酸棗葉表皮微形態對不同生態環境的適應特征
2016-10-25 08:02:56朱廣龍鄧榮華魏學智
生態學報 2016年16期
朱廣龍,鄧榮華,魏學智
山西師范大學生命科學學院, 臨汾 041000
?
酸棗葉表皮微形態對不同生態環境的適應特征
朱廣龍,鄧榮華,魏學智*
山西師范大學生命科學學院, 臨汾041000
以生長于煙臺—石家莊—寧夏—新疆不同生境形成的自然梯度干旱環境中的酸棗(Ziziphusjujubavar.spinosas)為材料,應用掃描電鏡和能譜分析研究酸棗葉表皮微形態對不同自然梯度干旱環境的適應特征。結果表明:酸棗葉片的上、下表皮都有表皮毛著生且在葉脈處分布濃密;葉片表層覆蓋有較厚的角質層和蠟質并形成突起,其主要成分為C、O、Ca三種元素;葉片表面氣孔數量平均40個/視野且按一定規律分布。不同生境中酸棗葉片表面都分布有大量的晶體,且晶體的形態和組成成分表現出一定的生態適應特征。晶體的形狀多樣,有正六棱體、長六棱體、不規則片狀、正方體、棒狀晶體或針狀,晶體的分布方式有單晶和簇晶。從煙臺到新疆隨干旱梯度的加劇,酸棗葉片表皮毛逐漸增多,氣孔數量減少,氣孔腔下陷明顯,有助于植株減少蒸騰保水抗旱;晶體的體積逐漸變小,數量增多,密度增大,提升了葉片硬度及抗旱性;酸棗葉片晶體的主要成分為C、O、Ca三種元素,且隨生境干旱加劇,C元素的含量逐漸增加,O和Ca元素含量依次減少,此外寧夏的晶體中還含有Si元素,進一步提升了葉片的機械性能。酸棗葉表皮微形態中表皮毛、角質層與蠟質、氣孔、晶體等在形態結構、生物功能和組成成分上特異性的變化是其長期生存于不同梯度干旱生境的生態適應特征。
葉表皮微形態;能譜分析;晶體;氣孔;生境
干旱是限制植物生長發育及生產的主要環境因子[1],植物長期生長于干旱的生境中,其生理特性和結構特征必然發生適應性的變化,并形成特異的抗旱特征。植物適應干旱脅迫的策略有兩種途徑:一是改變細胞代謝途徑,啟動各種防御機制[2- 3],如抗氧化保護酶活性的提高、滲透調節物質的積累[4];二是形態結構特征的改變,諸如根系和植株地上部分縮短節間、降低株高,葉片變小、著生腺毛、角質層加厚等[4- 9]。隨著全球氣候變化,區域性干旱頻發,干旱直接影響植物的生態環境和分布格局[10- 11]。植物對不同生態環境條件的適應機制存在差異,研究同種植物對不同生境的適應特征及機理對評估氣候變遷對植物的影響及指導生態建設恢復有重要的作用。
酸棗(ZizyphusjujubaMill.)為鼠李科(Rhamnaceae)棗屬(ZizyphusMill.)植物,原產于我國,廣布于華北、西北、東北和華東的向陽山坡、荒蕪丘陵和平原,從濕潤的海濱到干旱的荒漠均有分布[5]。酸棗根系深廣,抗旱、耐寒、耐貧瘠能力強,具有重要的水土保持作用,是綠化荒山的先鋒樹種[12]。目前對該植物的研究主要分布在生物活性物質提取[13- 14]、藥理作用探索[15]、組培嫁接[16]、抗旱生理及結構研究等方面[17- 19]。魏學智等[18]首次對酸棗葉片的結構進行了初步研究,發現其具有典型的旱生結構。賀少軒[19]、梁靜[20]和朱廣龍[5]等分別用野外覆膜、盆栽控水等干旱脅迫的方法研究了酸棗生理特性對干旱的響應,結果發現酸棗在生理特性上有較強的抗旱機能。康東東等[21]采用石蠟切片技術從形態結構上探究了不同地理環境中酸棗葉片的適應特征,但關于葉表皮微形態及其晶體的生態學意義,迄今尚不清楚。因此,本研究以不同生境條件下酸棗葉片為研究材料,利用掃描電鏡和能譜分析技術研究不同生境條件下酸棗葉表皮形態和晶體的適應特征,首次對酸棗葉表皮晶體的組成成分和生態功能進行分析闡釋,以期了解在全球氣候變遷的背景下植物對不同生態環境的適應機理。
1 材料與方法
1.1采樣地概況
實驗材料分別采自煙臺(萊山區大莊村)、石家莊(贊皇縣北溝村)、寧夏(銀川市賀蘭山附近)、新疆(吐魯番市紅星農場三隊),同時采集酸棗各個原生境土樣,測定土壤含水量。煙臺的萊山區大莊村(37°24′N,121°26′E)屬溫帶海洋性季風氣候,年降雨量651.9 mm,空氣濕潤系數為1.65,樣地土壤為壤土,含水量為14%左右。石家莊贊皇縣北溝村(37°42′N,114°18′E)屬暖溫帶半濕潤季風型大陸性氣候,年降雨量568 mm,空氣濕潤系數為1.07,樣地土壤為壤土,含水量為9%左右。寧夏銀川市賀蘭山(38°31′N,106°12′E)屬中溫帶大陸性氣候,年降雨量200 mm,空氣濕潤系數為0.50,樣地土壤為沙土,含水量為2%左右。新疆吐魯番市紅星農場三隊(42°55′N,89°11′E)屬大陸性暖溫帶干旱荒漠氣候,年降雨量15.6 mm,空氣濕潤系數為0.04,樣地土壤為壤土,含水量為4%。樣地從東到西(煙臺—石家莊—寧夏—新疆)構成一個自然的梯度干旱環境(表1)。

表1 采樣地區地理環境及氣候特征
濕潤系數是指一地降水量與蒸發量的對比關系,濕潤系數越小,則該地干燥程度越明顯; 各個樣地的濕潤系數根據, 當地氣象部門提供的氣象資料計算而得
1.2采樣方法
2010年7月,按自然氣候干旱梯度,分別從煙臺(萊山區大莊村)、石家莊(贊皇北溝村)、寧夏(銀川賀蘭山附近)、新疆(吐魯番紅星農場三隊)等地依次采樣。分別選取上述樣地樹齡5年(依據芽鱗痕判斷樹齡)的酸棗植株5株,選取植物頂部往下數第3個枝條、且都靠東部方向上枝條的成熟葉片10片,共選取50張葉片摘下葉片后立即分裝于(每個植株的葉片樣品單獨保存)FAA固定液(配方:95%酒精90 mL,福爾馬林5 mL,冰醋酸5 mL)中固定保存。
1.3電鏡掃描
選取固定液固定24 h以上、生長成熟、無病斑的酸棗葉片,乙醇(50%、70%、80%、90%、100%)逐級脫水,純乙醇洗滌2—3次, 轉入乙酸異戊酯中,CO2臨界點干燥儀(E- 3100,英國)干燥;JFC (JFC- 1600,日本) 型離子濺射及真空鍍膜儀中噴金鍍膜;JSM- 7500F(日本)掃描電鏡觀察拍照,并對葉表面晶體及蠟質成分用能譜儀(INCA mics 04BCC,日本)進行能譜分析[8]。
1.4統計分析
每個樣地隨機取 30 張切片拍照(從每個樣地每個植株的10個葉片中隨機取 6個葉片進行拍照,共計30張切片),FSX100 OLYMPS 觀察分析圖像, Motic Images Plus 2.0 圖像分析軟件測定,然后用SPSS13.0統計軟件對數據進行統計分析,SigmaPlot 10.0 軟件作圖, Photoshop 軟件圖像處理,排版。
2 結果
2.1酸棗葉片掃描電鏡
電鏡掃描顯示,酸棗葉片上、下表皮均有表皮毛著生,上表皮毛稀少且短(圖1 B),下表皮毛數量多,尤其是葉脈處表皮毛濃密較長(圖1A);上表皮細胞橢圓形或近圓球形,排列緊密(圖1C);下表皮氣孔數量較多,且按一定的規律分布:即保衛細胞較大的氣孔位于中央,周圍較小的氣孔圍繞其呈輻射狀圓環形分布(圖1D);葉片上表皮覆有厚厚的角質及蠟質,蠟質在表面形成突起,成塊狀分布(圖1E);葉片上、下表皮均分布有大量的晶體,尤其在下表皮氣孔周圍晶體數量較多(圖1E、F),晶體的形狀多樣,有正六棱體(圖1G)、長六棱體(圖1H)、不規則片狀(圖1I)、正方體(圖1J)、棒狀晶體或針狀(圖1J、K),晶體的分布方式有單晶(圖1K)和簇晶(圖1L),單晶是晶體單個分布,簇晶是晶體聚集成簇。在掃描電鏡下發現葉片下表皮氣孔腔中含有大量的晶體顆粒,有的晶體排列整齊,有的搖搖欲墜,即將排出(圖2)。

圖1 掃描電鏡下酸棗葉表皮共同特征Fig.1 The common characteristic of leaf epidermis of Ziziphus jujuba var. spinosas under scanning electron microscopeA:酸棗葉片背面主脈上著生的腺毛,×100;B:酸棗葉片正面主脈處的腺毛,×300;C:酸棗葉片正面的表皮細胞,×1000;D:酸棗葉片正面氣孔的分布,×500;E:酸棗葉片表面的蠟質,×10000;F:酸棗葉片表面的氣孔及晶體,×1500;G、H、I、J、K、L:酸棗葉片不同類型的晶體;G、H、×20000;L:×50000. I:K:×10000
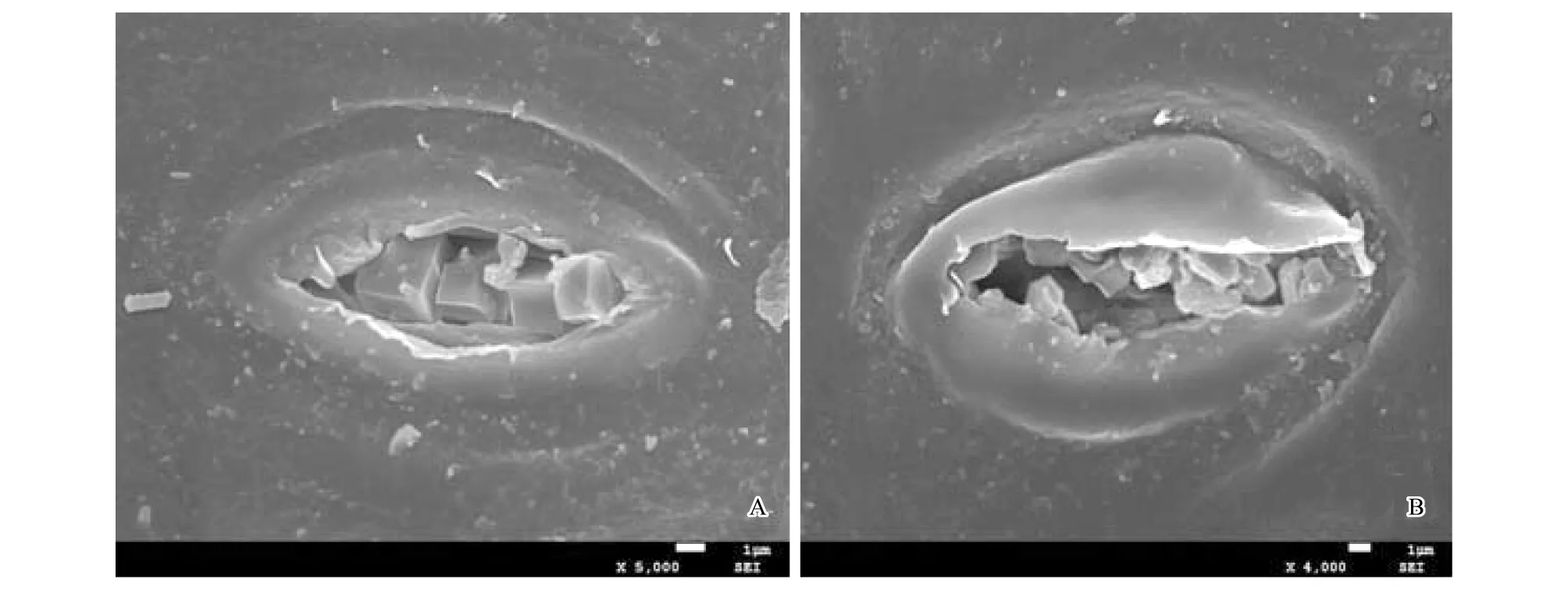
圖2 酸棗葉片氣孔排泄晶體,示葉表皮晶體的來源Fig.2 The source of crystals on the leaf epidermis of Ziziphus jujuba var. spinosas It showed the source of crystalsA:×5000;B:×4000
從煙臺、石家莊、寧夏到新疆,隨著自然干旱梯度的加劇,酸棗葉表皮毛逐漸增多(圖3Y1、S1、N1、X1),氣孔數量總體呈減少的趨勢,其中煙臺平均38.88個/視野(×300),石家莊和新疆的氣孔數量與煙臺相比分別減少了0.54%、8.26%,新疆的氣孔數量最少,平均為35.67個/視野,寧夏的氣孔數量最多,比煙臺增加了15.1%(圖3Y2、S2、N2、X2,圖4),然而4個樣地間酸棗葉表皮氣孔數差異不顯著(P>0.05)。氣孔長度以寧夏的最大,為 514.83 um,和寧夏的相比,石家莊和新疆酸棗葉表皮的氣孔長度分別減小了24.05%、26.18%(P<0.05);氣孔寬度總體呈變小的趨勢,煙臺的氣孔寬度最大,寧夏次之,新疆的最小,與煙臺相比,石家莊和新疆的氣孔寬度分別減小了47.13%和50.46%(P<0.05),而寧夏的減小了3.56%,差異不顯著(P>0.05)(圖3Y3、S3、N3、X3,圖5)。保衛細胞的厚度呈先增后減的趨勢,寧夏的保衛細胞厚度最大,平均為180.8μm,而新疆的最小,平均為 93.6μm,與煙臺相比,石家莊和寧夏分別增加了33.57%、46.05%,新疆的減小了24.39%(P<0.05)(圖3Y3、S3、N3、X3,圖6)。煙臺葉表皮晶體的形狀主要是多面體,無針晶或棒狀晶體,石家莊葉表皮中有針晶或棒狀晶體的分布,但數量較少,寧夏和新疆葉片中針晶或棒狀晶體分布較多,尤其新疆的葉片中,針晶聚集成簇(圖1L,圖3Y4、S4、N4、X4)。
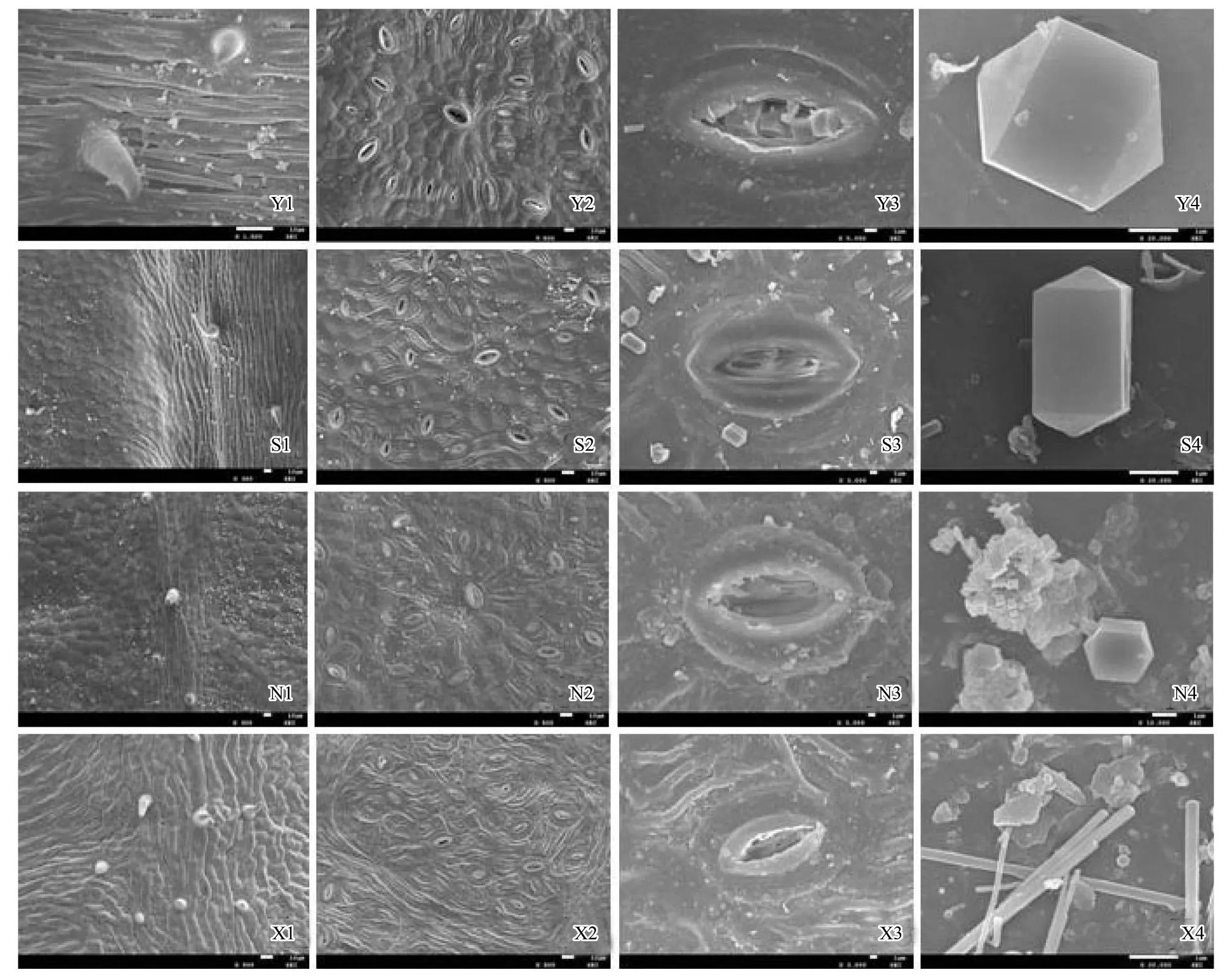
圖3 掃描電鏡下不同地區酸棗葉表皮微形態特征Fig.3 The characteristic of leaf epidermal micromorphology of Ziziphus jujuba var. spinosas in different ecotypes under scanning electron microscopeY:煙臺,S:石家莊,N:寧夏,X:新疆;Y1:煙臺酸棗葉片表面腺毛,×1500;S1:石家莊酸棗葉片氣孔分布,×300; N1:寧夏酸棗葉片氣孔分布,×300、X1:新疆酸棗葉片氣孔分布,×300;Y2:煙臺酸棗葉片氣孔分布,×500;S2:石家莊酸棗葉片氣孔分布,×500;N2:寧夏酸棗葉片氣孔分布,×500;X2:新疆酸棗葉片氣孔分布,×500;Y3:煙臺酸棗葉片氣孔腔,×3000;S3:石家莊酸棗葉片氣孔腔,×3000;N3:寧夏酸棗葉片氣孔腔,×3000;X3:新疆酸棗葉片氣孔腔,×3000;Y4:煙臺酸棗葉片晶體形態,×20000;S4:石家莊酸棗葉片晶體形態,×20000;N4:寧夏酸棗葉片晶體形態,×10000;X4:新疆酸棗葉片晶體形態,×20000
2.2酸棗葉片能譜分析
能譜分析結果表明:酸棗葉表皮蠟質的主要成分為C、O、Ca三種元素,其中C占62.09%,是蠟質的主要組成成分,O占19.69%,Ca占18.22%(圖7,表2)。煙臺、石家莊、新疆3個地域中,酸棗葉片晶體的成分都為C、O、Ca三種元素(圖8,圖9,圖10),寧夏的葉片晶體除含有C、O、Ca三種元素外,還含有少量的Si元素(圖11),據此推斷煙臺、石家莊、新疆的酸棗葉片晶體為碳酸鈣晶體或草酸鈣晶體,寧夏的還含有硅質體。
煙臺、石家莊、寧夏、新疆各地酸棗葉表皮晶體的成分能譜分析中,C元素的含量逐漸增多,新疆酸棗葉表皮晶體的C含量最高達到59.46%;O元素的含量逐漸減少,以煙臺的含量最高為42.91%,新疆的最少為24.03%;Ca元素的含量先增后減,石家莊的Ca含量最高為47.33%,新疆的Ca含量最少為16.51%(表3)。晶體形狀和組成成分的變化,是其長期適應梯度干旱的結果。

表 2 葉表皮蠟質能譜分析
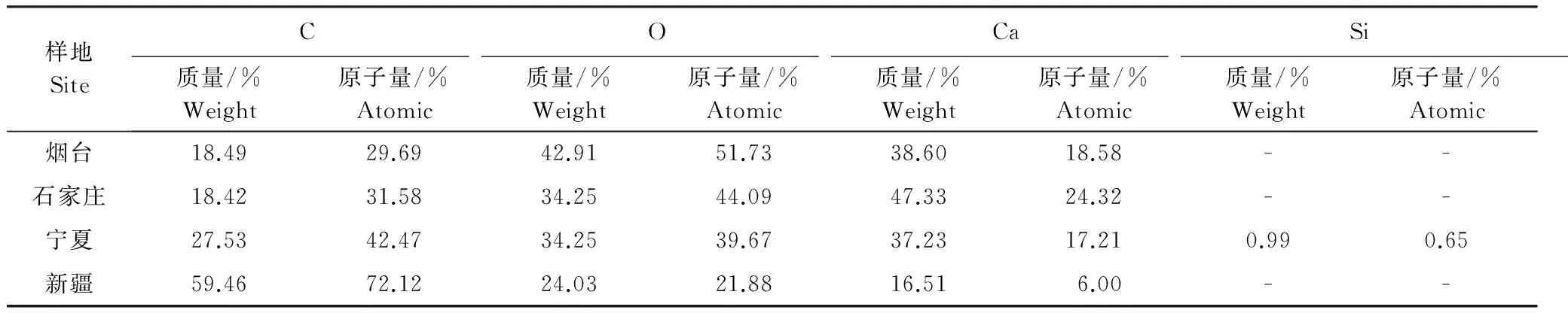
表3 不同地區酸棗葉表皮晶體能譜分析
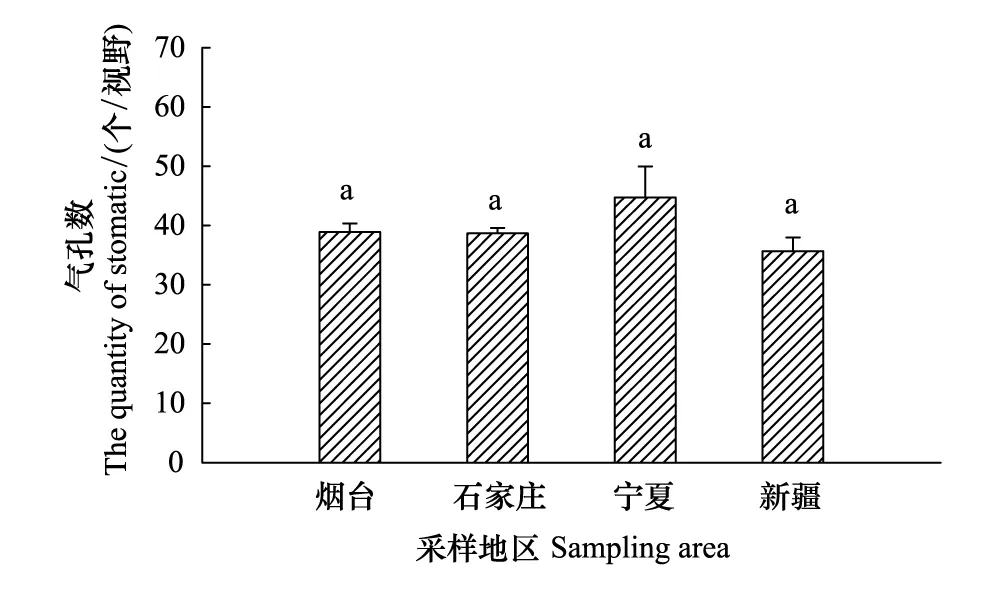
圖4 不同地區酸棗葉表皮氣孔數量變化Fig.4 The quantity of stomas of leaf epidermis of Ziziphus jujuba var. spinosas in different ecotypes
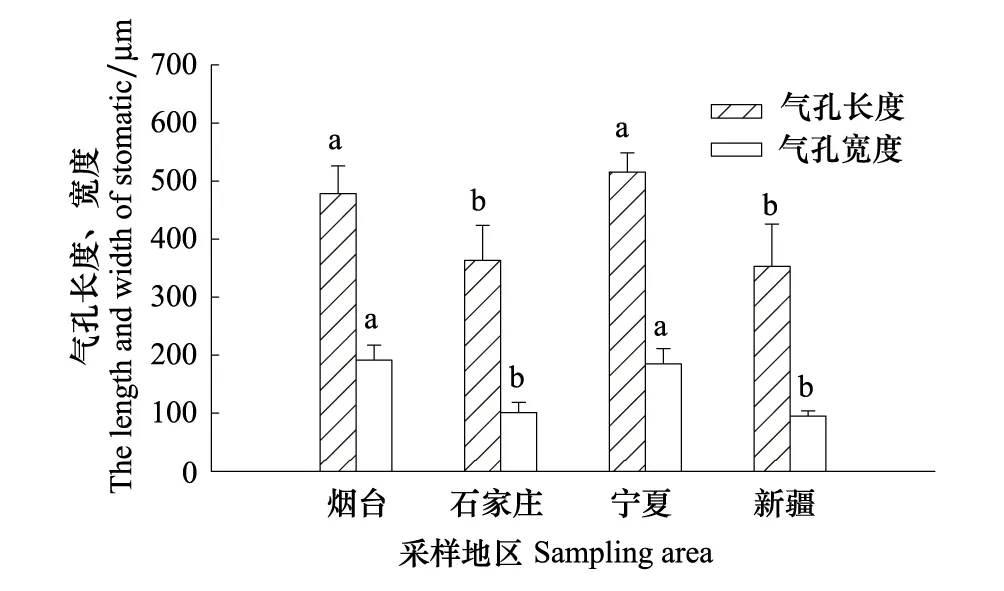
圖5 不同地區酸棗葉表皮氣孔長度、寬度變化Fig.5 The length and width of stomas of leaf pidermis of Ziziphus jujuba var. spinosas in different ecotypes圖中誤差異為SE,n=30圖中誤差異為SE,n=30

圖6 不同地區酸棗葉表皮氣孔保衛細胞厚度變化 Fig.6 The thickness of stomas guard cells of leaves of Ziziphus jujuba var. spinosas in different ecotypes圖中誤差異為SE,n=30
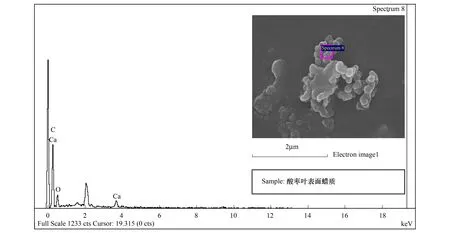
圖7 葉表皮蠟質能譜分析Fig.7 The energy spectrum analysis for waxiness of leaf epidermis

圖8 煙臺葉表皮晶體能譜分析Fig.8 The energy spectrum analysis for crystal of leaf epidermis in Yantai
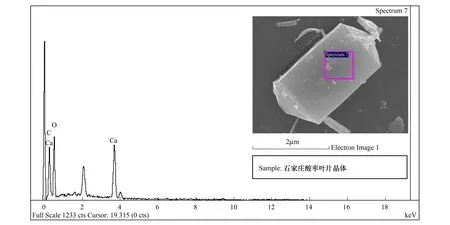
圖9 石家莊葉表皮晶體能譜分析Fig.9 The energy spectrum analysis for crystal of leaf epidermis in Shijiazhuang

圖10 新疆葉表皮晶體能譜分析Fig.10 The energy spectrum analysis for crystal of leaf epidermis in Xinjiang
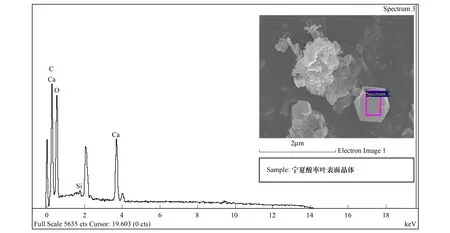
圖11 寧夏葉表皮晶體能譜分析Fig.11 The energy spectrum analysis for crystal of leaf epidermis in Ningxia
3 討論
水分是限制植物生長發育和作物生產的主要環境因子[23],干旱是植物生長過程中經常面臨的逆境之一,經過長期的演化,植物可對干旱環境采取多種適應對策[24]。從煙臺—石家莊—寧夏—新疆4個采樣地的土壤含水量、年降雨量及濕潤系數來看,其形成一個大尺度的自然梯度干旱環境的(表1),為酸棗的研究提供了一個得天獨厚的條件。酸棗在長期適應梯度干旱的過程中,演化出了一些高度耐旱的特性。從煙臺到新疆,四個樣地的酸棗葉表皮都有表皮毛分布,葉表皮覆蓋有較厚的角質層和蠟質并形成突起(圖3),且隨干旱程度的加劇,葉表皮表皮毛逐漸增多,角質層逐漸增厚(數據未發表,將在另一篇文章中呈現)。葉片是植物體暴露于環境中面積最大的器官,最易感知和適應環境變化而發生形態和結構上的改變,葉表皮表皮毛和厚的角質層可以有效防止水分蒸騰,是反映植物抗旱性的一個重要指標[23- 24]。趙小仙等[25]研究表明,隨干旱程度的加劇,細胞角質化外壁增厚,氣室變大,能有效減少水分蒸發,增加沙拐棗的抗旱性。這與本文對酸棗葉表皮的研究結果相似,角質層和蠟質加厚形成突起都是植物對不同干旱生境長期適應的結構特征。此外,堅硬的角質層除具有保水作用外,還具有機械支撐作用,使植株在水分缺乏時不會立即萎蔫[26]。酸棗葉表皮表皮毛、角質層與蠟質的變化,是其長期對不同生境的適應特征之一。
氣孔是蒸騰作用的主要通道,同時也是光合作用和呼吸作用交換氣體的開關,此外氣孔還具有排泄功能。本研究發現,從煙臺到新疆隨著生境干旱程度的加劇,酸棗葉表皮氣孔的紋飾變密,氣孔密度增加(研究發現隨干旱梯度增加,4個樣地中酸棗葉片葉面積顯著減小,而氣孔數量減少的差異不顯著,因此氣孔密度增加),氣孔腔由外凸逐漸下陷,氣孔開度變小,保衛細胞厚度增加。
一般認為旱生植物的氣孔密度增加是一種適應特征、氣孔密度增加是植物對旱生環境的適應,氣孔密度低是植物對濕生環境的適應,在環境干旱程度增加時植物提高氣孔分化的程度,增加氣孔密度,待水分供應充足時,可增加氣體交換,以提高光合作用效率[27]。氣孔腔下陷、凹陷的氣孔可能有利于滯留CO2和H2O,在旱生條件下可為光合作用提供充足的原料,同時還可以使內在空氣的飽和差值減小,阻止水分散失,形成較濕潤的小環境,這可能是植物長期在逆境中產生的抗御蒸騰的機制[28];此外下陷的氣孔在減少光線輻射和風沙損害方面有積極的作用[29]。氣孔長寬比減小可能是為在長徑一定的情況下增加氣孔開口度,加大呼吸面積,從而補償呼吸效率,增加通氣量,使酸棗的氣孔結構具有較強的抗逆特性[30],這與康東東等[21]對不同地理環境下酸棗葉的形態解剖特征研究結論一致。酸棗葉表皮氣孔形態特征的變化,是其長期適應于不同生境的又一重要策略。
晶體是植物產生的一種具有特殊形態結構與生理功能的代謝物,晶體多集中在沙生、旱生植物的葉表皮、內部和軸器官中,被認為是荒漠植物抗旱結構特征之一, 晶體的形成可能與荒漠植物適應干旱和鹽堿環境有關[22,31-32]。本研究發現酸棗的葉表皮分布有較多的晶體,且從煙臺到新疆隨生境干旱程度加劇,酸棗葉表皮晶體的形狀由多面體向針晶過渡(圖3),干旱越嚴重的生境晶體的體積越小,數量和密度越大。晶體可增加葉片的硬度,防止風沙損傷[33],因此寧夏和新疆生境中酸棗葉表皮晶體數量和形態特征的變化是長期適應其多風沙環境的結果。有研究表明,晶體具有較高的滲透勢, 吸水能力強, 在外界環境條件適宜、植物水分輸導良好時, 晶體可以吸收并貯存水分呈溶解狀態;當外界環境干旱、導管中水分輸導受阻而不能正常滿足植物需水時,晶體呈析出狀態而釋放水分,為其周邊細胞提供一個較為濕潤的小環境,從而提高植物的抗旱性[22,33- 34]。此外還有研究表明晶體的存在一方面可加強葉片的機械性能,另一方面可避免重金屬離子對細胞的毒害作用、改變細胞的滲透勢、提高吸水和保水能力[33- 34]。
能譜分析表明,晶體的組成元素主要為C、O、Ca三種元素,說明該晶體主要為碳酸鈣晶體或/和草酸鈣晶體,且各元素的含量及比例隨干旱梯度環境呈規律性的分布,C含量逐漸增加,O和Ca的含量依次減少(表3)。從化學結構上分析推測可知,C含量的增加可增大碳鏈的長度和不飽和程度,有利于增加晶體對水的吸水和束水能力,在寧夏和新疆極度干旱的野外生境中,酸棗葉表皮晶體在雨季可以涵養更多的水分,在旱季或逆境時將析出釋放水分貢獻于植株維持生命活動,從而提高植株的耐旱性。Ca含量隨生境干旱加劇而依次減少的變化特征可能與各地生境的氣候特征和各生境中植株各自的代謝水平有關,我國中、東南部地區工業發達環境污染較重,NO2、SO2等大量排放造成酸雨危害趨勢加重[35- 38]。受酸雨等環境變化的影響,植被代謝過程中產生較多的草酸等酸性代謝物[39],此類物質對植物體會造成傷害,植株通過吸收或從晶體中析出更多的鈣離子與草酸等代謝物相結合,形成中性的螯合物減輕對植物的傷害[40];而生存于寧夏和新疆等西北地區植株體內酸性代謝物少,所以酸棗植株對鈣離子等堿性離子的吸收較少,則儲藏于晶體中的鈣離子含量就少[40]。此外,寧夏的酸棗葉表皮的晶體還有少量的Si元素,反映出寧夏酸棗葉表皮的晶體還有硅質體,硅質體的主要成份是SiO2,硅質體多為圓簇狀小團塊, 除氫氟酸外不溶于其它任何酸,其多存在于表皮細胞或薄壁組織中, 少數分布在纖維束周圍的細胞中而類似晶鞘纖維,可增強葉片的機械性能、減少水分散失[22]。由此可見,酸棗葉表皮晶體的形態特征、類型及組成元素含量的變化是其長期適應于不同生境的又一重要適應機制。
[1]Xu S C, Ding H D, Su F X, Zhang A Y, Jiang M Y. Involvement of protein phosphorylation in water stress-induced antioxidant defense in maize leaves. Journal of Integrative Plant Biology, 2009, 51(7): 654-662.
[2]Zhu J K. Salt and drought stress signal transduction in plants. Annual Review of Plant Biology, 2002, 53: 247- 273.
[3]Boudsocq M, Lauriere C. Osmotic signaling in plants: Multiple pathways mediated by emerging kinase families. Plant Physiology, 2005, 138(3): 1185- 1194.
[4]汪攀, 陳奶蓮, 鄒顯花, 馬祥慶, 吳鵬飛. 植物根系解剖結構對逆境脅迫響應的研究進展. 生態學雜志, 2015, 34(2): 550- 556.
[5]朱廣龍. 酸棗生理生化特性及結構特征對梯度干旱的響應[D]. 臨汾: 山西師范大學, 2012.
[6]Brodribb T J, Holbrook N M. Water stress deforms tracheids peripheral to the leaf vein of a tropical conifer. Plant Physiology, 2005, 137(3): 1139- 1146.
[7]Atkin O K, Macherel D. The crucial role of plant mitochondria in orchestrating drought tolerance. Annals of Botany, 2009, 103(4): 581- 597.
[8]朱廣龍, 趙挺, 康冬冬, 魏學智. 木香薷腺毛形態結構發生發育規律的研究. 植物研究, 2011, 31(5): 518- 523.
[9]王順才, 鄒養軍, 馬鋒. 干旱脅迫對3種蘋果屬植物葉片解剖結構、微形態特征及葉綠體超微結構的影響. 干旱地區農業研究, 2014, 32(3): 15- 23.
[10]黎祜琛, 邱治軍. 樹木抗旱性及抗旱造林技術研究綜述. 世界林業研究, 2003, 16(4): 17- 22.
[11]趙成章, 高福元, 石福習, 任珩, 盛亞萍. 高寒退化草地甘肅臭草種群分布格局及其對土壤水分的響應. 生態學報, 2011, 32(22): 6688- 6695.
[12]王慧莉, 田濤, 王建永, Batool A, 趙旭喆, 莫非, Akram N A, 熊友才. 旱區農業雨水資源利用與生態系統可持續性: 2013干旱農業和生態系統可持續性國際會議綜述. 生態學雜志, 2014, 33(11): 3127- 3136.
[13]王旭峰, 何計國, 陳陽, 李欣, 李智欣. 酸棗仁皂苷的提取及改善睡眠功效的研究. 食品科學, 2006, 27(4): 226- 229.
[14]丁軻, 張彤楠, 韓濤. 快速溶劑萃取聯合高效液相色譜法測定不同產地酸棗仁中皂苷A、B的含量. 食品工業科技, 2014, 35(23): 265- 270.
[15]Barrieu F, Chaumont F, Chrispeels M J. High expression of the tonoplast aquaporinZmT1P1 in epidermal and conducting tissues of maize. Plant Physiology, 1998, 117(4): 1153- 1163.
[16]Yu S W. Plant Physiology and Molecular Biology. Beijing: Science Press, 1992.
[17]曹正其. 加強技術機制創新 提高退耕還林科技含量. 四川林業科技, 2001, 22(2): 1- 3.
[18]魏學智, 畢潤成. 旱生植物酸棗的葉結構. 山西師大學報: 自然科學版, 1997, 11(3): 44- 47.
[19]賀少軒, 梁宗鎖, 蔚麗珍, 周自云. 土壤干旱對2個種源野生酸棗幼苗生長和生理特性的影響. 西北植物學報, 2009, 29(7): 1387- 1393.
[20]梁靜, 魏學智. 酸棗葉對土壤水分的生理生化響應. 植物研究, 2010, 30(5): 549- 555.
[21]康東東, 韓利慧, 馬鵬飛, 魏學智, 畢潤成. 不同地理環境下酸棗葉的形態解剖特征. 林業科學, 2008, 44(12): 135- 140.
[22]朱廣龍, 馬茵, 韓蕾, 霍張麗, 魏學智. 植物晶體的形態結構、生物功能及形成機制研究進展. 生態學報, 2014, 34(22): 6429- 6439.
[23]Jaleel C A, Manivannan P, Wahid A, Farooq M, Al-Juburi H J, Somasundaram R, Panneerselvam R. Drought stress in plants: a review on morphological characteristics and pigments composition. International Journal of Agriculture and Biology, 2009, 11(1): 100- 105.
[24]王勛陵, 馬驥. 從旱生植物葉結構探討其生態適應的多樣性. 生態學報, 1999, 19(6): 787- 792.
[25]趙小仙, 李毅, 蘇世平, 白瀟. 3個地理種群蒙古沙拐棗同化枝解剖結構及抗旱性比較. 中國沙漠, 2014, 34(5): 1293- 1300.
[26]林植芳, 林桂珠. 鼎湖山植物葉片的一些與光合作用有關的結構特征 // 中國科學院華南植物研究所集刊. 北京: 科學出版社, 1989: 101- 107.
[27]Cutter E G. 植物解剖學. 李正理譯. 北京: 科學出版社, 1973.
[28]胡適宜. 植物學(形態學及解剖學部分). 北京: 高等教育出版社, 1959.
[29]張道遠, 張娟, 譚敦炎, 潘伯榮. 國產檉柳科3屬6種植物營養枝的解剖觀察. 西北植物學報, 2003, 23(3): 382- 388.
[30]李正理, 李榮敖. 我國甘肅九種旱生植物同化枝的解剖觀察. 植物學報, 1981, 23(3): 181- 185.
[31]劉家瓊. 我國荒漠不同生態類型植物的旱生結構. 植物生態學與地植物學叢刊, 1982, 6(4):314- 319.
[32]蘇培璽, 安黎哲, 馬瑞君, 劉新民. 荒漠植物梭梭和沙拐棗的花環結構及C4光合特征. 植物生態 學報, 2005, 29(1): 1- 7.
[33]李芳蘭, 包維楷. 植物葉片解剖形態結構對環境變化的響應與適應. 植物學通報, 2005, 22(增刊): 118- 127.
[34]韋存虛, 張軍, 王建軍, 孫國榮. 星星草營養器官適應鹽脅迫的結構特征. 植物資源與環境學報, 2006, 15(1): 51- 56.
[35]邵學新, 黃標, 顧志權, 錢衛飛, 金洋, 畢葵森, 嚴連香. 長三角經濟高速發展地區土壤pH時空變化及其影響因素. 礦物巖石地球化學通報, 2006, 25(2): 143- 149.
[36]郝吉明, 段雷, 謝紹東. 中國土壤對酸沉降的相對敏感性區劃. 環境科學, 1999, 20(4): 1- 5.
[37]賈曼. 石家莊污灌區土壤重金屬污染現狀調查及其風險評價[D]. 石家莊: 河北醫科大學, 2012.
[38]趙艷霞, 侯青. 1993- 2006 年中國區域酸雨變化特征及成因分析. 氣象學報, 2008, 66(6): 1032- 1042.
[39]黃曉華, 陸天虹, 周青, 曾慶玲, 梁嬋娟. 酸雨傷害植物機理與稀土調控研究. 中國生態農業學報, 2004, 12(3): 116- 118.
[40]徐靜靜, 慈華聰, 何興東, 薛蘋蘋, 趙雪萊, 郭健潭, 高玉葆. 天津鹽漬化生境54種植物鈣晶體與鈣組分特征. 應用生態學報, 2012, 23(5): 1247- 1253.
Leaf epidermal micromorphology ofZiziphusjujubavar.spinosain response to a gradient of drought stress
ZHU Guanglong, DENG Ronghua, WEI Xuezhi*
CollegeofLifeSciences,ShanxiNormalUniversity,Linfen041000,China
Global climate change is predicted to increase average temperatures, alter geographical patterns of rainfall, and increase the frequency of extreme climatic events. Such changes are likely to alter the timing and magnitude of drought stresses experienced by plants. Drought and heat events are occurring more frequently with climate change; therefore, plants must develop morphological and physiological characteristics that confer tolerance to different drought conditions (ecotypes). The objective of this study was to explore leaf epidermal micromorphology ofZiziphusjujubavar.spinosain response to different natural drought conditions. The plant samples were collected from Yantai, Shijiazhuang, Ningxia, and Xinjiang provinces, China, which form a gradient of environmental drought according to soil moisture, annual precipitation, and humidity. We found that hairs developed in both the upper and lower epidermis, which was thicker in leaf veins and functioned to reduce evaporation from the leaf surface. Cuticles and wax composed of carbon (C), oxygen (O), and calcium (Ca) were distributed in the leaf epidermis and formed protuberances. Large numbers of stomata (approximately 40 actual unit area were arranged in regular patterns on the leaf epidermis. Numerous crystals were also observed on the epidermis in each ecotype, and the shape and composition of the crystals differed among the ecotypes. The crystals were categorized according to shape and included six-surfaced, irregularly schistose, cubic, rhabdolith, and raphide forms. Single crystals or crystal clusters were the common crystal types. As the natural drought gradient increased from Yantai to Xinjiang, the density of leaf epidermal hairs inZ.jujubavar.spinosaincreased, the number of stomata decreased, and stoma were more shrunken, which could help plants to reduce transpiration and tolerate drought. The numbers of crystals increased and crystal volume diminished, which would enhance leaf rigidity and drought resistance. Crystals mainly included C, O, and Ca; however, silica (Si) was detected in the Ningxia ecotype, in which it enhanced leaf mechanical properties. With increasing drought severity, carbon content increased but oxygen and calcium decreased. In summary, changes in morphological structure, biological function, and leaf epidermal micromorphology (including epidermal hairs, cuticles, wax, and crystals) represented adaptive ecological characteristics for long-term drought tolerance.
leaf epidermal micromorphology; energy spectrum analysis; crystal; stoma; ecotype
國家自然科學基金資助項目(30972396);山西省自然科學基金項目資助(2009011041- 1)
2015- 01- 24; 網絡出版日期:2015- 11- 30
Corresponding author.E-mail: wxz3288@163.com
10.5846/stxb201501240191
朱廣龍,鄧榮華,魏學智.酸棗葉表皮微形態對不同生態環境的適應特征.生態學報,2016,36(16):5193- 5203.
Zhu G L, Deng R H, Wei X Z.Leaf epidermal micromorphology ofZiziphusjujubavar.spinosain response to a gradient of drought stress.Acta Ecologica Sinica,2016,36(16):5193- 5203.
猜你喜歡
國畫家(2022年2期)2022-04-13 09:07:46
四川文學(2021年4期)2021-07-22 07:11:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
絲綢之路(2014年9期)2015-01-22 04:24:46
兒童與健康(2011年4期)2011-04-12 00:00:00
