干旱脅迫對頂果木生理特性及根系形態特征的影響
2016-10-19 13:11:57李付伸覃德文杜佩連秦武明鐘連香
湖北農業科學 2016年8期
關鍵詞:植物
李付伸 覃德文 杜佩連 秦武明 鐘連香
摘要:通過盆栽試驗,用Win Rhizo根系分析軟件,分析不同模擬干旱脅迫條件對頂果木(Acrocarpus fraxinifoliusa Wight ex Arn.)根系生長和生理動態的影響。結果表明,干旱脅迫降低了頂果木葉片 CAT 活性及 MDA 含量,升高了SOD、POD活性,這些生理變化能有效降低植物細胞膜脂過氧化的程度,提高了頂果木抗干旱脅迫的能力。并且根半徑在逐漸降低,根系皮層和維管束厚度隨干旱脅迫程度的加重總體趨勢在變薄,這有利于水分與營養物質的輸送。說明頂果木具備一定的抗旱能力,可在廣西大力推廣種植。
關鍵詞:頂果木(Acrocarpus fraxinifoliusa Wight ex Arn.);干旱;根系;生理特性
中圖分類號:ST92.99 文獻標識碼:A 文章編號:0439-8114(2016)08-2019-04
DOI:10.14088/j.cnki.issn0439-8114.2016.08.026
Abstract: Through pot experiment and Win Rhizo root system analysis software, the analysis of different simulated drought stress conditions on top of Acrocarpus fraxinifoliusa Wight ex Arn. root system growth and physiological dynamic effects. The results showed that drought stress reduced the A. fraxinifoliusa leaf CAT activity and MDA content and increased SOD, POD activity, these physiological changes can effectively reduce the degree of plant cell membrane lipid peroxidation and improve A. fraxinifoliusa drought resistance ability. And root radius is gradually reduced, the root cortex and vascular cylinder thickness with the drought stress degree aggravating the overall trend in the thinning, which is conducive to the transport of water and nutrients. That A.fraxinifoliusa has the capacity of drought resistance, and can be planted in Guangxi to promote.
Key words: Acrocarpus fraxinifoliusa Wight ex Arn.; drought stress; root system; physiology characteristics
頂果木(Acrocarpus fraxinifoliusa Wight ex Arn.)為蘇木科(Caesalpiniaceae)頂果樹屬(Acrocarpus Wight ex Arn.)高大無刺喬木,樹高平均達到40 m,枝下高均為20 m,胸徑最大可達120 cm以上,其分布于廣西壯族自治區的西部、西南部,貴州、云南省也有大量分布,為國家三級重點保護植物[1]。頂果木適應能力較強,土壤適應范圍在pH 4.3~7.9。在廣西的龍州縣、田林縣、田陽縣等石灰巖山地或土山區域分布較多;其中,22年生的頂果木樹干通直,單株材積可達1.189 5 m3[2]。該樹作為優質木材出材率高、材質堅硬、耐腐、紋理美觀,為優質家具和纖維用材;其樹形美觀呈傘狀,常應用于行道樹、風景樹或生態防護林建設。頂果木根系較為發達,樹冠冠幅寬大,林分生長速度快,更新速率高,可應用于涵養水源、保持水土和改良土壤等生態建設工程,尤其是用于石山巖溶地區石漠化治理和荒山綠化效果顯著[3,4]。由于頂果木眾多生物學特性未被挖掘,國內外學者普遍關注頂果木的研究,近年來主要對頂果木的生物學特性[5]、蟲害[6]、育苗造林[7]、無性繁殖[8]、立地適應性[9]等方面展開了研究。然而,隨著全球氣候變化劇烈,廣西大部分地區普遍出現了久旱等極端氣候現象,長時間缺少降雨,嚴重影響了眾多物種的正常生長,也給林業工作者提出了挑戰,植物如何應對干旱極端生境已成為了當今生態學家普遍關注的熱點。試驗主要是對頂果木進行干旱脅迫處理,觀察頂果木如何調節生理生化特性與根系的生長來應對缺乏水分的逆境脅迫,從而為干旱地區的森林培育和營林規劃提供科學依據。
1 材料與方法
1.1 材料
參試材料為廣西國有維都林場優質一年生頂果木實生幼苗,在2014年3月1日移栽到廣西大學林學院苗圃試驗大棚內,采用盆栽方法對苗木進行干旱脅迫試驗。用黃土∶細沙為3∶1的混合土作為育苗基質,基質土在裝盆前用0.5%的高錳酸鉀溶液消毒。
1.2 處理
參考有關研究結果[10,11],結合當地干旱時測定的土壤含水量,試驗設置4個水分梯度(模擬干旱脅迫)處理,分別為①CK,正常供水(土壤水分含量為田間最大持水量的80%,簡寫為80%,下同);②P1,輕度干旱脅迫(65%);③P2,中度干旱脅迫(35%);④P3,重度干旱脅迫(15%)。2014年4月30日,將溫室中培育的生長狀況良好的具有12~20片葉、地下根系長度5 cm、株高為30~35 cm的頂果木幼苗移植入苗木盆內,每個處理6盆,每盆1株苗木,在此期間進行日常的養護管理。待移植苗木適應了苗木盆生長環境后,在2014年9月10日至10月1日,用稱重法控制土壤含水量,各處理于每日17:00時用電子天平稱重補水,保證土壤內的含水量控制在試驗設定的范圍內。
1.3 測定方法
1.3.1 生理指標測定 干旱處理至2014年10月20日,不同處理植株葉片各取0.5 g,加 0.05 mo1/L磷酸緩沖液(pH 7.8,內含 1% PVP)5.0 mL及少量石英砂,在碎冰上研磨,樣品研碎后置于15 000 r/min、4 ℃下離心15 min,選取上清液置于10 mL容量瓶內,定容,即為酶提取液,對酶提取液分別進行丙二醛(MDA)含量、質膜相對透性、過氧化物酶(POD)、過氧化氫酶(CAT)及超氧化物歧化酶(SOD)活性測定,都重復6次。MDA含量采用硫代巴比妥酸法測定[12];質膜相對透性采用數顯電導率儀測定,以相對電導率表示[12],
相對電導率=(浸泡電導率-空白電導率)/(煮沸后電導率-空白電導率)×100%。
SOD活性采用氮藍四唑(NBT)光氧化還原法測定,酶活單位用U/mg(FW)表示[13]。CAT 活性采用高錳酸鉀還原法測定,酶活單位用μmol(H2O2)/[g(FW)·min]表示[12]。POD活性采用愈創木酚法測定,酶活單位用以△OD470/[g(FW)·min]表示[14]。
1.3.2 根系測定 在2014年10月20日試驗處理完成后,采用Win RHIZON Pro STD160+型根系圖像分析系統(Regent Instruments Ine,Quebec,Canada)對不同處理的苗木根系總根長、平均根系直徑和根尖數等參數進行測定。并結合光學顯微鏡(MOTIC BA300,USA)對頂果木根系皮層厚度變化、維管束厚度變化、細胞破碎化程度進行形態觀察,結合顯微測微尺測量,進行統計計算。每個測定指標取顯微鏡觀察下20個視野的平均值。
1.4 數據處理
試驗結果以測定的平均值表示。數據的統計分析采用Microsoft Office Excel 2003軟件進行,利用鄧肯氏新復極差檢驗法(Duncan′s new multiple range test,DMRT)進行差異顯著性檢驗。
2 結果與分析
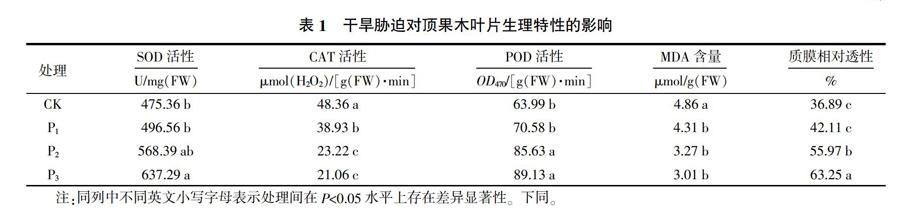
2.1 干旱脅迫對頂果木葉片生理特性的影響
干旱脅迫對頂果木葉片生理的影響情況見表1。由表1可知,不同干旱脅迫處理影響了頂果木葉片的SOD、POD、CAT活性和質膜相對透性及MDA含量。與正常供水條件相比,輕度干旱、中度干旱脅迫處理提高了頂果木葉片的SOD活性,但差異不顯著(P>0.05),而重度干旱脅迫處理大幅度提高了SOD活性,并且差異顯著(P<0.05)。在CAT活性方面,3個干旱脅迫處理都大幅度降低了頂果木葉片的CAT活性,并與對照差異顯著(P<0.05)。而在POD活性方面,3個干旱脅迫處理都提高了頂果木葉片的POD活性,其中輕度干旱脅迫處理提高的幅度不大,差異不顯著(P>0.05);而中度干旱、重度干旱脅迫處理增加幅度很大,與對照差異顯著(P<0.05)。在MDA含量方面,隨干旱程度的加重MDA含量降低,3個干旱脅迫處理都與對照差異顯著(P<0.05)。在質膜相對透性方面,隨干旱程度的加重質膜相對透性都在提升,其中輕度干旱脅迫處理與對照差異不顯著(P>0.05);而中度干旱、重度干旱脅迫處理與對照差異顯著(P<0.05)。
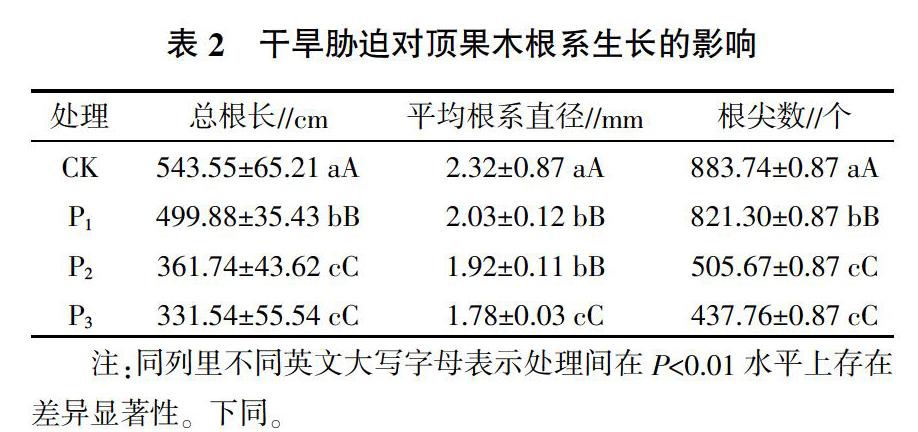
2.2 干旱脅迫對頂果木幼苗根系生長的影響
干旱脅迫對頂果木幼苗根系生長的影響情況見表2。由表2可見,隨著干旱強度的增加,頂果木幼苗的平均根系直徑、總根長和根尖總數都呈降低趨勢,3個干旱脅迫處理的平均根系直徑、總根長和根尖總數都與對照的平均根系直徑、總根長和根尖總數差異極顯著(P<0.01)。
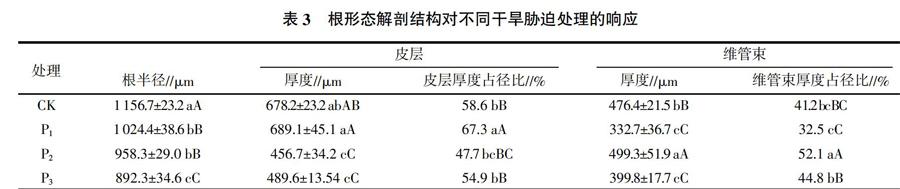
2.3 干旱脅迫下頂果木根形態解剖結構的變化
干旱脅迫對頂果木根形態解剖結構的影響情況見表3。由表3可見,隨著干旱脅迫程度的加重,頂果木的根半徑與對照相比在逐漸降低,并且皮層厚度和維管束厚度也呈降低的趨勢,尤其是重度干旱脅迫處理與對照相比,在根半徑、皮層厚度和維管束厚度上都與對照差異顯著(P<0.05)。然而從另一個角度來看,皮層厚度和維管束厚度的占徑比例呈增加的趨勢,同樣是重度干旱脅迫處理與對照相比,在皮層厚度占徑比和維管束厚度占徑比上都與對照差異不顯著(P>0.05),說明頂果木對干旱脅迫具有很強的自我調節能力。
3 討論
水分是影響植物生長發育的重要因子,在干旱生境下植物的細胞膜活性氧自由基活性加強,直接對細胞造成損害,此時植物細胞內的SOD、POD和CAT活性強弱成為植物抵御干旱脅迫的主要方式[15]。有研究表明,向日葵、高粱和煙草幼苗在缺水條件下CAT活性提高,植物出現萎蔫[16,17];但也有研究表明,隨水分脅迫的加重木麻黃小枝[18]和花生葉片[19]的SOD、CAT和POD活性下降,并使花生葉片MDA含量降低。頂果木試驗結果表明,干旱脅迫降低了頂果木葉片CAT活性及MDA含量,升高了SOD、POD活性,這與周建等[20]、李妮亞等[21]的研究結果相同。干旱脅迫下頂果木葉片的這些生理特性變化能夠有效降低植物細胞膜脂過氧化的程度,提高了頂果木抗干旱脅迫的能力,這與在棉花[22]、咖啡[23]和玉米[24]等作物上的研究結論相一致。
植物根系不但是吸收養分和水分的器官,同時是同化、轉化或合成多種營養物質的重要場所[25],根系的生長發育和活力狀況對植物的抗逆境能力至關重要[21]。根系皮層、維管束對研究植物在干旱逆境下的生長特性具有重要的生物學價值[26];干旱脅迫試驗中頂果木的根半徑在逐漸降低,根系皮層和維管束厚度隨干旱脅迫程度的加重總體趨勢在變薄,可能是由于外界環境中離子濃度較高、水勢較低等一系列不利于植物吸收水分的環境影響下皮層變薄有利于水分吸收所導致的。根系維管束中木質部和韌皮部分別是運送水分和礦質元素的通道,其變薄有利于導管和篩管中水分、無機鹽與有機物的輸送,從而保證植物在干旱脅迫下植物體內的細胞仍可處在濕潤的環境下,進而維持正常的生長代謝。這個結果與田麗麗等[27]對類短命植物阿爾泰獨尾草的形態解剖研究結果相反,田麗麗等研究發現,阿爾泰獨尾草發達的皮層組織不僅可以儲存足夠的水分以應對干旱,而且還可能是營養物質的儲存場所。這可能是由于2種植物的地域分布不同而造成的抗逆境機制不同所產生的環境適生性作用的結果。
頂果木作為珍貴鄉土樹種,抗旱能力較弱;但該樹種在適當干旱條件下具有相當的調節能力,能夠維持植株在一定的生長水平上。因此,在發展種植頂果木過程中,盡管該植物適宜種植于南亞熱帶季風帶降雨量充足的地區,但也需要保證充足的水分供應。今后還需深入對該植物的葉脈結構和細胞超微結構等方面進行研究。
參考文獻:
[1] 朱積余,廖培來.廣西名優經濟樹種[M].北京:中國林業出版社,2006.89-90.
[2] 梁瑞龍,黃開勇.廣西熱帶巖溶區林業可持續發展技術[M].北京:中國林業出版社,2010.153.
[3] 胡鎮江.速生樹種——頂果木[J].云南林業,1982(4):24-25.
[4] 李干善.石山造林綠化樹種引種簡介[J].廣西林業,1992(5):22.
[5] 朱積余,侯遠瑞,劉 秀.廣西巖溶地區優良造林樹種選擇研究[J].中南林業科技大學學報(自然科學版),2011,31(30):81-85.
[6] 楊成華.速生珍稀樹種頂果木[J].貴州林業科技,1989,17(2):59-61.
[7] 何關順,文 寶,何廣瓊.鄉土速生樹種頂果木育苗技術[J].廣西林業,2008(5):32-34.
[8] 黎素平,郭 曹,朱昌叁,等.頂果木高產栽培技術[J].現代農業科技,2011(14):229-230.
[9] 周傳明,秦武明,呂曼芳,等.頂果木離體培養研究[J].安徽農業科學,2012,40(3):1457-1458.
[10] 裴宗平,余莉琳,汪云甲,等.4種干旱區生態修復植物的苗期抗旱性研究[J].干旱區資源與環境,2014,28(3):204-208.
[11] 韓 剛,趙 忠.不同土壤水分下4種沙生灌木的光合光響應特性[J].生態學報,2010,30(15):4019-4026.
[12] 李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000.
[13] BUSCH F, HUNTER N P A,ENSMINGER I. Biochemical constrains limit the potential of the photochemical reflectance index as a predictor of effective quantum efficiency of photosynthesis during the winter-spring transition in Jack pine seedlings[J]. Functional Plant Biology,2009,36(11):1016- 1026.
[14] WILLIAMS M H, ROSENQVIST E, BUCHHAVE M. Response of potted miniature roses (Rosa × hybrida) to reduced water availability during production[J]. Journal of Horticultural Science and Biotechnology, 1999, 4:301-308.
[15] JIANG Y, HUANG B. Drought and heat stress injury to two cool season turf grasses in relation to antioxidant metabolism and lipid peroxidation[J]. Crop Science, 2001, 41: 436-442.
[16] ZHANG J, KIRKHAM M B. Antioxidant responses to drought in sunflower and sorghum seedlings[J]. New Phytologist, 1996, 132: 361-373.
[17] HURNG W P, KAO C H. Effect of flooding on the activities of some enzymes of activated oxygen metabolism, the levels of antioxidants, and lipoid per oxidation in senescing tobacco leaves[J]. Plant Growth Regulation, 1994, 14: 37-44.
[18] 葉冰瑩,陳由強,朱錦懋.水分脅迫對三種木麻黃小枝活性氧傷害的研究[J].福建師范大學學報(自然科學版),2000,16(1):76-79.
[19] 張智猛,戴良香,宋文武,等.干旱處理對花生品種葉片保護酶活性和滲透物質含量的影響[J].作物學報,2013,39(1):133-141.
[20] 周 建,楊立峰,郝峰鴿,等.低溫脅迫對廣玉蘭幼苗光合及葉綠素熒光特性的影響[J].西北植物學報,2009,29(1):136-142.
[21] 李妮亞,高俊鳳,汪沛洪.小麥幼芽水分脅迫誘導蛋白的特征[J].植物生理學報,1998,24(1):65-71.
[22] 劉靈娣,李存東,孫紅春,等.干旱對不同鈴重基因型棉花葉片細胞膜傷害、保護酶活性及產量的影響[J].棉花學報,2009, 21(4):296-301.
[23] 楊華庚,顏速亮,陳慧娟,等.干旱脅迫對中粒種咖啡幼苗膜脂過氧化、抗氧化酶活性和滲透調節物質含量的影響[J].熱帶作物學報,2014,35(5):944-949.
[24] 張仁和,鄭友軍,馬國勝,等.干旱脅迫對玉米苗期葉片光合作用和保護酶的影響[J].生態學報,2011,31(5):1303-1311.
[25] 張偉明,孟 軍,王嘉宇.生物炭對水稻根系形態與生理特性及產量的影響[J].作物學報,2013,39(8):1445-1451.
[26] 陳選陽,鄭金貴,袁照年.類胡蘿卜素生物合成代謝工程研究進展[J].福建農業大學學報,2003(3):348-352.
[27] 田麗麗,馬 淼.類短命植物阿爾泰獨尾草的解剖學研究[J].植物研究,2013,45(2):56-60.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
