微脈管系統之前進入固有膜。在這一通道中,氨基酸被黏膜和黏膜下層的多種細胞利用,包括纖維細胞、巨噬細胞、淋巴細胞、平滑肌細胞和內皮細胞(Reeds等,1996)。由于小腸內氨基酸分解代謝的途徑不同,日糧氨基酸進入門靜脈的比例差別很大。例如,給體重為8 kg豬飼喂以酪蛋白為主的飼料,56%的日糧氨基酸進入門脈循環,而10%和34%的日糧氨基酸在小腸分別被用于蛋白質的合成與分解(Wu,1998)。同樣,我們的研究結果表明,腸內的食物中,95%~97%谷氨酸和天冬氨酸,67%~70%谷氨酰胺,以及40%的精氨酸和脯氨酸被斷奶和妊娠豬的小腸分解代謝(Wu等,2010;Kim和Wu,2009)。值得注意的是,哺乳動物的小腸中利用了大量的谷氨酰胺,但沒有其他來自動脈血的氨基酸。腸道氨基酸代謝產物包括氨、瓜氨酸、精氨酸、丙氨酸、脯氨酸(Chen等,2007)。
對小腸氨基酸代謝的認識標志著我們對動物蛋白質營養理解的一個新模式的轉變。根據傳統的觀點,人們認為膳食氨基酸不能被小腸降解,并且單個必需氨基酸與賴氨酸在機體內的比例與其在腸內飲食中的比例相似,這是動物營養中長期存在的“理想蛋白質”的概念(Cuthoys和Watford,1995)。這種觀點與研究結果不一致,研究結果表明,膳食氨基酸在小腸中的物質代謝選擇性地修正了它們進入體循環被腸外組織利用的途徑。
3 膳食氨基酸在器官間的代謝
3.1一氧化氮依賴的血液流動膳食氨基酸的利用依賴于許多不同器官之間的協作。這需要足夠的血液流動以及多種細胞運輸氨基酸。血液流速部分影響了氨基酸和其他營養物質的跨組織運輸,其被一氧化氮所調節,一氧化氮是由內皮細胞釋放的主要血管擴張劑。子宮胎盤血流量的增加對胎兒的成長是非常重要的,這需要來自其母的氧氣和營養物質。此外,整個哺乳期乳腺的血流速度通過母方嚴重影響產奶基質的供應(Kim等,2012)。值得注意的是,一氧化氮是由精氨酸通過四氫生物蝶呤和NAD(P)H-依賴的一氧化氮合酶來合成的。在內皮細胞中,精氨酸通過激活GTP環水解酶I的表達來刺激四氫生物蝶呤的合成,這表明氨基酸對動物維持的全身內環境穩定和健康起著重要的調節作用。
3.2腸-腎軸的精氨酸合成如前所述,膳食中的谷氨酰胺、谷氨酸、脯氨酸以及動脈谷氨酰胺,在大多數哺乳動物腸上皮細胞中轉化為瓜氨酸和精氨酸,而瓜氨酸則被腎臟和肝外組織用于精氨酸的合成(Wu和Jr,1998)。精氨酸隨后可被機體組織利用。瓜氨酸和精氨酸的合成速度取決于生長階段,底物的可用性,以及關鍵酶類的腸道活動,如N-乙酰谷氨酸合成酶、吡咯啉-5-羧酸合成酶和脯氨酸氧化酶(Wu和Jr,1998)。
3.3腎臟中的谷氨酰胺用于調節酸堿平衡通過腎臟中氨的合成,谷氨酰胺在調節酸堿平衡上起著重要的作用,NH3結合H+以NH4+的形式存在于尿液中(Suryawan等,1998)。在酸中毒的情況下(如劇烈運動、泌乳早期、妊娠晚期、糖尿病),腎臟吸收和分解代謝的谷氨酰胺大大增加從而維持氨的合成和H+的去除。在正常的生長發育中,谷氨酰胺在氨基糖和糖蛋白的合成中是必不可少的。氨基酸和糖蛋白都與妊娠母畜(或母親)的胚胎和胎兒有很高的相關性(Wu和Kuabe,1994)。這與谷氨酰胺是人類和大多數動物(包括大鼠和雞)血漿中最豐富的氨基酸的事實相一致。
3.4由支鏈氨基酸(BCAAs)合成谷氨酰胺因為日糧谷氨酰胺大多數不進入門脈循環,血液中的谷氨酰胺主要來自腸外組織中支鏈氨基酸,葡萄糖衍生的α-酮戊二酸和NH4+的從頭合成,包括骨骼肌、心臟、肺、脂肪組織及胎盤。由于肝臟中支鏈氨基酸轉氨酶的活性有限,膳食支鏈氨基酸可供肝外組織利用,包括骨骼肌、妊娠母畜(或母親)的胎盤和哺乳母性的乳腺(Davis等,1994)。由肝外組織分泌的支鏈α-酮酸主要被肝臟用于氧化或糖異生。
3.5由羥脯氨酸合成甘氨酸母乳中極度缺乏甘氨酸卻包含大量以肽和游離形式存在的羥脯氨酸。例如,由母豬哺乳的仔豬,母乳提供的最多只有23%的甘氨酸用于組織蛋白質合成和其他代謝途徑(Wang等,2013a、b)。作為翻譯后的修飾,羥脯氨酸也可由膠原蛋白中的脯氨酸殘基形成(Melendez-Hevia等,2009)。在這些蛋白質水解后,羥脯氨酸(主要是4-羥脯氨酸)被釋放出來。羥脯氨酸在肝臟中轉化為甘氨酸,提供了內源性氨基酸以維持新生兒的生長、發育和健康(Yeung等,2015)。因此,脯氨酸分子中的碳原子和氮原子被有效分布于喂養新生兒的母乳中。同樣,正常的飲食最多提供僅30%的甘氨酸用于成人代謝(Dillon,2012)。
3.6色氨酸的分解代謝色氨酸被依次分解為色氨酸-2,3-二氧酶或吲哚胺-2,3-二氧酶和犬尿氨酸甲酰胺酶,從而在肝臟和免疫細胞中產生犬尿氨酸。通過結合其芳香烴受體,犬尿氨酸增加肥大細胞的活性,引起低血壓,并調節細胞凋亡。最近的研究結果表明,淋巴細胞中的吲哚胺-2,3-二氧酶和多種類型細胞(例如,外周血單核細胞、樹突狀細胞、間充質干細胞)是由細胞活素類誘導的,并且犬尿氨酸途徑在免疫反應中起著重要作用(NRC,1978)。因此,色氨酸能調節膳食氨基酸在腸黏膜和小腸腔內細菌中的代謝,從而影響其在腸外組織中的利用率。
4 非必需氨基酸在動物體內合成不足
氨基酸的合成需要底物(例如,營養性必需氨基酸和α-酮酸)、輔助因子和酶。在動物和人類中,因為有大量由其他氨基酸釋放出的氨,用過量的日糧蛋白質來提供一些限制性氨基酸是不明智的(Wu等,2014b)。同樣地,由于組織(尤其是骨骼肌)的退化、生長發育受損、低抗氧化能力和次優免疫力,蛋白質的消耗低于需求是令人沮喪的(Trumbo等,2002)。NRC研究表明,精氨酸和谷氨酰胺是豬的條件性必需氨基酸但未說明其他可合成氨基酸的膳食需求,然而醫學研究所并沒有考慮人體任何非必需氨基酸的膳食需要(Wu等,2004;Mavromichalis等,2001)。
5 哺乳新生兒的氨基酸代謝
雖然母乳氨基酸消化率在96%以上,但人工飼養結果表明,母豬飼喂的仔豬最多僅表現出57.5%的生長潛力(Brosnan等,2011)。在仔豬日糧中必需氨基酸含量大大超過了蛋白質合成的需要(表1)。在非必需氨基酸中,日糧中只有天冬酰胺和絲氨酸超過新生兒對蛋白質合成的需要,其他日糧非必需氨基酸不能滿足新生兒對蛋白質合成的需要。這其中天冬氨酸、谷氨酸、甘氨酸、精氨酸嚴重不足(表1)。相應的,這些氨基酸的從頭合成在母乳喂養的新生兒中有很高的活性(Wu等,2014a)。例如,在14日齡乳豬中,精氨酸、谷氨酰胺、甘氨酸合成的凈速率分別至少是每天0.58、1.15和1.20 g/kg體重(BW)(Sun等,2015)。
通過非蛋白質合成途徑供腸外利用的必需氨基酸的氮總量僅比用于合成氨基酸的氮含量高1.9%,并且用于腸外非蛋白質合成途徑的碳總量比用于合成氨基酸的碳含量高107%(表1)。可用于氨基酸合成的必需氨基酸的量甚至不能維持14日齡哺乳仔豬體重增加2%。絕對量上只有4.63 g必需氨基酸(20%的膳食必需氨基酸進入門靜脈)和總計2.97 g的丙氨酸、天冬酰胺、羥脯氨酸和絲氨酸(11%的非必需氨基酸進入門靜脈)可用于腸外代謝(表1)。目前,即使給予新生仔豬充足的氨基酸合成底物,也不能確定它們能夠合成所有的非必需氨基酸(Wang等,2015)。我們對仔豬的研究結果表明,細胞外谷氨酰胺的濃度并不是17~21日齡仔豬腸上皮細胞合成瓜氨酸的主要限制因子(Wang等,2014)。相反,線粒體內低濃度的N-乙酰谷氨酸(變構激活線粒體內的氨甲酰磷酸合成酶I)限制了從鳥氨酸合成瓜氨酸(Sun等,2015)。此外,仔豬由補充的亮氨酸和甘氨酸分別合成絲氨酸與谷氨酰胺的速度較慢(Kim等,2004)。因此,新生兒氨基酸的合成被底物的可用性和酶活性所限制。
因此,給新生兒直接補充非必需氨基酸、它們合適的前體以及優質蛋白質(所有預成氨基酸的來源)可促進其生長。支持這一觀點的是,給出生體重正常的仔豬補充1%的谷氨酰胺、1%~2%的甘氨酸、0.2%~0.4%的精氨酸,1%~2%的亮氨酸(谷氨酸、天冬氨酸和精氨酸合成的前體),或牛奶蛋白(是從基礎日糧攝入蛋白的雙倍)后,促進了其瘦肉組織的生長和體重的增長(Lorenz等,2010;Davis和Fiorotto,2009)。大量的證據表明,缺乏合成的精氨酸、谷氨酰胺、甘氨酸限制了仔豬的最大生長。已有報道稱,缺乏合成精氨酸的大鼠和犢牛與缺乏合成谷氨酸和谷氨酰胺的大鼠有類似的結果(Sevasiadou等,2011)。另外,口服谷氨酰胺和甘氨酸可預防和治療新生兒的腸功能障礙,而口服谷氨酰胺可促進早產兒的腸道健康(Wu等,2011)。此外,日糧中添加甘氨酸可以改善乳蛋白攝取量未達標準的嬰幼兒的生長遲緩(Tan等,2009)。
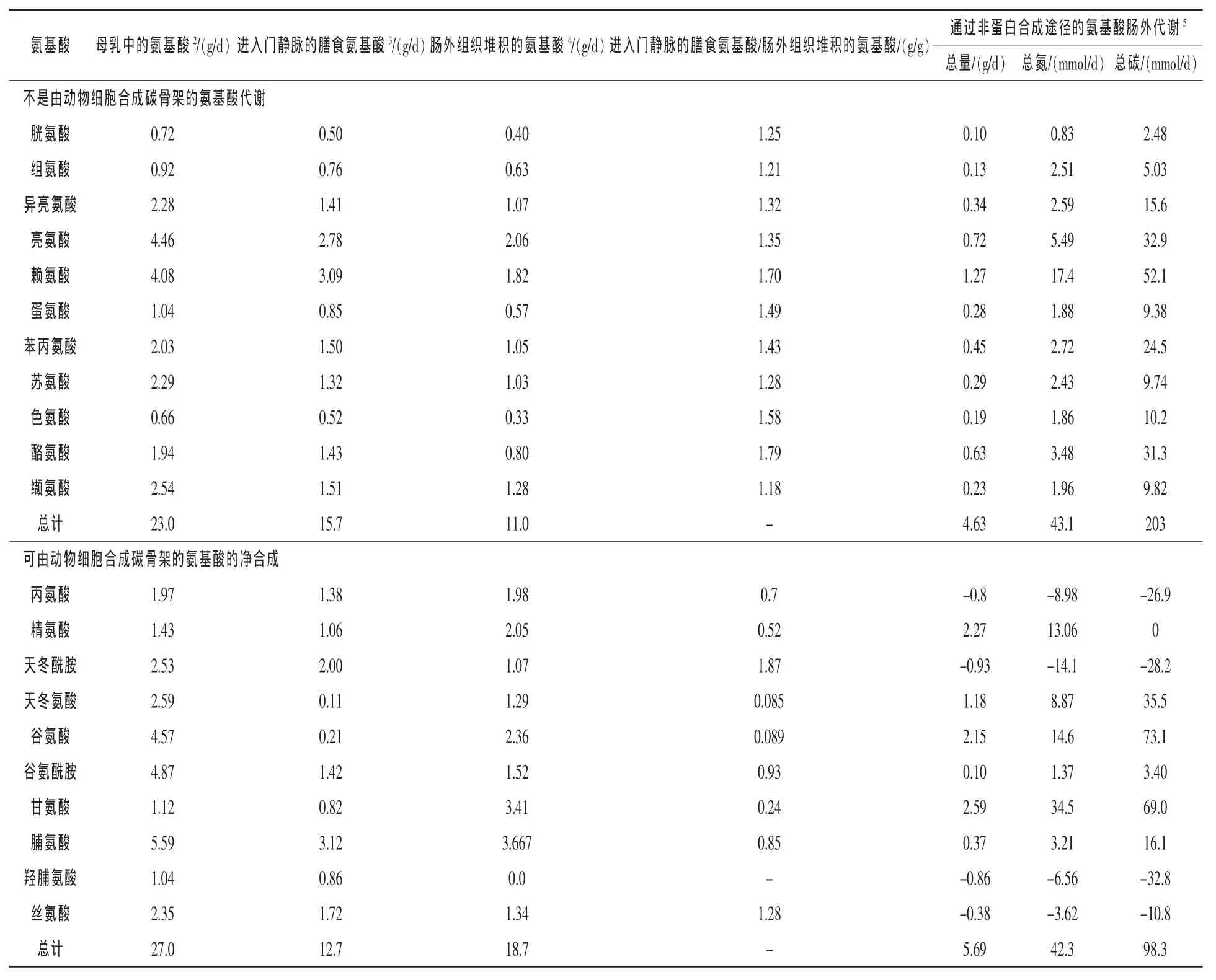
表1 14日齡哺乳仔豬(體重3.9 kg)的氨基酸代謝1
飼喂高蛋白配方(比從母乳攝入的蛋白質多50%)日糧會導致出生體重不足的仔豬生長減緩和33%的死亡率,可能因為它們不能通過肝尿素循環使由過量氨基酸衍生的氨無毒化(Tan等,2009)。因此,提高膳食蛋白質的攝入量不能用于促進早產兒的生長,營養學家應該考慮使用某些氨基酸在醫學和動物生產上實現這一目標。這很重要,因為出生體重不足的嬰兒和仔豬分別代表著5%~10%和15%~25%的新生兒總數。
[1]Baker D H.Advances in protein-amino acid nutrition of poultry[J].Amino Acids,2008,37(1):29~41.
[2]Brosnan J T,Silva R P D,Brosnan M E.The metabolic burden of creatine synthesis[J].Amino Acids,2011,40(5):1325~1331.
[3]Chen L,Peng L,Wang J,et al.Catabolism of nutritionally essential amino acids in developing porcine enterocytes[J].Amino Acids,2009,37(1):143~152.
[4]Chen L,Yin Y L,Jobgen W S,et al.In vitro oxidation of essential amino acids by jejunal mucosal cells of growing pigs☆[J].Livestock Science,2007,109(1~3):19~23.
[5]Curthoys N P,Watford M.Regulation of glutaminase activity and glutamine metabolism[J].Annual Review of Nutrition,1995,15(15):133~159.
[6]Dai Z L,Li X L,Xi P B,et al.l-Glutamine regulates amino acid utilization by intestinal bacteria[J].Amino Acids,2013,45(3):501~512.
[7]Dai Z L,Li X L,Xi P B,et al.Regulatory role for l-arginine in the utilization of amino acids by pig small-intestinal bacteria[J].Amino Acids,2012,43(1):233~244.
[8]Dai Z L,Wu G,Zhu W Y.Amino acid metabolism in intestinal bacteria:links between gut ecology and host health[J].Frontiers in Bioscience,2011,16(1):1768~1786.
[9]Davis T A,Fiorotto M L.Regulation of muscle growth in neonates[J]. Current Opinion in Clinical Nutrition&Metabolic Care,2009,12(1):78~85.
[10]Davis T A,Nguyen H V,Garcia-Bravo R,et al.Amino acid composition of human milk is not unique[J].Journal of Nutrition,1994,124(7):1126~1132.
[11]Dillon E L.Nutritionally essential amino acids and metabolic signaling in aging[J].Amino Acids,2012,45(3):431~441.
[12]Kim J,Song G,Wu G,et al.Functional roles of fructose[J].Proceedings of the National Academy of Sciences,2012,109(25):1619~1628.
[13]Kim S W,Mcpherson R L,Wu G.Dietary arginine supplementation enhances the growth of milk-fed young pigs[J].Journal of Nutrition,2004,134(3):625~630.
[14]Kim S W,Wu G.Regulatory role for amino acids in mammary gland growth and milk synthesis[J].Amino Acids,2009,37(1):89~95.
[15]Li P,Mai K,Trushenski J,et al.New developments in fish amino acid nutrition:towards functional and environmentally oriented aquafeeds.Amino Acids[J].Amino Acids,2008,37(1):43~53.
[16]Lorenz I,Fagan J,More S J.Calf health from birth to weaning.II.Management of diarrhoea in pre-weaned calves[J].Veterinary Ireland Journal,2010,64:9(1):975~980.
[17]Mavromichalis I,Parr T M,Gabert V M,et al.True ileal digestibility of amino acids in sow's milk for 17-day-old pigs[J].Journal of Animal Science,2001,79(3):707~713.
[18]Meléndez-Hevia E,Paz-Lugo P D,Cornish-Bowden A,et al.A weak link in metabolism:the metabolic capacity for glycine biosynthesis does not satisfy the need for collagen synthesis[J].Journal of Biosciences,2009,34(6):853~872.
[19]Reeds P J,Burrin D G,Jahoor F,et al.Enteral glutamate is almost completely metabolized in first pass by the gastrointestinal tract of infant pigs[J]. American Journal of Physiology,1996,270(270):413~418.
[20]Sevastiadou S,Malamitsi-Puchner A,Costalos C,et al.The impact of oral glutamine suppleme ntation on t he intestinal permeability and incidence of necrotizing enterocolitis/septicemia in premature neonates[J].The journal of maternal-fetal&neonatal medicine:the official journal of the European Association of Perinatal Medicine,the Federation of Asia and Oceania Perinatal Societies,the International Society of Perinatal Obstetricians,2011,24(10):1294~1300.
[21]Stoll B,Burrin D G.Measuring splanchnic amino acid metabolism in vivo using stable isotopic tracers[J].Journal of Animal Science,2006,84(13):60~72.
[22]Sun Y,Wu Z,Wei L,et al.Dietary l-leucine supplementation enhances intestinal development in suckling piglets[J].Amino Acids,2015,47(8):1~9.
[23]Suryawan A,Hawes J W,Harris R A,et al.A molecular model of human branched-chain amino acid metabolism[J].American Journal of Clinical Nutrition,1998,68(1):72~81.
[24]Tan B,Yin Y,Liu Z,et al.Dietary L-arginine supplementation increases muscle gain and reduces body fat mass in growing-finishing pigs[J].Amino Acids,2009,37(1):169~175.
[25]Trumbo P,Schlicker S,Yates A A,et al.Dietary reference intakes for energy,carbohydrate,fiber,fat,fatty acids,cholesterol,protein and amino acids[J]. Journal of the American Dietetic Association,2002,102(102):1621~1630.
[26]USA NRC.Nutrient Requirements Of Swine[M].National Academy of Sciences 1978.373~381.
[27]Wang H,Zhang C,Wu G,et al.Glutamine enhances tight junction protein expression and modulates corticotropin-releasing factor signaling in the jejunum of weanling piglets[J].Journal of Nutrition,2015,145(1):25~31.
[28]Wang W,Dai Z,Wu Z,et al.Glycine is a nutritionally essential amino acid for maximal growth of milk-fed young pigs[J].Amino Acids,2014,46(8):2037~2045.
[29]Wang W,Rezaei R,Wu Z,et al.Concentrations of free and peptide~bound hydroxyproline in the sow's milk and piglet plasma[J].Amino Acids,2013,45(3):595~595.
[30]Wang W,Wu Z,Dai Z,et al.Glycine metabolism in animals and humans:implications for nutrition and health[J].Amino Acids,2013,45(3):463~477.
[31]Wu G,Bazer F W,Burghardt R C,et al.Proline and hydroxyproline metabolism:implications for animal and human nutrition[J].Amino Acids,2011,40(4):1053~1063.
[32]Wu G,Bazer F W,Cross H R.Land-based production of animal protein:impacts,efficiency,and sustainability[J].Annals of the New York Academy of Sciences,2014b,1328(1328):18~28.
[33]Wu G,Bazer F W,Dai Z,et al.Amino acid nutrition in animals:protein synthesis and beyond[J].Annual Review of Animal Biosciences,2014a,2(1):387~417.
[34]Wu G,Bazer F W,Johnson G A,et al.Important roles for L-glutamine in swine nutrition and growth[J].Journal of Dairy Science,2010,93:3~3.
[35]Wu G,Jr M S.Arginine metabolism:nitric oxide and beyond[J].Biochemical Journal,1998,336(2):1~17.
[36]Wu G,Knabe D A,Kim S W.Arginine nutrition in neonatal pigs[J].Journal of Nutrition,2004,134(10).
[37]Wu G,Knabe D A.Free and protein-bound amino acids in sow’s colostrum and milk[J].Journal of Nutrition,1994,124(3):415~424.
[38]Wu G.Functional amino acids in growth,reproduction,and health[J].Advances in Nutrition,2010,1(1):31~37.
[39]Wu G.Intestinal mucosal amino acid catabolism[J].Journal of Nutrition,1998,128(8):1249~1252.
[40]Yeung A W,Terentis A C,King N J,et al.Role of indoleamine 2,3-dioxygenase in health and disease[J].Clinical Science,2015,129(7):601~672.
(未完待續)
S816.7
A
1004-3314(2016)09-0039-05
10.15906/j.cnki.cn11-2975/s.20160910
