洪澤湖浮游藻類變化動態及影響因素
2016-10-19 10:09:46舒衛先張云舒韋翠珍
水資源保護 2016年5期
關鍵詞:水質
舒衛先,張云舒,韋翠珍
(1.淮河流域水資源保護局,安徽 蚌埠 233001; 2.南京大學生命科學學院, 江蘇 南京 210023)
?
洪澤湖浮游藻類變化動態及影響因素
舒衛先1,張云舒2,韋翠珍1
(1.淮河流域水資源保護局,安徽 蚌埠233001; 2.南京大學生命科學學院, 江蘇 南京210023)
針對2011年7月至2013年10月在洪澤湖11個采樣點20次采樣獲得的湖水表層浮游藻類和水質理化指標數據,采用Shannon-Wiener指數H和優勢度指數Y進行數據統計處理,分析洪澤湖浮游藻類時空分布動態及其影響要素。結果表明:洪澤湖浮游藻類共有7門60屬144種,主要包括綠藻門66種、硅藻門36種、藍藻門23種、裸藻門13種。浮游藻類群落具有明顯的時空異質性,物種豐富度夏季最高,秋季中等,冬春季最低;西北部(成子湖區)和河流入湖口(溧河洼)高,湖心和河流出湖口(蔣壩)低;浮游藻類細胞密度全湖平均值變幅為157萬~604萬個/L,夏秋高,冬春--低;成子湖區等靜水區高,入湖和出湖口低。浮游藻類組成和細胞密度的時間動態與溫度、水位及營養鹽(TN、TP等)的季節差異有關,而其空間動態與水動力因素和營養鹽(NH3-N)的空間差異有關。建議限制洪澤湖營養鹽陸地輸入,合理調控洪澤湖水動力,以遏制洪澤湖藍藻水華的發生。關鍵詞:浮游藻類;空間差異;季節動態;水質;洪澤湖
洪澤湖(33°06′~33°40′N,118°10′~118°52′E)是我國第4大淡水湖泊,是南水北調東線工程的過水通道,主要由江蘇盱眙縣至洪澤縣淮河河段及其北岸的溧河洼、安河洼、成子湖3大湖灣組成。在正常蓄水位13.0 m(蔣壩水位站)時,湖區水面面積為2 151.9 km2;洪澤湖承泄淮河上中游15.8萬km2的來水,上承支流河道主要在湖西,下泄河道都在湖東[1]。洪澤湖是典型的過水性、大型淺水湖泊,浮游生物、底棲動物和魚類等資源豐富,具有很高的生物多樣性保護價值[2]。20世紀80年代以來,隨著流域經濟的迅速發展和人口的不斷增加,該湖泊富營養化趨勢日益明顯,水華現象頻繁出現,尤其是90 年代初期至中期,水質急劇惡化[3]。洪澤湖不斷加劇的水體富營養狀況,使水生生物群落和物種多樣性都發生了很大變化[4],其中浮游藻類的變化尤為明顯[5]。研究洪澤湖浮游藻類群落結構和物種多樣性的時空變化動態,有利于了解洪澤湖生態系統現狀,為湖泊水生態恢復提供科學依據。
洪澤湖浮游藻類多樣性研究最早始于20世紀60年代,后來又有多次的野外調查和分析[5-6],但這些成果缺乏系統全面的時空格局研究。首先,以往的研究通常只選擇在生長季節(4—9月)調查取樣,采樣周期較短,取樣持續時間也不夠連貫[5]。其次,取樣點分布不夠廣,范圍較為局限,浮游藻類的空間變化特征不清楚[6]。更為重要的是,連續性的調查研究時間大都在2000年以前,而近十幾年洪澤湖水質、浮游藻類多樣性和密度都有較大改變[7],因此有必要對洪澤湖浮游藻類開展新一輪的詳細監測,以進一步了解湖泊的生態現狀。基于此,筆者開展了為期兩年多的詳細監測,分析了浮游藻類群落結構和細胞密度的時空動態,并結合水體理化性質,探討藻類動態與水位和水質特征的關系。研究結果有助于認識洪澤湖富營養化的格局和形成機制,以期為洪澤湖的開發利用和生態保護提供理論依據。
1 研究方法
1.1采樣點
根據洪澤湖湖盆特點,結合支流位置、植被和水質狀況,選定了11個采樣點,采樣點涵蓋成子湖灣、溧河湖灣、淮河湖灣以及湖中心區等水域,且反映了洪澤湖不同的水動力特征(圖1)。其中龍集北、龍集北偏北、成河北和成河東位于成子湖區,屬靜水區;溧河洼、臨淮、成河西位于溧河湖灣;高良澗位于二河閘,靠近蘇北灌溉總渠的出湖口;老子山處于淮河湖灣,靠近淮河入河口;蔣壩位于入江水道出湖口區,靠近三河閘;成河中位于洪澤湖中心區域。

圖1 洪澤湖采樣點分布
1.2樣品采集與處理
分別于2011年7月到2012年6月、2013年3—10月采集洪澤湖11個采樣點的浮游藻類。其中,浮游藻類定性標本用25號浮游生物網在表層水中撈取,加甲醛溶液(含37%~40%甲醛溶液,用量為水樣體積的4%)固定,帶回室內鑒定種類;種類鑒定根據光學顯微鏡下形態學特征參考相關文獻和書籍進行鑒定[8-9]。對于定量樣品,用8L的采水器在水深0.5 m處采集水樣,經P32(20 μm)號微型浮游生物網過濾后倒入標本瓶中,加魯哥氏液固定(同上),帶回實驗室后直接鏡檢,每個采樣點重復5次[10]。浮游藻類計數方法為目鏡行格法,用0.1 mL浮游藻類計數框在倒置顯微鏡100或400倍下進行測定。每次觀察100個視野,每個樣品計數2片,取其平均值作為該樣品的最終結果。另外,在每次采樣檢測過程中,對每一種藻隨機選取10個以上的個體,直接計數組成個體的細胞數量,將平均值作為該種藻的細胞數。
1.3環境因子測定
水溫和pH值采用YS1-85水質分析儀(美國)現場原位監測;TN、NH3-N、TP和COD通過采集表層下 0.5 m 處水樣2.5 L帶回實驗室進行測定,樣品測定重復3次,測定方法分別采用堿性過硫酸鉀消解分光光度法(HG 636—2012)、納氏試劑光度法(GB7479—1987《水質 銨的測定 納氏試劑比色法》)、鉬酸銨分光光度法(GB11893—1989《水質 總磷的測定 鉬酸銨分光光度法》)和標準重鉻酸鉀法(GB11914—1989《水質 化學需氧量的測定 重鉻酸鹽法》)測定;水體DO采用便攜式DO測定儀進行測定(Hach HQ40d oxygen probe,美國哈希公司);水位數據來自淮安市水文信息網(http://www.haswj.com,蔣壩水位站數據)。但2013年度TN含量測定因為儀器問題,未能獲取數據。
1.4數據分析方法
浮游藻類群落指數采用Shannon-Wiener指數H[11]和優勢度指數Y:
(1)
(2)
式中:s為種數;ni為第i種的個體數;N為全部樣品中的總個體數;fi為第i種在各樣品中出現的頻率。
多重比較采用Duncan 檢驗,顯著度水平為 0.05。因子間相關分析采用雙變量相關分析,選擇Pearson 相關系數。并用線性回歸的方法探討水溫對浮游藻類總密度的影響。數據處理采用統計軟件 SPSS19.0。

表1 2011—2013年洪澤湖浮游藻類種類組成 種
2 結果與分析
2.1環境參數
2.1.1水溫與水位
洪澤湖水溫季節波動明顯,夏季水溫高,冬季水溫低,春季和秋季中等。2011—2012年度2月水溫最低(3.51℃),8月最高(29.08℃),2013年度 3月最低(9.14℃),8月最高(31.48℃)。
2011—2012年度,平均水位為12.89 m, 2011年7月較低,8月隨雨季來臨水位開始上升,9月達到最高值為13.43 m,此后水位逐漸下降至次年6月為止(11.92 m)。2013年度(4—11月),平均水位為12.48 m,3月水位最高達12.97 m,9月初水位達到最低值,為11.6 m。
2.1.2化學性質
2011—2012年度監測表明,洪澤湖水質偏堿性(pH值為7.81~8.43),不同月份不同樣點間變化不明顯。全湖DO質量濃度變化為4.0~14.3 mg/L,夏秋低,冬春高;空間上,靜水區成子湖區域相對較低,成河西、老子山、溧河洼及臨淮中等,而河流出湖區成河中、蔣壩及高良澗區域最高。7月水體NH3-N質量濃度最高,達0.70 mg/L,其他月份變幅較小;在空間上,靜水水域龍集北偏北監測點最高為0.59 mg/L,河流入湖口老子山、溧河洼兩地適中,分別為0.41 mg/L和0.43 mg/L,而其他監測位點NH3-N質量濃度相對較低(變幅0.16~0.36 mg/L)。TP平均質量濃度變化范圍為0.04~0.11 mg/L,4月最低,11月最高;空間上,溧河洼較高為0.22 mg/L,其他監測位點TP質量濃度較低,不同位點間差異較小。全湖TN質量濃度的變化范圍為0.90~3.37 mg/L,夏秋低(2011年7—10月),冬春高 (2011年11月至2012年1月);空間上,淮河入湖口老子山水域含量最高,蔣壩其次,臨淮、龍集北和成河西最低。
2013年監測結果與上一年度相似。水體pH值范圍在7.86~8.25間,偏堿性;DO質量濃度變化范圍為5.30~11.12 mg/L,3月最高,此后逐漸降低,至8月為最低(6.88 mg/L),不同位點之間DO質量濃度差異較小。NH3-N質量濃度變化范圍為0.20~0.48 mg/L,9月最高,4月最低;空間上,成子湖區域相對較高,范圍在0.31~0.82 mg/L,高良澗、蔣壩、成河中等出湖口區域NH3-N質量濃度較低;TP質量濃度變化范圍為0.05~0.12 mg/L,6月最低,10月最高;不同監測位點間空間差異較小。
2.2浮游藻類物種組成
2011—2013年調查期間,共檢測到綠藻、硅藻、藍藻、裸藻、甲藻等浮游藻類,隸屬7門60屬144種(表1),其中,弱細顫藻(優勢度指數0.24)、不定微囊藻(優勢度指數0.17)和水華微囊藻(優勢度指數0.07)為全湖優勢物種。
2011年7月至2012年6月間,觀測到的綠藻門種類最多,達66種,占全部浮游藻類種類的50.4%,是最重要的群落組成者;硅藻門其次(36種),占總數的27.5%;藍藻門16種,裸藻門8種,黃藻門3種,甲藻門2種,分別占總數的12.2%、6.1%、2.3%和1.5%。具體看,顫藻(Oscillatoria)為絕對優勢屬種,其相對密度為60.9%,其次依次有:微囊藻(Microcvstis)21.3%、鏈絲藻(Ulothrix)14.4%、魚腥藻(Anabaena)1.2%。
2013年3—10月間,主要為綠藻、硅藻和藍藻3個類群。其中綠藻門49種,硅藻門29種,藍藻門23種,分別占總數的41.5%、24.6%和19.5%。此外還有少量裸藻和甲藻,其中裸藻門13種,甲藻門3種,分別占總數的11.0%和2.5%。
浮游藻類群落物種組成,在季節變化上,表現為夏秋種類多,冬春少。第1個監測周期內,除3月外,各月浮游藻類種類中均以綠藻門為主,占浮游藻類總種數的43.10%~64.71%;硅藻門和藍藻門次之。第2個監測周期內,3—5月綠藻門和硅藻門種類數基本相同,其次為藍藻門;6—10月浮游藻類基本都以綠藻門為主,其次為硅藻門和藍藻門。
在空間上,成子湖區(含龍集北、龍集北偏北、成河北和成河東等采樣點)及溧河洼(為入湖口)藻種類較多,而蔣壩、老子山、高良澗(為出湖口)及湖區中心(成河中)較低(表2),各個監測位點均以綠藻門的浮游藻類最多。第1個監測周期內,成子湖區藻類較多,種類最少的樣點為成河中和高良澗;各樣點中,綠藻門占浮游藻類總種數的30.2%~54.1%,硅藻門、藍藻門次之。第2個監測年度內,溧河洼藻種類最多,成子湖水域次之,老子山最少(49種),各樣點中綠藻門占浮游藻類總種數的38.2%~44.1%;硅藻門和藍藻門分別占浮游藻類總種數的2.8%~30.3%和18.8%~30.6%。
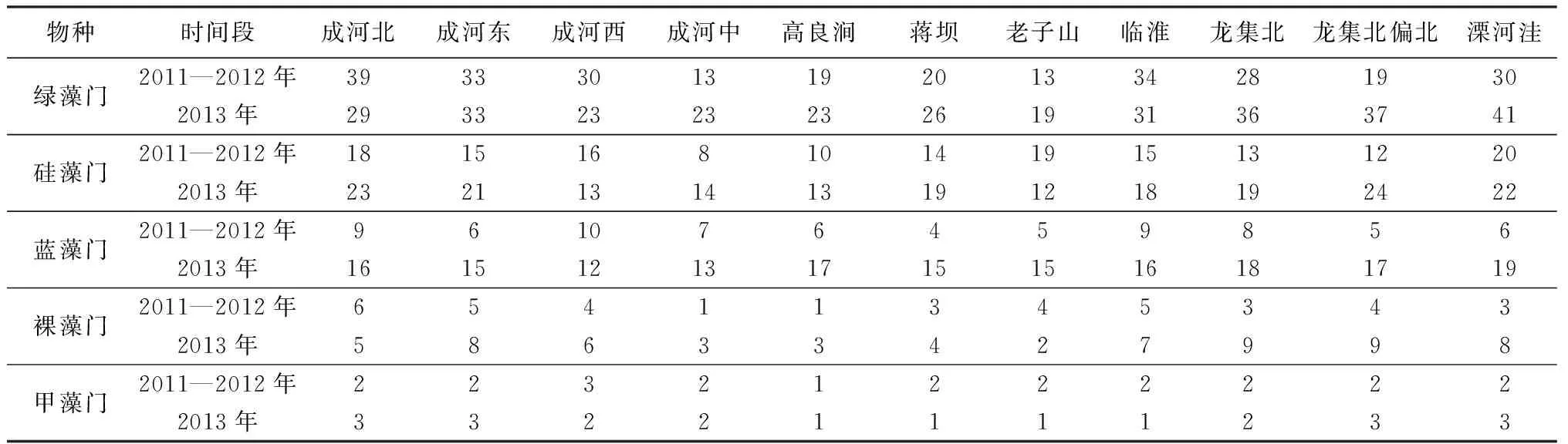
表2 洪澤湖浮游藻類群落物種組成空間動態 種

表3 2011—2013年度洪澤湖浮游藻類細胞密度空間動態 萬個/L
2.3浮游藻類細胞密度
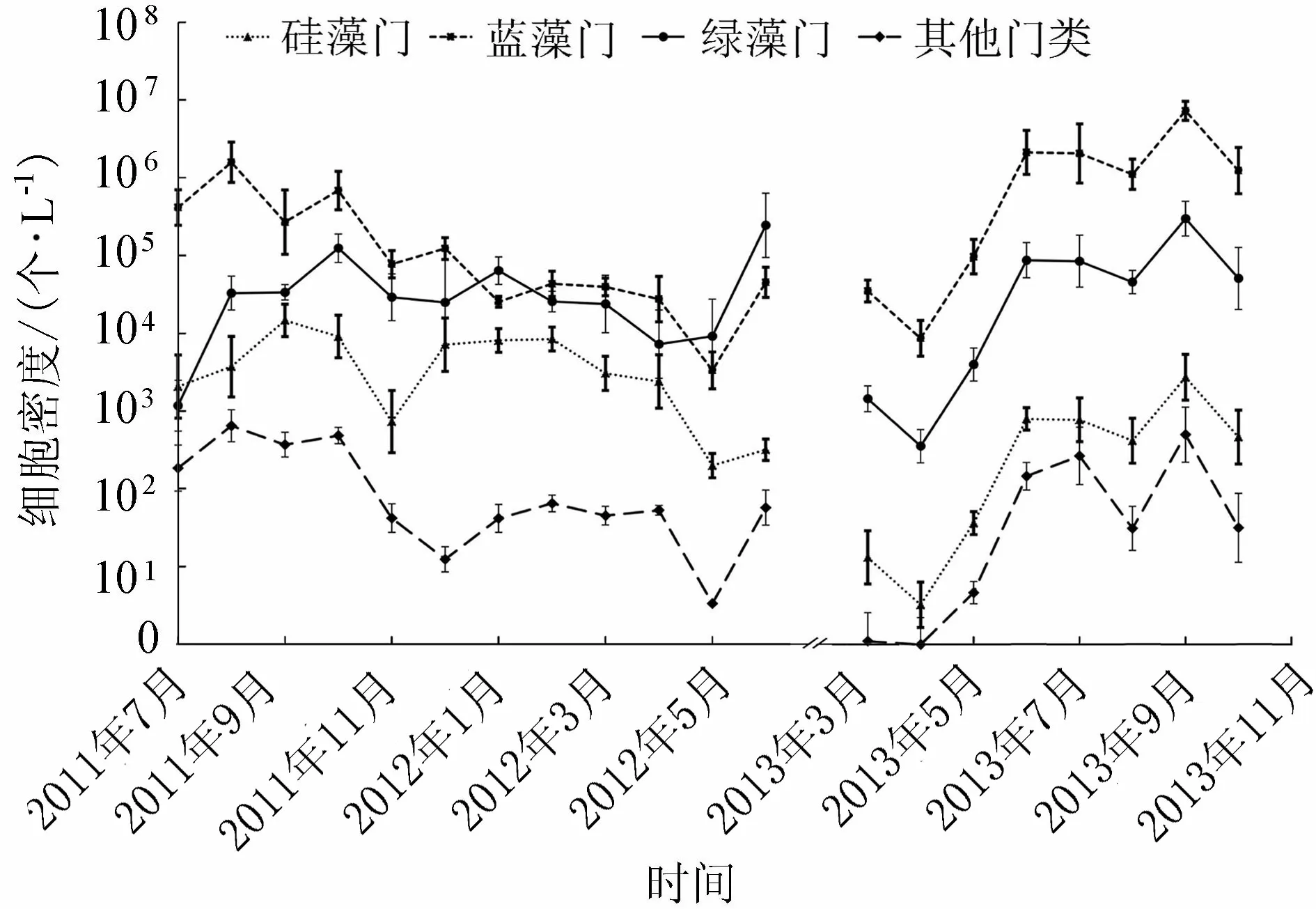
圖2 2011—2013年洪澤湖浮游藻類細胞密度時間動態
浮游藻類細胞密度夏秋多,冬春少(圖2)。第1個監測周期內,平均細胞密度為157萬個/L,各月間差異很大,其中2011年8月細胞密度最大,主要由微囊藻和弱細顫藻的快速生長繁殖引起;10月后細胞密度逐漸下降,至第2年春天溫度升高,鏈絲藻開始快速生長繁殖,引起了2012年6月細胞密度的迅速增長。第2個監測周期內,浮游藻類細胞密度平均值約為604萬個/L,其中, 4月前細胞密度較低;5月開始增加,6—7月浮游藻類細胞密度達到較高水平,其增長主要由弱細顫藻的大量繁殖引成,9月出現當年峰值(1 090萬個/L),此時微囊藻取代弱細顫藻成為全湖優勢物種。
不同位點間浮游藻類細胞密度差異非常明顯(表3),在水流相對較緩的成子湖區(包括龍集北、龍集北偏北、成河北和成河東)最高,溧河洼、成河西和臨淮次之,而強水動力區(主要為湖泊入湖和出湖口等,包括蔣壩、老子山、高良澗)最低。具體而言,第1個監測周期內龍集北偏北最高,湖心采樣點成河中最低;第2個周期內成河北和成河東最高,臨淮、溧河洼、成河西浮游藻類中等,老子山、蔣壩和高良澗最低。

表4 2011—2013年洪澤湖浮游藻類物種多樣性的空間動態
2.4物種多樣性
洪澤湖浮游藻類多樣性指數存在明顯的時空動態,時間上表現為秋季多樣性指數較高,冬春季較低其中,2011—2012年間,整個湖區各月浮游藻類多樣性指數在0.77~1.84之間波動(圖3),平均值為1.31,9月最高為1.84,6月最低為0.77,而2013年各月多樣性指數變化范圍為0.91~2.16,平均值為1.32,同樣為9月最高為2.16,6月最低為0.91。空間上,成子湖區域多樣性指數較低(表4),溧河洼水域(成河西、臨淮、溧河洼)較高。
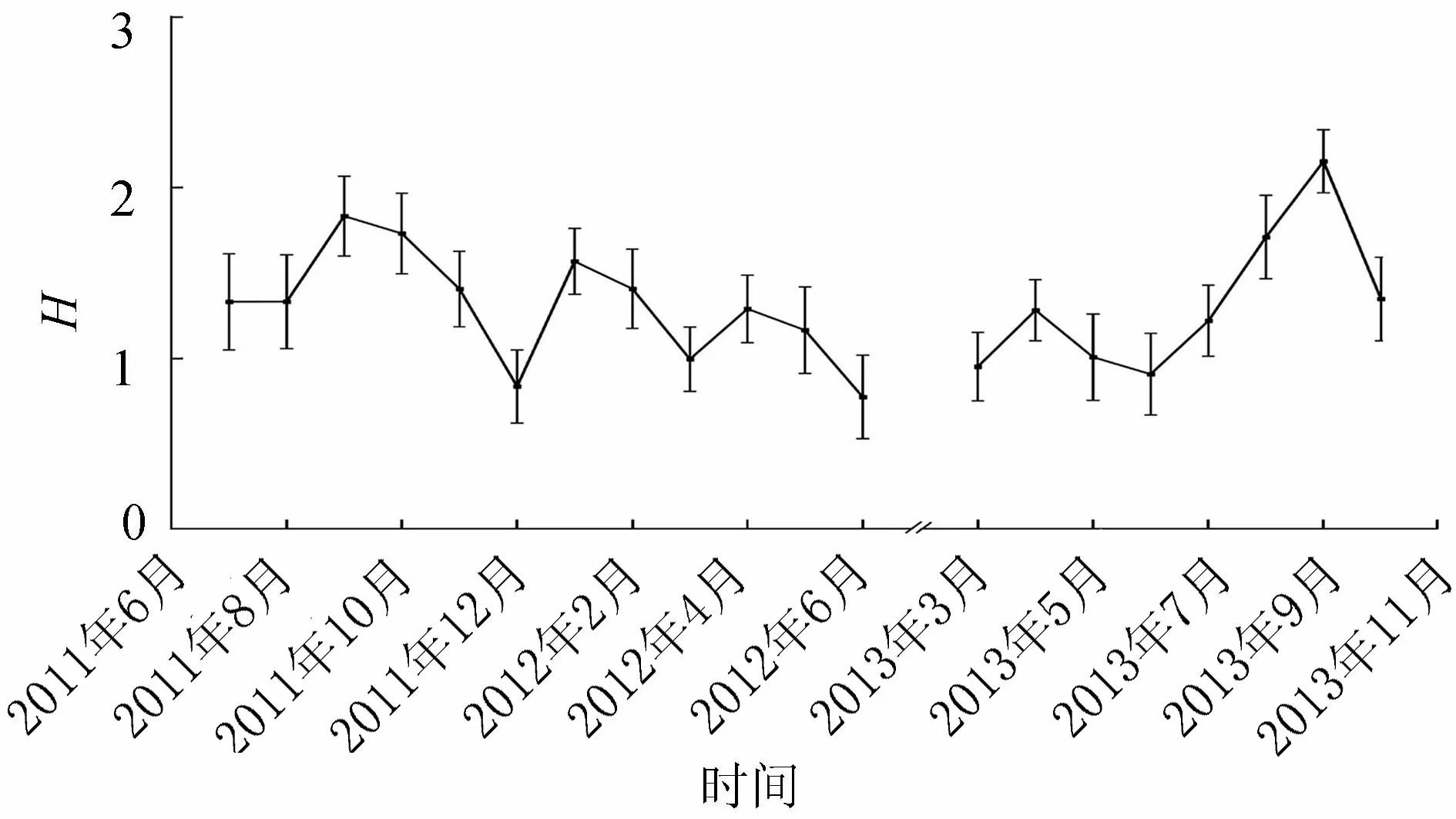
圖3 2011—2013年洪澤湖浮游藻類物種多樣性的時間動態
2.5細胞密度與環境因子關系
由表5可見,pH值與生物學指標間沒有顯著關系,藍藻細胞密度、其他門類細胞密度及總細胞密度與水溫度呈正相關,總細胞密度與藍藻細胞密度變化顯著正相關。具體而言,第1個監測周期內水溫和藍藻門及總細胞密度正相關,DO和藍藻門細胞密度及總細胞密度負相關;第2個監測周期內NH3-N和硅藻門、綠藻門正相關。
3 討 論
上述分析結果表明,洪澤湖在兩個監測周期中浮游藻類年平均細胞密度分別達到157萬個/L和604萬個/L,與2008年相比(小于85萬個/L),2011年細胞密度增長近1倍,2013年度也明顯超過湖泊水華閾值(大于100萬個/L)[17], 說明洪澤湖的部分水域在一定時間范圍內處于水華發生狀態。這與洪澤湖含較高的氮、磷結果是一致的。2011—2013年洪澤湖NH3-N、TN和TP平均質量濃度分別為0.29 mg/L、2.19 mg/L和0.08 mg/L,超出了Taheriyoun等[12]提出的湖泊富營養化閾值ρ(NH3-N):0.02 mg/L;ρ(TN):0.65~1.2 mg/L;ρ(TP):0.03~ 0.1 mg/L)。N、P是與藻類生長密切相關的營養元素,過高的N、P含量往往導致水華出現[13]。另外,雖然洪澤湖綠藻、硅藻種類在全年都較多,但藍藻門

表5 洪澤湖浮游藻類細胞密度和多樣性與水體物化特征關系的半矩陣
注:*表示P<0.05; **表示P<0.01;***表示P<0.001。
細胞密度與全年總細胞密度相關性最高,說明藍藻在全年浮游藻類細胞密度中具有較大貢獻。進一步分析發現,顫藻和微囊藻是洪澤湖的絕對優勢物種,兩者占總密度的80%以上。顫藻和微囊藻是淺水湖泊中兩種最典型的藍藻[13]。比如在太湖,微囊藻和顫藻水華都有發生,其中微囊藻全年發生,而顫藻一般在夏秋兩季暴發[14]。可見,洪澤湖富營養化狀態與多數淺水湖泊一致,水華的優勢物種都是藍藻[15-16]。雖然如此,與滇池(細胞密度為520萬~12億個/L[17];ρ(N)=2.0 mg/L,ρ(P)=0.21 mg/L[18])和太湖五里湖 (細胞密度386萬~558萬個/L[19];ρ(N)=7.2g/L和ρ(P)=0.15 mg/L[20]) 相比,洪澤湖的藻細胞密度和營養鹽濃度相對較低。但是,如果不及早重視洪澤湖的生態恢復和治理,其富營養化趨勢將難以得到遏制。
洪澤湖浮游藻類物種組成和細胞密度存在明顯的季節變化,這與溫度的季節性差異有關。溫度是藻類生長的重要限制因素,藻類在低溫季節難以生長。洪澤湖湖區屬北亞熱帶季風氣候,季節變化十分明顯。因此,在水溫較高的夏季6—8月,2011—2013年藻細胞密度超過水華閾值的1.4~20.3倍;而在水溫較低的3—5月,細胞密度不足水華閾值的1/3。相關分析表明,水溫和全湖各月細胞密度之間存在很好的線性正相關關系,說明水溫是導致洪澤湖細胞密度季節變化的重要環境因素。水溫的變化同樣會改變浮游藻類的群落組成。與其他浮游藻類相比,藍藻生長需要更高的溫度(其最適生長溫度為25~35℃[5])。在較高溫度下藍藻相比其他藻類更易取得競爭優勢[21]。一方面,高溫條件下藍藻分泌的次生代謝物(藻毒素)含量的增加抑制了浮游動物和其他浮游藻類的生長[10];另一方面,藍藻的快速生長,造成水體透明度下降,遮蔽光線而進一步抑制其他水生植物的生長(如沉水植物等),甚至導致其他水生植物死亡,死亡釋放的養分又提供了藍藻生長所需要的養分。因此,夏秋季節水溫高時,藍藻相對密度最高,隨著冬季和春季水溫降低,藍藻競爭抑制作用減弱,其他藻類(主要為綠藻和硅藻)的相對密度得到提高。
除了溫度,水位可能也是細胞密度季節變化的重要調節因子。水位變化說明水體容積發生了變化,從而直接通過“濃縮”或“稀釋”作用影響藻密度。例如,2012年6月、2013年7月和9月細胞密度驟然提升,可能與這個幾個月出湖流量增加、水位下降有關。此外,N、P等養分濃度的波動也引起藻類細胞密度變化。P含量相對較高的夏季和秋季藍藻細胞密度及總浮游藻類細胞密度相對較高,隨著冬季和春季P濃度降低,藍藻生長受到限制,綠藻逐漸取得競爭優勢,相對密度不斷增加。可見,洪澤湖浮游藻類的季節變化可能是溫度、水位和養分等要素共同作用的結果。
洪澤湖浮游藻類物種組成和細胞密度在空間上的差異也很明顯。浮游藻類物種數量在不同區域明顯不同,對比分析發現,湖區西部(如臨淮、成河西,以及溧河洼)離入湖口區域較近的樣點浮游藻類物種較多,主要是受入湖水流帶來的上游河流的浮游藻類物種的影響,而在出湖口一側(如蔣壩等)的東部湖區浮游藻類種類較少,原因可能跟出湖區水流速度較快不利于藻類的生長繁殖有關[22]。浮游藻類種類組成在區域上差異較小,均以綠藻為主,藍藻、硅藻次之。
同樣,細胞密度的空間差異也跟水動力要素有關。浮游藻類細胞密度最高的監測位點在成子湖水域,其中2011—2012年龍集北偏北夏季和秋季藻細胞密度分別超過湖泊水華閾值13.5和8.8倍,2013年成河北、成河東、龍集北偏北均超過湖泊水華閾值10倍以上;而細胞密度較低的監測點位是成河中和老子山,其藻類細胞密度水平通常低于水華閾值,甚至處于貧營養狀態(閾值小于30萬個/L)[9]。老子山處在淮河的入湖口,蔣壩處在三河區出湖口,高良澗位于二河閘,靠近蘇北灌溉總渠的出湖口,這些位點過水性都很強。因此,不同采樣點藻細胞密度的差異可能與洪澤湖獨特的水動力特征有關,即過水性越強、水流速度越大,越不容易發生水華[22]。此外,從整體上看,洪澤湖為一個典型的過水性湖泊,年換水在10次以上,正因為其徑流量大,流速較快,降低了藍藻細胞密度的增加速度,所以相比滇池和太湖五里湖等過水性相對較弱的富營養化湖泊,其藻類細胞密度要低。
水體營養鹽濃度也是影響浮游藻類細胞密度時空分布的重要因素。雖然整合數據的分析表明,營養鹽濃度與植物細胞密度沒有顯著相關關系,而且TN與總細胞密度呈顯著負相關。這可能與養分和水溫等其他要素的相互作用有關。洪澤湖處在溫帶地區,夏秋季節溫度高,藻類分裂快,細胞密度高,但同時因為夏秋季節降水量較大、進出湖泊的河流凈流量大,營養鹽濃度反而降低。相反,冬春季節溫度低,不適宜藻類生長,細胞密度低,但冬春降水量小、水位低,水體營養鹽濃度高。類似負相關的結果在杭州西湖等其他水體中也有報道[23]。但是營養鹽濃度仍可能是影響藻類細胞密度分布的重要因素。例如,在成子湖等浮游藻類細胞密度最高的區域,NH3-N質量濃度都相對較高(0.42 mg/L),而在細胞密度相對較低的成河中、高良澗、蔣壩、老子山等監測點NH3-N質量濃度較低(0.22 mg/L)。NH3-N作為浮游藻類生長可直接利用的營養鹽,其空間差異可能是導致浮游藻類細胞密度空間現狀的重要因素。
4 結 語
調查結果顯示,洪澤湖夏秋季節局部水域有明顯的富營養化特征,在水動力較弱的成子湖區域藍藻水華現象尤為明顯。洪澤湖藍藻水華的發生是營養鹽、水溫和水動力特征共同作用的結果。夏秋季節的炎熱高溫、豐富的營養鹽含量以及較低的水流速度導致了成子湖等區域藍藻水華的大量爆發。需要指出的是,本次定量實驗采用的是網篩過濾法,雖然該方法可能會低估水體當中小于20 μm 的浮游藻類的細胞密度,但即使如此,洪澤湖浮游藻類群落組成和細胞密度仍然存在明顯的時空動態,表現出富營養化加劇的趨勢,因此,洪澤湖富營養化問題更應該引起人們的重視。
南水北調東線工程通水后,水位抬高,水動力條件改變,可能會影響洪澤湖,特別是成子湖灣區域,浮游藻類時空格局的變化,因此需要開展持續的觀測研究。進一步加大洪澤湖入湖河流污染治理和湖區退耕還湖力度,逐步減少污染物輸入,從而降低洪澤湖營養鹽含量,是遏制洪澤湖富營養化加劇的根本措施。
[ 1 ] 《中國河湖大典》編纂委員會.中國河湖大典 淮河卷[M].北京:中國水利水電出版社,2010.
[ 2 ] 楊士建.洪澤湖濕地資源保護與可持續利用研究[J].重慶環境科學,2003,25(2):15-17.(YANG Shijian.Study on protection and sustainable use of wetland around Hongze Lake[J].Chongqing Environmental Sciences,2003,25(2):15-17.(in Chinese))
[ 3 ] 李波,濮培民.淮河流域及洪澤湖水質的演變趨勢分析[J].長江流域資源與環境,2003,12(1):68-73.(LI Bo,PU Peimin.Study on the evolution tendency of water quality in Huai River Basin and Hongze Lake[J].Resources and Environment in the Yangtze Basin,2003,12(1):68-73.(in Chinese))
[ 4 ] 葛緒廣,王國祥.洪澤湖面臨的生態環境問題及其成因[J].人民長江,2008,39(1):28-30.(GE Xuguang,WANG Guoxiang.Eco-environmental problems in Hongze lake and their causes of formation[J].Yangtze River,2008,39(1):28-30.(in Chinese))
[ 5 ] 王兆群,張寧紅,張詠.洪澤湖藻類與環境因子逐步回歸統計和藍藻水華初步預測[J].中國環境監測,2012,28(4):17-20.(WANG Zhaoqun,ZHANG Ninghong,ZHANG Yong.Prediction of blue-green algae bloom using stepwise multiple regression between algae & related environmental factors in Hongze Lake[J].Environmental Monitoring in China,2012,28(4):17-20.(in Chinese))
[ 6 ] 陳翔,禹繼華,劉杰,等.洪澤湖藍藻暴發的氣象條件分析[J].安徽農業科學,2010,38(15):8141-8142,8177.(CHEN Xiang,YU Jihua,LIU Jie,et al.Analysis on the meteorological condition of blue-green algae fast growth in Hongze Lake[J].Journal of Anhui Agricultural Science,2010,38(15):8141-8142,8177.(in Chinese))
[ 7 ] 王兆群,張寧紅,張詠,等.洪澤湖水質富營養化評價[J].環境監控與預警,2010,2(6):31-35.(WANG Zhaoqun,ZHANG Ninghong,ZHANG Yong,et al.Eutrophication assessment of water quality in Hongze Lake[J].Environmental Monitoring and Forewarning,2010,2(6):31-35.(in Chinese))
[ 8 ] 周鳳霞,陳劍虹.淡水微型生物圖譜[M].北京:化學工業出版社,2005.
[ 9 ] DAVIS T W,BERRY D L,BOYER D L,et al.The effects of temperature and nutrients on the growth and dynamics of toxic andnon-toxic strains of Microcystis during cyanobacteria blooms[J].Harmful Algae,2009,8:715-725.
[10] 張婷,李林,宋立榮.熊河水庫浮游藻類群落結構的周年變化[J].生態學報,2009,29(6):2971-2979.(ZHANG Ting,LI Lin,SONG Lirong.Annual dynamics of phytoplankton abundance and community structure in the Xionghe Reservoir[J].Acta Ecologica Sinica,2009,29(6):2971-2979.(in Chinese))
[11] SHANNON C E,WIENER W.The mathematical theory of communication[M].Urbana:University Illinois Press,1963.
[12] TAHERIYOUN M,KARAMOUZ M,BAGHVAND A.Development of an entropy-based fuzzy eutrophication index for reservoir water quality evaluation[J].Iranian Journal of Environmental Health Science & Engineering,2010(7):1-14.
[13] CHU Z S,JIN X C,IWAMI N,et al.Eutrophication of Shallow Lakes with Special Reference to Lake Taihu,China[M].Berlin:Springer,2007.
[14] 張軍毅,朱冰川.太湖五里湖藍藻水華種類及其演替規律研究[C]//中國環境科學學會.2012中國環境科學學會學術年會論文集(第2卷).北京:中國農業大學出版,2012:621-625.
[15] 秦伯強.太湖水環境面臨的主要問題、研究動態與初步進展[J].湖泊科學,1998,10(4):1-9.(QIN Boqiang.A review and prospect about the aquatic environment studies in Taihu Lake[J].Journal of Lake Sciences,1998,10 (4):1-9.(in Chinese))
[16] 秦伯強.長江中下游淺水湖泊富營養化發生機制與控制途徑初探[J].湖泊科學,2002,14(3):193-202.(QIN Boqiang.Approaches to mechanisms and control of eutrophication of shallow lakes in the Middle and Lower Reaches of the Yangze River[J].Journal of Lake Sciences,2002,14(3):193-202:1-9.(in Chinese))
[17] 李原,張梅,王若南.滇池的水華藍藻的時空變化[J].云南大學學報(自然科學版),2005,27(3):272-276.(LI Yuan,ZHANG Mei,WANG Ruonan.The temporal and spation variation of the cyanobacteria which caused the water bloom in the Dianchi Lake,Kunming,China[J].Journal of Yunnan University(Natural Sciences),2005,27 (3):272-276.(in Chinese))
[18] 張治中.滇池氮與富營養化研究[J].環境科學導刊,2007,26(6):34-36.(ZHANG Zhizhong.Study on nitrogen and eutrophication of Dianchi Lake[J].Environmental Science Survey,2007,26(6):34-36.(in Chinese))
[19] 孟順龍,陳家長,范立民,等.2007年太湖五里湖浮游藻類生態學特征[J].湖泊科學,2009,21(6):845-854.(MENG Shunlong,CHEN Jiazhang,FAN Limin,et al.Eco-characteristics of phytoplankton in Lake Wuli,Lake Taihu in 2007[J].Journal of Lake Sciences,2009,21(6):845-854.(in Chinese))
[20] 宋曉蘭,劉正文,潘宏凱,等.太湖梅梁灣與五里湖浮游藻類群落的比較[J].湖泊科學,2007,19(6):643-651.(SONG Xiaolan,LIU Zhengwen,PANG Hongkai,et al.Phytoplankton community structure in Meiliang Bay and Lake Wuli of Lake Taihu[J].Journal of Lake Sciences,2007,19(6):643-651.(in Chinese))
[21] NALEWAJKO C,MURPHY T P.Effects of temperature,and availability of nitrogen and phosphorus on the abundance of anabaena and microcystis in Lake Biwa,Japan:an experimental approach[J].Limnology,2001(2):45-48.
[22] 林秋奇,胡韌,韓博平.流溪河水庫水動力學對營養鹽和浮游藻類分布的影響[J].生態學報,2003,23(11):2278-2284.(LIN Qiuqi,HU Ren,HAN Boping.Effect of hydrodynamics on nutrient and phytoplankton distribution in Liuxihe Reservoir[J].Acta Ecologica Sinica,2003,23(11):2278-2284.(in Chinese))
[23] 毛成責,余雪芳,邵曉陽.杭州西湖總氮、總磷周年變化與水體富營養化研究[J].水生態學雜志,2010,3(4):1-6.(MAO Chengze,YU Xuefang,SHAO Xiaoyang.Study on the annual variations of TN and TP and the eutrophication in Hangzhou West Lake[J].Journal of Hydroecology,2010,3(4):1-6.(in Chinese))
Seasonal dynamics of and factors in phytoplankton in Hongze Lake
SHU Weixian1, ZHANG Yunshu2, WEI Cuizhen1
(1.HuaiheRiverBasinWaterResourcesProtectionBureau,Bengbu233001,China;2.SchoolofLifeSciences,NanjingUniversity,Nanjing210023,China)
In order to explore the temporal-spatial pattern of phytoplankton distribution in Hongze Lake and associated factors, we sampled surface water 20 times at 11 sites across the lake from July 2011 to October 2013, and obtained physical and chemical indices of phytoplankton and water quality. We processed the data using the Shannon-Wiener indexHand dominance indexY. Then, we found 144 phytoplankton species belonging to 60 genera, and seven phyla (including 66 species of green algae, 36 species of diatoms, 23 species of cyanobacteria, and 13 species of euglena). The phytoplankton distribution presented an obvious spatial-temporal variability. The species richness was at a high level in the summer, at an intermediate level in the autumn, and at low levels in the winter and spring; it exhibited large values in the northwestern part (Chengzi Lake area) and the inflow area of the lake (site Lihewa), and small values in the central area and the outflow area of the lake. The average cell density of phytoplankton ranged from 1.57×106to 6.04×106cells/L. The cell density was higher in the summer and autumn, and lower in the winter and spring; it exhibited large values in the Chengzi Lake area and small values at the inflow and outflow areas of the lake. Data analysis shows that the temporal dynamics of the species composition and cell density were significantly associated with the seasonal variations of water temperature, water level, and nutrients (mainly total nitrogen and total phosphorus), while the spatial dynamics of the species composition and cell density were correlated with the spatial variations of hydrodynamic factors and nutrients (particularly ammonia nitrogen). We suggest restricting nutrient input and modulating the hydrological dynamics of Hongze Lake, in order to prevent algal blooms in the lake.
phytoplankton; spatial disparity; seasonal dynamics; water quality; Hongze Lake
10.3880/j.issn.1004-6933.2016.05.022
淮河流域重要河湖健康評估試點項目( 1261320320004)
舒衛先(1973—),男,高級工程師,博士,主要從事水生態保護與修復管理工作。E-mail:weixians@163.com
X173
A
1004-6933(2016)05-0115-08
2015-11-29編輯:徐娟)
猜你喜歡
環境(2023年5期)2023-06-30 01:20:01
小星星·閱讀100分(低年級)(2022年6期)2022-08-22 02:33:40
供水技術(2021年3期)2021-08-13 09:08:34
建材發展導向(2019年10期)2019-08-24 06:25:02
當代水產(2019年1期)2019-05-16 02:42:04
當代水產(2019年3期)2019-05-14 05:42:48
電子制作(2018年14期)2018-08-21 01:38:16
水利規劃與設計(2018年1期)2018-01-31 01:53:53
水利規劃與設計(2016年7期)2016-02-28 15:06:27
世界文學評論(2014年2期)2014-04-12 06:23:43
