阿魏酸酯酶的研究與應用進展
2016-09-29 07:30:31王麗孫欽棟王賀祥
山東農業大學學報(自然科學版) 2016年4期
王麗,孫欽棟,王賀祥
1.山東農業大學植物保護學院,山東泰安2710182.中國農業大學生物學院,北京100193
阿魏酸酯酶的研究與應用進展
王麗1,2,孫欽棟1,王賀祥2*
1.山東農業大學植物保護學院,山東泰安271018
2.中國農業大學生物學院,北京100193
阿魏酸酯酶可以水解植物細胞壁中由阿魏酸、p-香豆酸及二聚阿魏酸等酚酸與半纖維素和木質素形成的酯鍵,高效降解木質纖維素的同時可以釋放出阿魏酸、p-香豆酸等抗氧化物質,在食品、造紙、醫藥、化妝品、生產生物乙醇等行業具有很高的應用價值和廣闊的市場前景。本文系統總結了阿魏酸酯酶的來源、理化性質、結構特征與催化機理及發酵生產、應用等方面的最新研究進展。
阿魏酸酯酶;研究進展;應用進展
阿魏酸酯酶(Feruloylesterase,EC 3.1.1.73)又稱為肉桂酸酯酶,屬于羧酸酯水解酶亞類,該酶可以水解植物細胞壁中由阿魏酸、p-香豆酸及二聚阿魏酸等酚酸與半纖維素和木質素形成的酯鍵,打破它們形成的網狀結構,將半纖維素和木質素分開,使纖維素酶可以與纖維素充分接觸,大大提高纖維素的降解率。阿魏酸酯酶在加速木質纖維素降解的同時還釋放出阿魏酸、p-香豆酸等抗氧化物質,在食品、造紙、醫藥、化妝品、生產生物乙醇等行業具有很高的應用價值和廣闊的市場前景[1]。圍繞阿魏酸酯酶的來源、分類、結構特征及催化機理,理化性質與應用等方面在近年來取得的最新進展進行了綜述分析。
1 阿魏酸酯酶的來源
由于在木質纖維素降解中的獨特功能,阿魏酸酯酶自從被發現后,一直是近些年研究的熱點。目前已從多種生物中分離純化,根據其來源,可以分為微生物阿魏酸酯酶和植物阿魏酸酯酶。
1.1微生物阿魏酸酯酶
微生物阿魏酸酯酶又分為真菌阿魏酸酯酶和細菌阿魏酸酯酶。
1.1.1真菌阿魏酸酯酶目前已發現的阿魏酸酯酶大部分都是真菌阿魏酸酯酶,這可能跟真菌的培養基質有關系。自然界的真菌主要生長在一些枯枝和草類的莖稈上,而阿魏酸酯酶是誘導酶,只有當培養基中含有木質纖維素類物質時才會被誘導產生。當培養基質改變時,有些真菌便不再產生阿魏酸酯酶,不同真菌的阿魏酸酯酶誘導培養基也是不同的。裂褶菌(Schizophyllum commune)在以纖維素為碳源的培養基中可以產生高活力的阿魏酸酯酶,而當以木聚糖和麥麩等木聚糖含量較高的底物作為碳源時則幾乎不產生阿魏酸酯酶。黑曲霉(Aspergillusniger)是目前阿魏酸酯酶研究最多的真菌,先后已經有六種阿魏酸酯酶從中分離純化出來,分別為FAE-I,FAE-II,AnFaeA,AnFaeB,FAE-1和FAE-2,它們的理化性質和底物特異性均不相同,其中FAE-I,FAE-II,AnFaeA和AnFaeB都是酸性阿魏酸酯酶,在酸性條件下較為穩定,而FAE-1和FAE-2則在堿性條件下比較穩定[2]。瘤胃真菌新美鞭菌屬Neocallimastix MC-2菌株可以產生兩種阿魏酸酯酶,分子量大小分別為69 kDa 和24 kDa,最適pH值都靠近中性,在40℃較穩定,其中一種酶對阿魏酰低聚糖和對香豆酰低聚糖都有水解作用,而另一種酶只對阿魏酰低聚糖具特異性,研究表明瘤胃真菌產生的阿魏酸酯酶能加快植物細胞壁等成分在瘤胃的降解速度,提高反芻家畜對飼料的消化程度[3]。同一真菌菌株中分離到的阿魏酸酯酶的最適溫度、最適pH值等理化性質都比較相似,但在底物特異性方面都存在較大差異,這表明它們在木質纖維素降解過程中發揮的作用是不同的,具體機制還有待進一步研究。
隨著研究的進行,在很多植物病原真菌和大型真菌中也開始有該酶的報道[4-6]。
1.1.2細菌阿魏酸酯酶產阿魏酸酯酶的細菌主要為動物和人的腸道細菌,最近在植物病原細菌和瘤胃細菌中也有報道。腸道細菌產生的阿魏酸酯酶能從膳食纖維中釋放出腸道易于吸收的游離阿魏酸,使其進入血液中發揮抗氧化的功效。這已經被證實是乳酸菌抗氧化性的主要原因之一。Coutea等對采集自人糞便排泄物的35種細菌分離物進行了阿魏酸酯酶活性的測定,最后發現6種分離物具有阿魏酸酯酶的活性,研究中作者還首次證實了腸道細菌產生的阿魏酸酯酶能將膳食纖維中的具抗氧化活性的阿魏酸釋放出來,是有益于人體健康的[7]。Abeijo′n Mukdsi等用含有產阿魏酸酯酶的發酵乳酸桿菌的羊奶奶酪喂食小鼠,發現攝食奶酪的小鼠比對照組小鼠的抗氧化能力明顯提高,表現為血漿中的硫代巴比妥酸物質降低了2倍,谷胱甘肽還原酶提高了3倍,進一步證實乳酸菌產生的阿魏酸酯酶能提高腸道內的阿魏酸等抗氧化類物質的生物利用率[8]。瘤胃微生物產生的阿魏酸酯酶對飼料中木質纖維素類物質的快速消化起著重要作用,最初只在瘤胃真菌中被發現。李家寶等在瘤胃細菌Cellulosilyticum ruminicola H1的基因組中克隆了三種性質各異的阿魏酸酯酶,分別命名為FaeⅠ、FaeⅡ和FaeⅢ,三種酶的最適反應溫度在40℃左右,最適pH分別為6.0~7.0、8.0和9.0,其中FaeⅡ對玉米芯和人工合成的阿魏酸甲酯、咖啡酸甲酯等模式底物都具有較高的催化活性,而FaeⅢ只對人工合成底物具特異性。通過分析作者推測出了其中兩種阿魏酸酯酶在木質纖維素降解中的協同作用機制:FaeⅠ首先松散天然纖維素結構,將纖維素暴露出來,隨后大分子底物被(半)纖維素酶降解成小分子酚酸——糖酯;而FaeⅡ對小分子底物具有較高親和力,可迅速降解被轉運進細胞的底物[9]。
1.1.3植物阿魏酸酯酶隨著人們對阿魏酸酯酶研究的深入和廣泛,在麥芽等發芽的谷物中也檢測到了阿魏酸酯酶活性,但其報道相對較少,而且目前還沒有關于其在植物體內具體功能的報道。Sancho等首次在大麥粒和麥芽中檢測到了阿魏酸酯酶的活性,并發現大麥在25℃發芽2 d后阿魏酸酯酶活性達到最高,因此推測該酶可能在大麥的發芽過程中起作用,Humberstone等也曾在大麥麥芽的提取物中檢測到阿魏酸酯酶的活性,但只到2010年大麥芽中的阿魏酸酯酶才被分離純化出來,分子量為29.3 kDa,最適pH和溫度分別為5.5和50℃[10]。Latha等在發芽的龍爪稷中也分離純化到了一個阿魏酸酯酶,分子量為79.4 kDa,有四個亞基,其最適pH值和最適溫度分別為7.5和45℃,且在pH 6.0~9.0和30~40℃時比較穩定[11]。
2 阿魏酸酯酶的理化特性
從1991年至今,有60多種阿魏酸酯酶已經被分離純化。其中主要為微生物阿魏酸酯酶,植物中只有大麥芽和龍爪稷的阿魏酸酯酶已經被分離純化。這些阿魏酸酯酶的理化性質差異很大,主要表現在分子量大小、最適pH值和溫度、等電點、底物特異性等方面。如分子量大小主要分布在30~70 kDa之間;最適反應pH值主要集中在5~7;最適反應溫度主要集中在40~60℃(表1)。
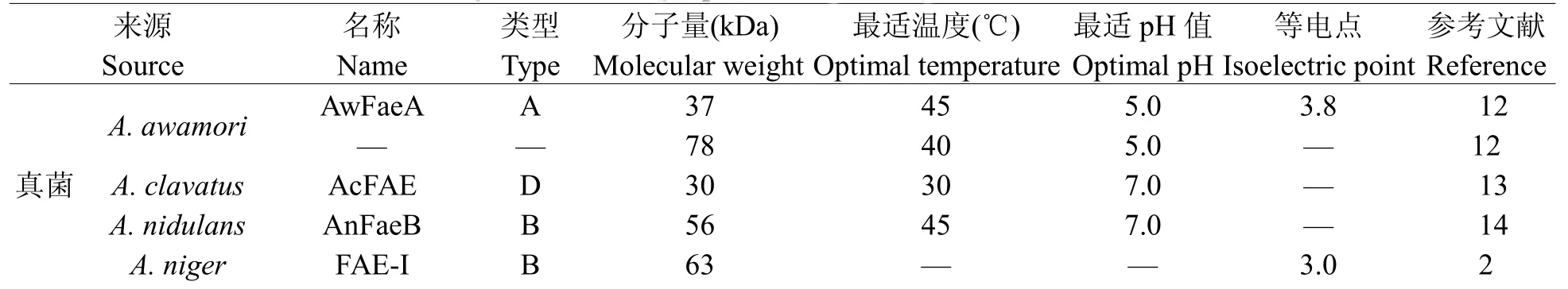
表1 部分已純化的阿魏酸酯酶的理化性質Table1Physicochem icalpropertiesof partial purified FAEs known to date
“—”:沒有相關數據.“—”:no relevantdata.
3 阿魏酸酯酶的分類
阿魏酸酯酶最初根據它們包含的芳香基團被分為兩大類。之后又根據其對四種羥基化肉桂酸甲酯模式底物阿魏酸甲酯、咖啡酸甲酯、p-香豆酸甲酯和芥子酸甲酯(圖1)的專一性、微生物生長中對底物的需求以及蛋白質序列特異性,將阿魏酸酯酶分為A、B、C、D四種類型(表2)[19]。
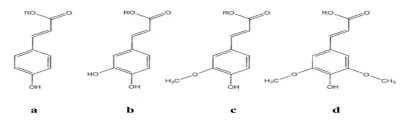
圖1 四種羥基肉桂酸的化學結構(a)p-香豆酸;(b)咖啡酸;(c)阿魏酸;(d)芥子酸。R=H時為游離酸形式;R=CH3時為它們的甲基酯形式Fig.1Chem icalstructuresof four hydroxycinnam ic acids (a)p-Coumarate;(b)Caffeate;(c)Ferulate;(d)Sinapinate.R=H-free acid form,R=CH3-methyl-esterified form

表2 基于底物特異性、是否釋放二聚阿魏酸以及序列相似性對阿魏酸酯酶進行的分類Table 2 Classification of feruloylesterases(FAEs)based on their activities toward synthetic substrates,their ability to release diferulic acids(diFerA)and their sequence sim ilarity
Shin等發現4類阿魏酸酯酶都有其特有的保守序列G-X-S-X-G。來源于塔賓曲霉(A.tubingensis)、黑曲霉(A.niger)和泡盛曲霉(A.awamori)的A型阿魏酸酯酶都具有共同的氨基酸序列G-H-S-L-G,這些氨基酸序列都存在于一個由16個氨基酸組成的保守序列中(ALTVTGHSLGASLAAL);來源于柄籃狀菌(T.stipitatus)、構巢曲霉(A.nidulans)和黑曲霉(A.niger)的C型阿魏酸酯酶都具有共同的氨基酸序列G-C-S-T-G,且其典型序列都存在于一個由13個氨基酸構成的保守序列(SYYLGCSTGGRQG)中,B型和D型阿魏酸酯酶的相應氨基酸序列不如A型和C型保守,作者推測阿魏酸酯酶在酯酶基序周圍的保守序列可能在底物特異性上起了決定作用[14]。
4 阿魏酸酯酶的結構特征和催化機理
4.1阿魏酸酯酶的結構特征
由于底物結構的復雜性,不同類型的阿魏酸酯酶在空間結構上存在著差異,但都為結構緊湊的球蛋白,整個拓撲結構為以多個α螺旋包圍多個連續的β折疊片形成的典型α/β水解酶結構,活性中心大都具有典型的催化三聯體結構“Ser-His-Asp”。如來自于黑曲霉的A型阿魏酸酯酶(FAEA或FAEA-Ⅲ,圖2a),FAE-Ⅲ中央含有9個連續的β折疊片,周圍被5個α螺旋和2個β折疊片包圍(Katherine etal.,2004);丁酸弧菌Butyrivibrio proteoclasticus的阿魏酸酯酶Est1E(圖2b),中央含有8個平行的β折疊片(其中一個反向平行),6個α螺旋分別圍繞在兩側,另外還有4個β折疊片和2 個α螺旋插入到其中[24]。
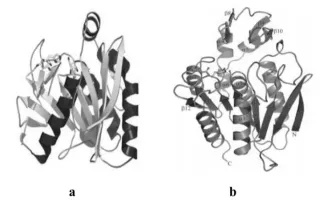
圖2 (a)黑曲霉阿魏酸酯酶FAEA-Ⅲ的三維晶體結構;(b)丁酸弧菌阿魏酸酯酶Est1E的三維晶體結構Fig.2(a)Representation of the three-dimensionalstructure of the FAEA-Ⅲ;(b)Representation of the three-dimensional structure of the Est1E
程凡升等應用生物信息學的蛋白質結構預測方法,探討了來源于荷斯坦奶牛瘤胃宏基因組文庫的一個新型阿魏酸酯酶(FAE-SH1)的結構,發現FAE-SH1有一個以G-X-S-X-G(121-127)為特征的酯酶保守區域,而且還具備α/β水解酶的典型特征:Ser123、Asp215和His244在空間構型上構成可能的催化中心。此外,編碼該酶的基因序列還存在一個Ile-His-Gly(49-51)保守區域,經分析該區域可以形成一個“氧空洞”,可能在酯鍵水解過程起穩定中間體的作用。FAE-SH1由7個α螺旋和8個平行的β折疊片組成,其中8個β折疊形成酶的主體結構,7個α螺旋分布于β折疊的周圍,所有α螺旋和β折疊的方向均是右手螺旋,三級結構建模發現該酶的三聯體催化中心Ser123、Asp215和His244可以在空間上接近并且在蛋白表面形成凹陷的裂縫,構成了供小分子進入的“入口”,這一結構是典型的酯鍵降解結構,是完成酯鍵水解過程的關鍵步驟,此外第136-176號氨基酸殘基在“裂縫入口”處形成一個類似“蓋子”的結構,該結構可阻止底物或者其它溶劑進入活性中心[25]。
4.2阿魏酸酯酶的作用機理
阿魏酸酯酶大多都含有Ser-His-Asp的“三點催化活性中心”,它的水解反應是通過共價的酰基-酶中間體的形成和隨后的分解完成的。以黑曲霉的阿魏酸酯酶FAE-Ⅲ為例,它的活性位點為Ser133,Asp194和His247,其中Ser催化殘基在保守序列G-X-S-X-G的中心,且正好處于連接蛋白質表面2個裂縫間的大腔中,完全暴露于外界溶劑,而Asp和His 2個催化殘基則位于α/β水解酶的典型位置。FAE-Ⅲ的活性部位由一個蓋狀區和一個殘基環組成,其中蓋狀區決定底物的特異性,殘基環為底物結合提供較好的調整性,而阿魏酸酯酶序列中特有的N-糖基化位點使活性部位一直處于開放構象狀態下,從而結合底物。在催化過程中,阿魏酸酯酶的催化中心和輔助中心將阿魏酸酯化合物中的阿魏酸殘基束縛,然后經過一系列化學過程將酯鍵斷裂,使阿魏酸游離出來[12]。Koseki等克隆了泡盛曲霉(A.awamori)的阿魏酸酯酶基因Aw faeA,發現它與黑曲霉阿魏酸酯酶FAE-Ⅲ的序列相似度為92%,且Ser133,Asp194和His247三個氨基酸殘基也是很保守的,由此推測Aw FAEA的活性中心也是由這三個氨基酸組成,通過對蓋狀區的四個氨基酸Asp71,Thr72,Asp77和Tyr80進行突變研究了蓋狀區的功能,進一步證明了蓋狀區結構是與底物的特異性密切相關的[16]。
5 阿魏酸酯酶基因的克隆表達與調控
5.1阿魏酸酯酶基因的克隆表達
目前已經有近20種不同來源的阿魏酸酯酶基因被克隆和測序,其中多個已經在原核和真核表達系統實現了異源或同源表達。研究發現,真菌阿魏酸酯酶在大腸桿菌(Escherichia coli)中表達時不能形成正確空間折疊的蛋白,導致其在大腸桿菌中不表達或表達量很低,如A.niger阿魏酸酯酶FAEA在大腸桿菌中的表達量僅為在黑曲霉中表達量的1/10[26]。相比而言,畢氏酵母(Pichia pastoris)是較理想的真菌阿魏酸酯酶的表達體系,以芽殖酵母的α因子為分泌信號,并利用乙醇氧化酶(AOXl)的啟動子在畢氏酵母中已經實現了多個真菌阿魏酸酯酶基因的異源表達[27]。
為了更好的發揮阿魏酸酯酶的功能,科研人員還將阿魏酸酯酶克隆到植物中使其分泌阿魏酸酯酶從而對自身細胞壁進行降解。Buanafina等將黑曲霉(A.niger)的阿魏酸酯酶基因轉入了高羊茅草(Festuca arundinacea)中,并分別利用組成型表達的水稻肌動蛋白啟動子、誘導型表達的花椰菜花葉病毒(CauliflowerMosaic Virus,CaMV)35s啟動子、玉米的泛素啟動子以及可誘導的熱休克和衰老啟動子進行了表達,結果發現幾種啟動子都可以有效的在轉基因植物體內表達阿魏酸酯酶并將其定位于植物液泡內,但可誘導的熱休克和衰老啟動子表達的效率最高,當細胞死亡時轉基因高羊茅內表達的阿魏酸酯酶可以對植物細胞壁進行“自體消化”,而且這種降解效率可隨著外源木聚糖酶的加入而大大增強,使得轉基因植物可以更好的用于動物飼料或是生物乙醇的生產。
5.2阿魏酸酯酶基因的調控
阿魏酸酯酶表達調控方面的研究較少,目前研究的比較深入的是編碼黑曲霉阿魏酸酯酶FAEA 的faeA基因。研究發現faeA的表達主要受木聚糖降解調節子XlnR、碳源代謝物抑制子CreA和對某些芳香族化合物做出反應的某個因子等三大因子的調控。faeA的誘導表達主要有2個途徑:一個為木聚糖/木糖誘導途徑,它受木聚糖降解調節子XlnR調節;另一個為芳香族化合物(如阿魏酸、香草酸、香蘭醇等)誘導途徑,適當濃度的阿魏酸能誘導阿魏酸酯酶的表達,但濃度過高時阿魏酸會對細胞產生毒害作用,反而使阿魏酸酯酶的產量下降,兩個途徑是有協同效果的,適當濃度阿魏酸的加入可以增強木聚糖對faeA的誘導表達。碳代謝物抑制子蛋白CreA介導的碳代謝產物抑制途徑對faeA的表達有抑制作用,外源加入葡萄糖能抑制以木糖為碳源的野生菌株faeA基因的表達,但對CreA突變株卻幾乎沒有抑制作用,說明碳代謝產物對faeA的抑制主要是由于CreA導致的。
6 阿魏酸酯酶的發酵生產
阿魏酸酯酶的發酵生產主要有液體發酵和固體發酵兩種,其中固體發酵以周期短、能耗少、水消耗量較低、對環境污染程度低、產量大等優點在阿魏酸酯酶的生產中受到廣泛的重視。目前在黑曲霉、青霉、嗜熱側孢霉和泡盛曲霉等微生物中都實現了固體發酵產阿魏酸酯酶。阿魏酸酯酶是誘導酶,常用麥麩、玉米麩、啤酒糟、燕麥木聚糖、甜菜渣等木質纖維素含量豐富的混合碳源作基質來進行生產,而當以葡萄糖、木糖、乳糖、麥芽糖等單糖或二糖作為碳源時,微生物不會產生阿魏酸酯酶。F.proliferatum NRRL26517在以玉米麩為碳源時產生的阿魏酸酯酶是以燕麥木聚糖為碳源時的7倍多,研究發現這是因為玉米麩中含有較多的阿魏酰酯鍵,而用去除阿魏酰酯鍵結構的玉米麩作為碳源時,該菌株產阿魏酸酯酶的能力大大下降,表明玉米麩中的阿魏酰酯鍵對誘導阿魏酸酯酶的產生是必需的[14]。Topakas等在誘導嗜熱側孢霉產阿魏酸酯酶時發現用堿處理除去麥秸中的阿魏酰酯鍵后,該菌株產阿魏酸酯酶的能力大大降低,再次證實了碳源中的阿魏酰酯鍵對誘導產生阿魏酸酯酶的重要性[2]。
7 阿魏酸酯酶的應用現狀
7.1分離阿魏酸
植物細胞壁是阿魏酸的最重要來源,糧食加工業每年產生大量副產品,如啤酒糟、麥麩、玉米麩、米糠等,這些物質的主要成分是細胞壁,含有豐富的阿魏酸。傳統工業中普遍采用堿處理的方法來提取阿魏酸,高濃度的堿易引起植物細胞壁中其他成分的改變而且還容易有化學殘留,不利于后期的提純。研究發現阿魏酸酯酶可以從麥麩、玉米麩、小麥秸稈、玉米秸稈、啤酒糟、甘蔗渣、燕麥殼、咖啡渣、蘋果渣等很多農副產品中釋放出阿魏酸[5,6,22,23]。近年來研究者越來越多地采用溫和而特異性高的酶法來提取阿魏酸并取得了很好的效果。
7.2用作膳食纖維補充劑
流行病學研究認為麥麩、燕麥等膳食纖維的消費與某些慢性病,如冠心病和某些癌,特別是與消化道有關的結腸癌的預防有著密切關系,這種對健康的有益作用常常歸功于膳食中的酚酸類物質。膳食中的酚酸類化合物主要是阿魏酸,而幾乎所有的阿魏酸都是以糖酯的形式存在,以這種復雜形式存在的阿魏酸通常認為不能被動物和人體所吸收。研究發現乳酸菌、枯草芽孢桿菌、雙歧桿菌等多種腸道菌產生的阿魏酸酯酶可以將膳食纖維中的將阿魏酸游離出來,但是這些微生物釋放的阿魏酸濃度太低,不能達到保健防護的效果,因此需要外源添加阿魏酸酯酶制劑來增強阿魏酸的釋放量,使之更好的發揮其生理活性功能。Abeijo′n Mukdsi等用含有能產生阿魏酸酯酶的發酵乳酸菌L. fermentum CRL1446的羊奶奶酪喂食小鼠,發現喂食奶酪的小鼠比對照組小鼠腸道內的阿魏酸酯酶活性提高2倍多,而且喂食奶酪的小鼠抗氧化能力明顯提高,表現為血漿中的硫代巴比妥酸物質(TBARS)降低了2倍,谷胱甘肽還原酶(GR)提高了3倍,進一步研究發現這都是因為乳酸菌產生的阿魏酸酯酶能將膳食纖維中的阿魏酸等抗氧化物質游離出來,提高其生物利用率,使其更好的發揮抗氧化活性[8]。
7.3合成酚酸化合物
阿魏酸、p-香豆酸和芥子酸等羥基肉桂酸具有抗菌、防紫外線、抗腫瘤、抗氧化等功效,因此在食品、醫藥和化妝品領域具廣泛的應用前景。但由于這些酚酸分子中的烷烴較短,含有雙鍵,導致其親水性較強,脂溶性較差,影響到其在體內的轉運和利用,因此大大限制了其在油脂類食品和化妝品中的應用。研究發現將阿魏酸以酯鍵的形式連接到糖、醇或者脂類的母體上,可以顯著改善阿魏酸的脂溶性,提高其穩定性。
阿魏酸酯酶可用于催化在水—有機溶劑混合體系或微乳液中發生的羥基肉桂酸的酯化或酯基交換反應,而且生成阿魏酸酯衍生物的酯化反應只能用阿魏酸酯酶進行催化。尖孢鐮刀菌F.oxysporum中分離的阿魏酸酯酶可以在沒有表面活性劑的微乳液中催化1-丙醇和羥基肉桂酸的酯化反應[2]。黑曲霉中的阿魏酸酯酶AnFaeA可以催化芥子酸甲酯的轉酯反應并生成1-或4-丁基芥子酸,研究發現芥子酸及其衍生物在抗衰老化妝品和防曬霜中有很好的應用前景,另外它還是增強DNA修復的重要因子[28]。
與游離的羥基肉桂酸相比,這些糖酯類衍生物具有更好的脂溶性,更強的活性,較低的毒性以及更強的適應性,在醫藥、食品、化妝品等方面有廣闊的應用前景。
7.4在造紙工業的應用
工業上要獲得高品質的紙漿,必須除去造成紙漿呈黑色的木質素,傳統的制漿法都采用氯化物去除木質素,但產生的廢水中含有大量的芳香族有毒污染物,對環境造成了嚴重的污染。而阿魏酸酯酶可以水解植物細胞壁間連接木質素和碳水化合物的阿魏酸酯鍵,所以在造紙行業中被認為是脫除木質素的一個新選擇。
Record等分別檢測了黑曲霉中的阿魏酸酯酶在單獨或添加漆酶介體系統和木聚糖酶的條件下脫除麥秸稈中木質素的效果,發現在麥秸稈紙漿漂白過程中單獨添加阿魏酸酯酶時可以去除38%的木質素,當與木聚糖酶和漆酶協同作用時,可以脫除74%的木質素,且卡伯(Kappa)值降至3.9[29]。
7.5在飼料工業上的應用
農作物秸稈是我國畜牧業中十分重要的粗飼料來源,其在反芻家畜日糧中所占的比例高達70%以上。但農作物秸稈中的阿魏酸、二聚阿魏酸等酚酸類分子以酯鍵方式與半纖維素分子間形成致密網狀交聯結構,空間上阻遏了瘤胃微生物所分泌的纖維素酶、木聚糖酶等木質纖維素降解酶對飼草中纖維素和半纖維素的有效酶解,一度成為瘤胃生物利用細胞壁類物質的瓶頸因素。
研究發現用阿魏酸酯酶預處理農作物秸稈等粗飼料,可促進瘤胃微生物對飼料細胞壁的降解,提高反芻家畜對飼料的消化程度,減少隨糞便排出的營養物質,從而提高飼料的利用效率[30]。
此外,因為阿魏酸酯酶可以加速木質纖維素的降解,因此在生物質利用和燃料乙醇的生產中發揮重要作用。
8 展望
綜上所述,阿魏酸酯酶在醫藥、食品、造紙、飼料等領域都具有重大應用前景,但目前已報道的阿魏酸酯酶普遍酶活力低,穩定性較差,一定程度上限制了其在實際生產中的應用。近年來國內外科學家對阿魏酸酯酶的研究主要側重其理化性質方面,而關于其結構特性和催化機理方面的報道較少,這無疑也成為限制高活性阿魏酸酯酶研制的一個關鍵因素。接下來,從該酶的結構和催化特性方面入手,進一步借助分子生物學的手段提高該酶的活性和穩定性將是值得深入研究的問題。
[1]Fazary AE,Ju,YH.Feruloyl esterases as biotechnological tools:currentand future perspectives[J].Acta Biochim ica et Biophysica Sinica,2007,39:811-828
[2]Topakas E,Vafiadi C,Christakopoulos P.M icrobial production,characterization and applications of feruloyl esterases[J].Process Biochem istry,2007,42:497-509
[3]Borneman WS,Ljungdahl LG,Hartley,etal.Purification and partial characterization of two feruloylesterases from the anaerobic Neocallimastix strain MC-2[J].Applied and EnvironmentalM icrobiology,1992,58:3762-3766
[4]Hassan S,Hugouvieux-Cotte-Pattat N.Identification of two feruloyl esterases in Dickeya dadantii3937 and induction of themajor feruloylesterase and of pectate lyasesby ferulic acid[J].Journalof Bacteriology,2011,193:963-970
[5]Wang L,Ma ZQ,Du F,etal.A novel feruloylesterase from theediblemushroom,Panusgiganteus:A potential dietary supplement[J].Journalof Agriculturaland Food chem istry,2014,62:7822-7827
[6]Wang L,Zhang R,Ma ZQ,et al.A feruloy esterase(FAE)characterized by relatively high thermostability from the ediblemushroom Russula virescens[J].Applied Biochemistry and Biotechnology,2014,172:993-1003
[7]Couteau DM,Gibson GR,Williamson G,et al.Isolation and characterization of human colonic bacteria able to hydrolyse chlorogenic acid[J].Journalof Applied M icrobiology,2001,90:873-881
[8]Abeijón Mukdsi MC,Haro C,González SN,et al.Functional goatmilk cheese w ith feruloyl esterase activity[J].Journalof Functional Foods,2013,5:801-809
[9]Li J,Cai S,Luo Y,et al.Three feruloyl esterases in Cellulosilyticum ruminicola H1 act synergistically to hydrolyze esterified polysaccharides[J].Applied and EnvironmentalM icrobiology,2011,77(17):6141-6147
[10]劉莎莎,李華,石慶華,等.大麥芽阿魏酸酯酶的分離純化及其部分酶學性質的測定[J].中國畜牧獸醫,2010,37:28-31
[11]Latha GM,Srinivas P,Muralikrishna G.Purification and characterization of ferulic acid esterase from malted finger millet[J].Journalof Agriculturaland Food chemistry,2007,55:9704-9712
[12]Kanauchi M.Characteristics and role of feruloyl esterase from Aspergillus Awamori in Japanese spirits,‘Awamori' production[J].Scientific,Health and SocialAspectsof the Food Industry,2012:145-162
[13]Damasio AR,Braga CM,Brenelli LB,etal.Biomass-to-bio-products application of feruloyl esterase from Aspergillus clavatus[J].Applied M icrobiology and Biotechnology,2013,97:6759-6767
[14]Shin HD,Chen RR.A type B feruloyl esterase from Aspergillus nidulans w ith broad pH applicability[J].Applied M icrobiology and Biotechnology,2007,73:1323-1330
[15]Hegde S,Muralikrishna G.Isolation and partial characterization of alkaline feruloyl esterases from Aspergillus niger CFR 1105 grown onwheatbran[J].world JournalofM icrobiology&Biotechnology,2009,25:1963-1969
[16]Koseki T,Hori A,Seki S,et al.Characterization of two distinct feruloyl esterases,AoFaeB and AoFaeC,from Aspergillus oryzae[J].Applied M icrobiology and Biotechnology,2009,83:689-696
[17]Yang SQ,Tang L,Yan QJ,etal.Biochem ical characteristics and gene cloning of a novel thermostable feruloyl esterase from Chaetomium sp.[J].Journalof Molecular Catalysis B-Enzymatic,2013,97:328-336
[18]Kuhnel S,Pouvreau L,Appeldoorn MM,et al.The ferulic acid esterases of Chrysosporium lucknowense C1:purification,characterization and their potential application in biorefinery[J].Enzyme and M icrobial Technology,2012,50:77-85
[19]Crepin VF,Faulds CB,Connerton IF.Identification of a type-D feruloyl esterase from Neurospora crassa[J].Applied M icrobiology and Biotechnology,2004,63:567-570
[20]McClendon SD,Shin HD,Chen RR.Novel bacterial ferulic acid esterase from Cellvibrio japonicus and itsapplication in ferulic acid release and xylan hydrolysis[J].Biotechnology Letters,2011,33:47-54
[21]Esteban-Torres M,Reveron I,Mancheno JM,et al.Characterization of a feruloyl esterase from Lactobacillus plantarum[J].Applied and EnvironmentalM icrobiology,2013,79:5130-5136
[22]Huang YC,Chen YF,Chen CY,et al.Production of ferulic acid from lignocellulolytic agricultural biomass by Thermobifida fusca thermostable esterase produced in Yarrowia lipolytica transformant[J].Bioresource Technology,2011,102:8117-8122
[23]Rakotoarivonina H,Hermant B,Chabbert B,et al.thermostable feruloyl-esterase from the hem icellulolytic bacterium Thermobacillus xylanilyticus releases phenolic acids from non-pretreated plant cellwalls[J].Applied M icrobiology and Biotechnology,2011,90:541-552
[24]Goldstone DC,Villas-Boas SG,Till M,et al.Structural and functional characterization of a prom iscuous feruloyl esterase(Est1E)from the rumen bacterium Butyrivibrio proteoclasticus[J].Proteins,2010,78:1457-1469
[25]程凡升,張茂秋,程凡杰,等.結構預測及分子對接方法研究幾種水解產物對新阿魏酸酯酶的抑制作用[J].高等學校化學學報,2012,33:1788-1793
[26]Benoit I,Navarro D,Marnet N,et al.Feruloyl esterases as a tool for the release of phenolic compounds from agro-industrialby-products[J].Carbohydrate Research,2006,341:1820-1827
[27]Zhang SB,Wang L,Liu Y,et al.Expression of feruloyl esterase A from Aspergillus terreus and its application in biomass degradation[J].Protein Expression and Purification,2015,115:153-157
[28]Vafiadi C,Topakas E,Alissandratos A,et al.Enzymatic synthesis of butyl hydroxycinnamates and their inhibitory effectson LDL-oxidation[J].Journalof Bacteriology,2005,133:497-504
[29]Record E,Asther M,Sigoillot C,et al.Overproduction of the Aspergillus niger feruloyl esterase for pulp bleaching application[J].Applied M icrobiology and Biotechnology,2003,62:349-355
[30]楊紅建,黎大洪,謝春元,等.阿魏酸酯酶處理對羊草、玉米秸、稻秸及麥秸瘤胃體外發酵特性的影響[J].動物營養學報,2010,22:207-211
A Review on Research and Applications of Feruloyl Esterase
WANG Li1,2,SUNQin-dong1,WANGHe-xiang2*
1.College ofPlantProtection/Shandong AgriculturalUniversity,Taian 271018,China
2.College ofBiological Sciences/China AgriculturalUniversity,Beijing 100193,China
Feruloyl esterases(FAEs)are able to hydrolyze the ester bond between hydroxycinnam ic acids and hem icellulosic sugars and release ferulic acid from the plant cellwall,which demonstrate awide variety of applications in food,cosmetic,medical,feed industries and producing bioethanol.This review provided an overview of the current know ledge on feruloyl esterases including its source,physicochem icalproperty,structural featuresand catalyticmechanism and itsapplications.
Feruloylesterase;research progress;application progress
Q55
A
1000-2324(2016)04-0628-08
2016-05-18
2016-05-24
山東省自然科學基金(ZR2015CL040);山東農業大學校青年科技創新基金(24031)
王麗(1985-),女,山東聊城人,博士,主要從事大型真菌中生物活性物質研究.E-mail:haoyou0102@163.com
Author for correspondence.E-mail:hxwang@cau.edu.cn
