乳酸菌中乳酸脫氫酶的研究進展
2016-09-14 12:25:38黃艷娜許光濤游春蘋乳業生物技術國家重點實驗室光明乳業股份有限公司乳業研究院上海00436邯鄲鋼鐵集團職工醫院河北邯鄲05600
食品工業科技 2016年8期
關鍵詞:研究
黃艷娜,許光濤,游春蘋,*(.乳業生物技術國家重點實驗室,光明乳業股份有限公司乳業研究院,上海00436;.邯鄲鋼鐵集團職工醫院,河北邯鄲05600)
乳酸菌中乳酸脫氫酶的研究進展
黃艷娜1,許光濤2,游春蘋1,*
(1.乳業生物技術國家重點實驗室,光明乳業股份有限公司乳業研究院,上海200436;2.邯鄲鋼鐵集團職工醫院,河北邯鄲056001)
乳酸脫氫酶(Lactate dehydrogenases,LDHs)是乳酸菌利用碳源轉化丙酮酸為乳酸的關鍵酶。本文從乳酸脫氫酶的分類、結構與功能、合成乳酸的微生物屬別及代謝途徑等方面,綜述了乳酸菌中乳酸脫氫酶的研究進展,以期為微生物合成乳酸的代謝調控提供一定的理論指導。
乳酸菌,L-乳酸脫氫酶,D-乳酸脫氫酶,分類,結構與功能
乳酸是一種重要的生物基平臺化合物,廣泛應用于農業、食品、醫藥、化工和環保等領域。根據其旋光性的不同可分為D-乳酸和L-乳酸,分別由D-乳酸脫氫酶(D-LDH,EC:1.1.1.28)和L-乳酸脫氫酶(LLDH,EC:1.1.1.27)催化[1-3]。近年來,隨著人們對利用L-乳酸聚合生成聚L-乳酸(PLA)以合成生物降解塑料的重視,乳酸菌也隨之得到了廣泛關注[4]。
乳酸菌是一類發酵糖類合成乳酸的革蘭氏陽性細菌的總稱,廣泛存在于動物、植物和乳制品中。乳酸脫氫酶(Lactate dehydrogenases,LDHs)作為乳酸菌合成乳酸的關鍵酶,目前已研究的較為透徹。盡管該酶分布十分廣泛,但不同的LDHs性質存在較大的差異。因此,本文將主要圍繞乳酸脫氫酶的分類、種群分布、結構與功能以及乳酸合成的代謝調控等方面,綜述乳酸菌中乳酸脫氫酶的研究進展,以解析其在乳酸合成中的作用機制。
1 乳酸脫氫酶的分類
根據結合輔酶的不同,微生物體一般包含兩種乳酸脫氫酶,NAD-依賴型乳酸脫氫酶(NAD-dependent lactate dehydrogenases,nLDHs)和NAD-非依賴型乳酸脫氫酶(NAD-independent lactate dehydrogenases,iLDHs)兩大類。iLDHs,也被稱作呼吸鏈乳酸脫氫酶,在微生物體內通常被認為是負責乳酸氧化的酶類。它們編碼的基因與乳酸利用相關的乳酸透性酶和調節蛋白編碼的基因較近[5]。相較于iLDHs,nLDHs分布范圍更廣泛,在人體、動物以及微生物中都普遍存在。在微生物中,nLDHs存在于其細胞質中,體外實驗證實:nLDHs能夠以NADH為輔因子催化丙酮酸還原生成乳酸,并且氧化NADH為NAD+,同時該酶在特定的條件下可以催化逆反應的進行,但是活力非常弱,不容易檢測。一般說來,與nLDHs以NADH為輔因子不同的是iLDHs主要利用黃素[6-7]。nLDHs也可以在
體內催化乳酸的氧化,但體外實驗可能需要較高的pH和底物濃度[8]。
按其催化底物的構型不同,nLDHs可以分為NAD依賴型-L-乳酸脫氫酶(L-nLDHs)和NAD依賴型-D-乳酸脫氫酶(D-nLDHs)兩大類,分別催化丙酮酸合成L-乳酸和D-乳酸[6]。研究表明,雖然L-nLDHs和D-nLDHs均催化丙酮酸合成乳酸,但二者的基因序列不具有相似性或一致性。已知的不同乳桿菌的D-nLDHs之間存在40%~50%的氨基酸同源性,而D-nLDHs和L-nLDHs之間則只存在10%左右的同源性,表明兩種脫氫酶分別由兩個完全不相關的家族進化而來。不僅如此,酶之間的差異也表現在動力學參數差異明顯,比如,兩者催化反應的反應常數kcat和米氏常數Km不同。
根據天然電子受體的不同,可以將iLDHs分為三類。第一類為膜蛋白,利用膜醌類作為外部的電子受體[7,9];第二類直接利用O2作為電子受體,根據氧化終產物的不同,又將其細分為乳酸氧化酶(Lactate oxidase,LOX)和乳酸單氧酶(Lactate monooxygenases,LMO),其中前者產生丙酮酸和H2O2,而后者產生乙酸、CO2和H2O[10-11];第三類是硫胺b2(flavocytochrome b2),存在于真菌中,它天然的電子受體為細胞色素c。大多數L-iLDHs屬于α-羥酸氧化黃素蛋白家族,利用還原型黃素單核苷酸(Flavin mononuleotide,FMN),而所有的D-iLDHs均可利用黃素腺嘌呤二核苷酸(FAD)作為電子受體,而不是FMN,它們具有FAD結合的氧化還原酶/轉移酶4的家族特征[12-13]。此外,D-iLDHs還可以利用醌類或細胞色素c作為電子受體,但是迄今為止,還沒有D-iLDHs直接利用O2的相關報道。Wolin[14]的研究表明,一些鏈球菌LDHs的催化活性絕對依賴于果糖-1,6-二磷酸(Fructose-1,6-Diphosphate,FDP)。其他鏈球菌、雙歧桿菌屬的所有種、干酪乳桿菌和木糖乳桿菌等都發現了相似的iLDHs。
2 乳酸脫氫酶的結構與功能
伴隨著全基因組測序技術的發展,科學家已發現很多乳酸菌中存在多個乳酸脫氫酶基因[15],這些基因表達的乳酸脫氫酶由于基因序列和氨基酸序列的不同,在酶學性質上有很大的差異,甚至在乳酸菌中的功能都有所差異[16]。研究表明,L-LDH和L-蘋果酸脫氫酶屬于同一家族,而D-LDH屬于D-2-羥酸脫氫酶家族[8]。
2.1 D-乳酸脫氫酶(D-LDH)的結構和功能
大多數D-LDH催化是可逆反應,極少數不可逆;對于可逆的D-LDH來說,只有環境中乳酸濃度較高時才催化逆反應,即催化乳酸合成丙酮酸,來參與細菌的代謝。
D-LDH的一級結構比對表明,不同種屬的D-LDH的氨基酸序列存在較大的差異,但是參與丙酮酸的結合與催化的氨基酸殘基卻十分保守。Kochhar等[17]的研究顯示L.bulgaricus的D-LDH是一個由相同亞基組成的二聚體,每個亞基由332個氨基酸殘基組成,分子量為36.8 ku,經化學修飾發現位于催化中心的三個組氨酸(His205、His296和His303)和Asp259對底物的催化具有十分重要的作用。Kochhar等還發現,His289也參與了底物結合,但His289不是D-2-羥基酸脫氫酶家族中保守的氨基酸殘基。另外,通過對Cys殘基的化學修飾發現Cys234可能參與底物結合和催化作用,這在L.helveticus的羥基丙酮酸還原酶中也能得以證實[17]。研究還發現L.helveticus CNRZ32 的D-LDH氨基酸序列和D-2-羥基酸脫氫酶家族的D-3-磷酸甘油酸脫氫酶、羥基丙酮酸還原酶和D-2-羥基己酸脫氫酶存在很高的同源性,這也暗示了該酶是D-2-羥基酸脫氫酶家族的一個成員[18]。Razeto等[19-20]對L.bulgaricus的D-LDH構象進行了研究,結果發現L.bulgaricus的D-LDH是一個由兩個相同的亞基構成的不對稱酶,A亞基的酶蛋白是典型的“開放”構象,而B亞基是典型的“閉合”構象。其中NADH的結合位點主要存在于B亞基中,而在A亞基中僅有30%,同時在底物結合口袋中有一個硫酸根離子。另外,該研究還建立了丙酮酸分子在活性位點的模型。在閉合域中,存在一簇疏水的氨基酸殘基緊緊圍繞著丙酮酸分子的甲基,在這個疏水氨基酸簇中至少有三個氨基酸殘基(Tyr52、Phe299和Trp135’)決定底物的專一性。研究表明該底物結合位點有利于閉合域的穩定和酶的激活。
2.2 L-乳酸脫氫酶(L-LDH)的結構與功能
大多數乳酸菌中不僅存在D-LDH也存在L-LDH,L-LDH催化丙酮酸還原生成L-乳酸。L-LDH分為兩型:一類可被FDP激活,屬于別構酶;另一類不需要FDP激活,不具有別構效應。據Hiroyuki Uchikoba報道,L.pentosus的L-LDH是一個非異構酶,但是它的氨基酸序列和一些細菌的別構LDH具有很高的相似性[21]。該酶的四聚體由相同亞基組成對稱的酶結構。它的活性構象和別構LDH的構象相似。
Kazuhito Arai等[22]的研究表明,Lactobacillus的L-LDH氨基酸序列中,Glu102、Asp197和Thr246是高度保守的,它們參與了底物的識別;另外,98-110氨基酸殘基構成了活性位點環,它們參與催化反應。Arg171的胍基和丙酮酸的羰基形成雙氫鍵,從而使丙酮酸在催化位點中處于正確的結合方向[23]。研究表明,在L-LDH的氨基酸序列中同樣存在一個與輔酶NADH結合的結構域Gly-Xaa-Gly-Xaa-Xaa-Gly-(17Xaa)-Asp,其中該結構域中的Asp決定了L-LDH的輔酶是NADH,而不是NADPH[24]。L.pentosus中L-LDH的活性位點和其他LDH相似,包括了參與催化與底物結合的保守氨基酸殘基,如Aspl68、Argl71和Hisl95[25]。其中,Argl71在底物的結合中起著重要作用;另外,Hisl95主要在催化過程中作為質子供體和受體[26]。值得注意的是,雖然Bacillus 和Lactobacillus的L-LDH在進化關系上存在很近的親緣關系,但是活性位點環的氨基酸殘基也存在差異,如101位Bacillus是Asn,而Lactobacillus是Pro。這表明L-LDH是否來自好氧或厭氧的革蘭氏陽性菌可以由這個位置的氨基酸殘基是Asn101還是Pro101來識別[27]。
3 合成乳酸的微生物屬別
3.1 合成L-乳酸的微生物屬別
產光學純的L-乳酸的乳酸菌主要有以下四個屬中的四十余種[24,28-29],包括乳桿菌屬(Lactobacillus)、鏈球菌屬(Streptococcus)、明串珠菌屬(Leuconostoc)和片球菌屬(Pediococcus),如L.casei、L.paracasei、L.mali、L.alimentarius、Lactococcus lactis、Streptococcus thermophilus和Pediococcus pentosaceus等均合成L-乳酸,也有利用嗜熱脂肪芽孢桿菌(Bacillus stearothermophilus)和 凝 結 芽 孢 桿 菌(Bacillus coagulans)生產L-乳酸的報道。
3.2 合成D-乳酸的微生物屬別
產光學純的D-乳酸的乳酸細菌主要分布在乳桿菌屬(Lactobacillus)[24,30]、芽孢桿菌屬(Bacillus)、芽孢乳桿菌屬(Sporolactobacillus)和明串珠菌屬(Leuconostoc)4個屬[31-32]。除此以外,還分布在肉桿菌屬(Carnobacterium)[33]、腸球菌屬(Enterococcus)[34]、乳球菌屬(Lactococcus)[35]、片球菌屬(Pediococcus)[36]、鏈球菌屬(Streptococcus)[37]、四聯球菌屬(Tetragenococcus)[38]、漫游球菌屬(Vagococcus)和魏斯氏菌屬(Weissella)等中[32]。目前國內外研究較多的D-乳酸生產菌主要集中在乳桿菌屬和芽孢乳桿菌屬,這兩類菌都是同型發酵菌,以葡萄糖為碳源,通過糖酵解途徑生成丙酮酸,然后丙酮酸經D-乳酸脫氫酶作用生成D-乳酸。
4 D/L-乳酸的合成
乳酸是由丙酮酸還原而成,由于菌體內酶系統的差異,其代謝途徑分為三類,即同型發酵(Homofermentation)途徑、異型發酵(Heterofermentation)途徑和雙歧(Bifidum Pathway)途徑,其中以前兩種為主(圖1)。工業生產乳酸的菌種為同型乳酸發酵類型,其理論轉化率為100%,發酵產物中乳酸占80%,其余為乙酸等[39]。
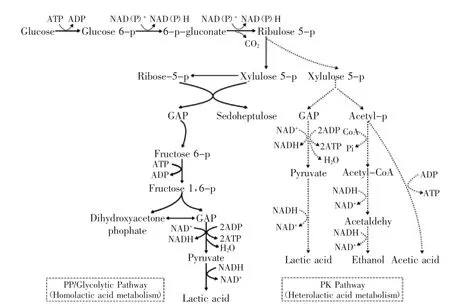
圖1 乳酸菌利用葡萄糖合成乳酸的代謝途徑[2]Fig.1 Metabolic pathways for lactic acid production from glucose by the lactic acid bacteria[2]
乳酸菌細胞內乳酸脫氫酶的立體特異性決定了形成乳酸的立體構型,如L.delbrueckii、L.bulgaricus、L.jensenii和L.coryniforis發酵產物以D-乳酸為主[11-12,31,40],有的菌株含有兩種乳酸脫氫酶或存在乳酸消旋酶,最終形成DL-乳酸。而有些乳酸菌產生的是L-乳酸,當L-乳酸積累時,則誘導產生乳酸消旋酶,該酶將L-乳酸轉變為D-乳酸,直至平衡。有研究顯示,改變pH或營養物質,D-乳酸或L-乳酸的比例將發生變化。
光學純的同分異構體可以通過表達光學特異活性的D或L-乳酸脫氫酶獲得。對于合成D-乳酸的乳酸菌來說,D-LDHs具有較L-LDHs更高的丙酮酸催化效率。采用D-LDHs或者L-LDHs生物轉化丙酮酸可產生有光學特性的D-乳酸或者L-乳酸。通過過量表達ldhD或ldhL基因,在合適的宿主中高效表達來實現相應構型乳酸產量的提高。楊登峰等[41]在大腸桿菌中過量表達L.rhamnosus的ldhL基因,成功生產獲得純度為99%的L-乳酸;Zhou等通過將乳酸片球菌Pediococcus acidilactici的L-LDH基因ldhL替換大腸桿菌中的D-LDH基因ldhA,從而得到的工程菌SZ85可生產產率為95%的高光學純的L-乳酸[42]。Okano等敲除L.plantarum NCIMB 8826的ldhL基因,表達來自牛鏈球菌Streptococcus bovis148的α-淀粉酶基因,得到的工程菌可有效利用淀粉合成高光學純的D-乳酸[43]。
乳酸的合成受到發酵溫度的影響,這與乳酸脫氫酶受到溫度的調控有關。Doo等[44]篩選到一株D-乳酸的高產菌株L.coryniformis subsp.torquens KCTC 3535,其合成D-乳酸的濃度可高達131.7 g/L,提高發酵溫度,L-乳酸的合成迅速增加。隨后他們開展了溫度相關的乳酸脫氫酶的研究,研究顯示L.coryniformis在培養溫度為30℃時發酵產物以合成D-乳酸為主,發酵溫度提高到40℃可合成大量的L-乳酸,這表明L-LDHs較高的熱穩定性可能是L.coryniformis在高溫條件下合成光學純D-乳酸產率降低的關鍵因素[40]。
較高的底物和乳酸濃度會影響細胞的滲透壓,從而抑制細胞的生長和乳酸的合成。Zou等在干酪乳桿菌L.casei搖瓶發酵過程中添加2 g/L甜菜堿代替Tween 80,可提高L-乳酸脫氫酶的活性,對該菌進行補料分批發酵,L-乳酸的濃度提高到190 g/L[45]。
乳酸的合成還受到氧的調控。Collins和Lascelles的研究顯示,厭氧條件下LDHs的酶活性是好氧條件的10倍左右[46],因此不同的發酵條件,如通氣量和攪拌轉速等的不同也會影響乳酸的合成。除此以外,還可以通過代謝調控的手段控制乳酸的合成。有研究表明,在鏈球菌中,FDP通過影響別構酶L-LDH的活力,可改變發酵代謝途徑的方向和通量:在細胞內FDP充足時,發酵產物主要是乳酸;而細胞內FDP缺乏時,發酵產物主要是乙酸、甲酸和乙醇[47]。適當調節葡萄糖碳源流向糖酵解途徑和磷酸戊糖途徑,使之積累更多的FDP,可達到積累乳酸的目的。所以如何調控胞內FDP濃度、能量水平、還原力以及幾種酶激活劑濃度,是研究的下一個目標。
5 展望
乳酸脫氫酶是一類分布非常廣泛的脫氫酶類,該酶是生物法合成乳酸的關鍵酶,尤其在乳酸菌中。
本文從乳酸脫氫酶的分類、結構和功能以及合成乳酸的種屬分布等方面,綜述了目前D-乳酸脫氫酶和L-乳酸脫氫酶在乳酸菌中的研究現狀。鑒于其在合成乳酸中的關鍵作用以及其清楚的蛋白結構和功能,可對該酶進行理性改造和定向進化,不僅可以拓寬其底物范圍、提高其酶活力,還可以賦予乳酸脫氫酶其他重要的生物功能。
[1]Abdel-Rahman M A,Tashiro Y,Sonomoto K.Lactic acid production from lignocelluloses derived sugars using lactic acid bacteria:overview and limits[J].J Biotechnol,2011,156:286-301.
[2]Abdel-Rahman M A,Tashiro Y,Sonomoto K.Recent advances in lactic acid production by microbial fermentation processes[J].Biotechnol Adv,2013,31:877-902.
[3]Gasser F.Electrophoretic characterization of lactic dehydrogenases in the genus Lactobacillus[J].J Gene Microbiol,1970,62:223-239.
[4]Kinnersley A M,Scott T C,Yopp J H.Promotion of plant growth by polymers of lactic acid[J].Plant Growth Regul,1990,9:137-146.
[5]Zhao R,Zheng S,Duan C C,et al.NAD-dependent lactate dehydrogenase catalyses the first step in respiratory utilization of lactate by Lactococcus lactis[J].FEBS Open Bio,2013,3:379-386.
[6]Jiang T Y,Gao C,Ma C Q,et al.Microbial lactate utilization:enzymes,pathogenesis,and regulation[J].Trends in Microbiol,2014,22:589-599.
[7]Garvie E I.Bacterial lactate dehydrogenases[J].Microbiol Rev,1980,44:106-139.
[8]Goffin P,Lorquet F,Kleerebezem M,et al.Major role of NAD-dependent lactate dehydrogenases in aerobiclactate utilization in Lactobacillus plantarum during early stationary phase[J].J Bacteriol,2004,186:6661-6666.
[9]Stansen C,Uy D,Delaunay S,et al.Characterization of a Corynebacterium glutamicum lactate utilization operon induced during temperature triggered glutamate production[J].Appl Environ Microbiol,2005,71:5920-5928.
[10]Furuichi M,Suzuki N,Dhakshnamoorhty B,et al.X-ray structures of Aerococcus viridans lactate oxidase and its complex with D-lactate at pH4.5 show an alpha-hydroxyacid oxidation mechanism[J].J Mol Biol,2008,378:436-446.
[11]Maeda-Yorita K,Aki K,Sagai H,et al.L-Lactate oxidase and L-lactate monooxygenase:mechanistic variations on a common structural theme[J].Biochimie,1995,77:631-642.
[12]Cristescu M E,Egbosimba E E.Evolutionary history of D-lactate dehydrogenases:a phylogenomic perspective on functional diversity in the FAD binding oxidoreductase/transferase type 4 family[J].J Mol Evol,2009,69:276-287.
[13]Fraaije M W,van Berkel W J,Benen J A,et al.Anovel oxidoreductase family sharing a conserved FAD-binding domain [J].Trends Biochem Sci,1998,23:206-207.
[14]Wolin M J.Fructose 1,6-diphosphate requirement of streptococcal lactic dehydrogenases[J].Science,1964,146:775-777.
[15]Cristescu M E,Innes D J,Stillman J H,et al.D-and L-lactate dehydrogenases during invertebrate evolution[J].BMC Evol Biol,2008,8:268.
[16]Denni D,Kaplan N O.D-and L-lactic acid dehydrogenases in Lactobacillus plantarum[J].J Biol Chem,1960,235:810-818.
[17]Kochhar S,Hunziker P E,Leong-Morgenthaler P,et al.Evolutionary relationship ofNAD+-dependent D-lactate dehydrogenase:comparison of primary structure of 2-hydroxy acid dehydrogenases[J].Biochem Biophy Res Commun,1992,184:60-66.
[18]Arthur M,Molinas C,Dutka-Malen S,et al.Structural relationship between the vancomycin resistance protein VanH and 2-hydroxycarboxylic acid dehydrogenases[J].Gene,1991,103:133-134.
[19]Razeto A,Kochhar S,Hottinger H,et al.Domain closure,substrate specificity and catalysis of D-lactate dehydrogenase from Lactobacillus bulgaricus[J].J Mol Biol,2002,318:109-119.
[20]Holton S J,Anandhakrishnan M,Geerlof A,et al.Structural characterization ofa D-isomer specific 2-hydroxyacid dehydrogenase from Lactobacillus delbrueckii ssp.bulgaricus[J].J Struct Biol,2013,181:179-184.
[21]Uchikoba H,Fushinobu S,Wakagi T,et al.Crystal structures of non-allosteric L-lactate dehydrogenase from Lactobacillus pentosus at 2.3 A resolution:specific interactions at subunit interfaces[J].Protein,2002,46:206-214.
[22]Arai K,Kamata T,Ucbikoba H,et al.Some Lactobacillus L-lactate dehydrogenases exhibit comparable catalytic activities for pyruvate and oxaloacetate[J].J Bacteriol,2001,183:397-400.
[23]Hart K W,Clarke A R,Wigley D B,et al.A strong carboxylatearginine interaction is important in substrate orientation and recognition in lactate dehydrogenase[J].Biochem Biophys Acta,1998,914:294-298.
[24]Bernard N,Ferain T,Garmyn D,et al.Cloning of the D-lactate dehydrogenase gene from Lactobcillus delbrueckii subsp.bulgaricus by complementation in Escherichia coli[J].FEBS Lett,1991,290:61-64.
[25]Taguchi H,Ohta T.D-Lactate dehydrogenase is a member of the D-isomer-specific 2-hydroxyacid dehydrogenase family [J].Biol Chem,1993,268:l8030-18034.
[26]Hart K W,Clarke A R,Wigley D B,et al.The importance of arginine 171 in substrate binding by Bacillus stearothermophilus lactate dehydrogenase isozymes[J].Biochem Biophys Res Comm,1987,146:346-353.
[27]Griffin H G,Swindell S R,Gasson M J.Cloning and sequence analysis of the gene encoding L-lactate dehydrogenase from Lactococcus lactis:evolutionary relationships between 21 different LDH enzymes[J].Gene,1992,122:193-197.
[28]Laopaiboon P,Thani A,Leelavatcharamas V,et al.Acid hydrolysis of sugarcane bagasse for lactic acid production[J].Bioresour Technol,2010,101:1036-1043.
[29]Wessels S,Axelsson L,Hansen B E,et al.The lactic acid bacteria,the food chain,and their regulation[J].Trends Food Sci Tech,2004,15:498-505.
[30]Lapierre L,Germond J E,Ott A,et al.D-Lactate dehydrogenase gene(ldhD)inactivation and resulting metabolic effects in the Lactobacillus johnsonii strains La1 and N312[J].Appl Environ Microbiol,1999,65:4002-4007.
[31]Savijoki K,Palva A.Molecular genetics characterization of theL-lactatedehydrogenasegene(ldhL)of Lactobacillus helveticus and biochemical characterization of the enzyme[J].Appl Environ Microbiol,1997,63:2850-2856.
[32]Li L,Eom H J,Park J M,et al.Characterization of the major dehydrogenase related to D-lactic acid synthesis in Leuconostoc mesenteroides subsp.mesenteroides ATCC 8293[J].Enzyme Microb Technol,2012,51:274-279.
[33]Leisner J J,Hansen M A,LarsenM H,et al.The genome sequence of the lactic acid bacterium,Carnobacterium maltaromaticum ATCC 35586 encodes potential virulence factors [J].Int J Food Microbiol,2011,152:107-115.
[34]Shibata K,Flores D M,Kobayashi G,et al.Direct L-lactic acid fermentation with sago starch by a novel amylolytic lactic acid bacterium,Enterococcusfaecium[J].Enzyme Microbiol Technol,2007,41:149-155.
[35]Wu C D,Zhang J,Du G C,et al.Heterologous expression of Lactobacillus casei RecO improved the multiple-stress tolerance and lactic acid production in Lactococcus lactis NZ9000 during salt stress[J].Bioresour Technol,2013,143:238-241.
[36]Zhao K,Qiao Q G,Chu D Q,et al.Simultaneous saccharification and high titer lactic acid fermentation of corn stover using a newly isolated lactic acid bacterium Pediococcus acidilactici DQ2[J].Bioresour Technol,2013,135:481-489.
[37]Gezginc Y,Topcal F,Comertpay S,et al.Quantitative analysis of the lactic acid and acetaldehyde produced by Streptococcus thermophilus and Lactobacillus bulgaricus strains isolated from traditional Turkish yogurts using HPLC[J].Bioresour Technol,2013,135:481-489.
[38]Masuda S,Yamaguchi H,Kurokawa T,et al.Immunomodulatory effect of halophilic lactic acid bacterium Tetragenococcus halophilus Th221 from soy sauce moromi grown in high-salt medium[J].Int J Food Microbiol,2008,121:245-252.
[39]楊潔彬.乳酸菌-生物學基礎及應用[M].北京:中國輕工業出版社,1996:203-204.
[40]Gu SA,Jun C,Joo J C,et al.Higher thermostability of L-lactate dehydrogenases is a key factor indecreasing the optical purity of D-lactic acid produced from Lactobacillus coryniformis [J].Enzyme Microb Tech,2014,58-59:29-35.
[41]楊登峰,潘麗霞,關妮,等.產高純度L-乳酸大腸桿菌基因工程菌的初步研究[J].現代食品科技,2010,26:126-128.
[42]Zhou S,Shanmugam K T,Ingram L O.Functional replacement of the Escherichia coli D-(-)-lactate dehydrogenase gene (ldhA)with the L-(+)-lactate dehydrogenase gene(ldhL)from Pediococcus acidilactici[J].Appl Environ Microbiol,2003,69:2237-2244.
[43]Okano K,Zhang Q,Shinkawa S,et al.Efficient production of optically pure D-Lactic acid from raw corn starch by using a genetically modified L-Lactate dehydrogenase gene-deficient and α-amylase-secreting Lactobacillus plantarum strain[J].App Environ Microb,2009,75:462-467.
[44]Doo E H,Kim H H,Oh J Y,et al.Mutants of Lactobacillus coryniformis producing d-lactic acid with high concentration,high yield,and high purity and their uses(for production of d-lactic acid)[J].Republic of Korea Patent,2013,KR2013001780.
[45]Zou H B,Wu Z Q,Xian M,et al.Not only osmoprotectant:Betaine increased lactate dehydrogenase activity andL-lactate production in lactobacilli[J].Bioresour Technol,2013,148:591-595.
[46]Collins F M,Lascelles J.The effect of growth conditions on oxidative and dehydrogenase activity in Staphylococcus aureus [J].J Gen Microbiol,1962,29:531-535.
[47]Thomas T D,Turner K W,Crow V L.Galactose fermentation by Streptococcus lactis and Streptococcus cremoris:pathways,products,and regulation[J].J Bacteriol,1980,144:672-682.
The research progress of the lactate dehydrogenases in lactic acid bacteria
HUANG Yan-na1,XU Guang-tao2,YOU Chun-ping1,*
(1.State Key Laboratory of Dairy Biotechnology,Dairy Research Institute,Bright Dairy&Food Co.,Ltd.,Shanghai 200436,China;2.Handan Steel&Iron Group Staff Hospital,Handan 056001,China)
The lactate dehydrogenase was a key enzyme which converts pyruvic acid to lactic acid in lactic acid bacteria.In this paper,the research progresses of the classification,structures and functions of the lactate dehydrogenases,as well as the microbial genuses and synthetic metabolic pathway of the lactic acid production were summarized.This review will assist in understanding the metabolic regulation of lactic acid synthesis in many different species and genera,hoping for theoretical guidance for others.
lactic acid bacteria;L-lactate dehydrogenase;D-lactate dehydrogenase;classification;structure and function
TS201.1
A
1002-0306(2016)08-0369-05
10.13386/j.issn1002-0306.2016.08.069
2015-09-08
黃艷娜(1980-),女,博士,工程師,研究方向:益生菌代謝工程及代謝調控,E-mail:huangyanna@brightdairy.com。
*通訊作者:游春蘋(1981-),女,博士,高級工程師,研究方向:益生菌功能研究,E-mail:youchunping@brightdairy.com。
國家“十二五”科技支撐計劃(2013BAD18B01,2012BAD12B08)。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
