組學技術在鑒定及預測豬肉質量特性生物標志物中的應用
2016-09-13 01:02:52劉昊天李媛媛孔保華
食品工業科技 2016年13期
劉昊天,李媛媛,孔保華
(東北農業大學食品學院,哈爾濱黑龍江 150030)
?
組學技術在鑒定及預測豬肉質量特性生物標志物中的應用
劉昊天,李媛媛,孔保華*
(東北農業大學食品學院,哈爾濱黑龍江 150030)
利用組學技術鑒別出的生物標志物在豬肉的加工和感官特性中得到了一定的應用。本文從生物標志物的定義和基本的鑒別方法出發,重點綜述了灰白肉(PSE),肉的嫩度,肌間脂肪含量以及肉的剪切應力的生物標志物的鑒定,介紹了生物標志物預測豬肉的加工及感官品質的研究進展,并對生物標志物在今后肉品科學研究中的應用進行展望,以期在豬肉制品生產鏈上對產品的質量進行更好的管理。
組學技術,豬肉質量,生物標志物
我國是豬肉生產大國,目前隨著生活水平的提高,人們對肉及肉制品的品質需求也在逐步上升。影響豬肉品質的因素很多,包括豬的品種、性別、飼養條件等,這些因素會影響宰后肉的蒸煮損失、滴水損失、肌間脂肪含量、嫩度和風味等,進一步影響豬肉與肉產品的加工特性及感官質量[1]。因組學技術其具有高通量、速度快等優點,因此利用組學技術識別與肉類質量有關的生物標志物,再通過生物標志物來預測肉品的質量特性,現已成為目前研究熱點。
利用組學技術可以使蛋白質或代謝物中數百個甚至數千個基因同時進行分析,因此一些特征蛋白質,mRNA或者與肉的質量特征相關的代謝產物可以作為對應的生物標志物在豬肉中檢測出來[2-3]。利用檢測出的生物標志物可以對豬肉制品的質量進行評估和預測,這種快速檢測方法的不斷發展,可以更好的對胴體進行評估,進而減少經濟損失。此外,還可以將其應用于改善動物的遺傳選擇和適應良種繁育體系以滿足預期的質量水平[4]。
本文簡要提及了常用的基因組學,蛋白質組學,轉錄組學及代謝組學技術,從生物標志物的定義和基本的鑒別方法出發,重點綜述了灰白肉(PSE),肉的嫩度,肌間脂肪含量以及肉的剪切應力的生物標志物的鑒定,介紹了生物標志物預測豬肉的加工及感官品質的研究進展,并對生物標志物在今后肉品科學研究中的應用進行展望,最終目的是為了在實際應用中在豬肉制品生產鏈上對產品的質量進行更好的管理。
1 生物標志物定義及鑒別中常用的組學技術
1.1生物標志物的定義
生物標志物(Biomarker)是指可以標記系統、器官、組織、細胞及亞細胞結構或功能的改變或可能發生的改變的生化指標,是生物體受到嚴重損害之前,在不同生物學水平(分子、細胞、個體等)上因受環境污染物影響而異常化的信號指標[1]。它可以對嚴重毒性傷害提供早期警報。這種信號指標是細胞分子結構和功能的變化、某一生化代謝過程的變化或生成異常的代謝產物或其含量,也可能是某一生理活動或某一生理活性物質的異常表現,亦或是個體表現出的異常現象,或是種群或群落的異常變化等,而換句話來說,生物標志物就是反映機體生物學進程及狀態的標記[5]。
1.2生物標志物鑒別中涉及的組學技術
1.2.1基因組學技術基因組學主要研究的是基因組的功能,關于基因組及測序工具的發展使得功能性或表達性基因組學也得到了發展[3]。利用基因組學可以在特定的序列上或使用特定的工具來同時分析數百或數千個基因的變化(單核苷酸多態性位點與芯片),近年來基因組學技術已經逐漸被應用于研究與肉品質量有所關聯的基因的表達水平或表達豐度上。
1.2.2蛋白質組學蛋白質組學是研究細胞內所有蛋白質及其動態變化規律的科學。蛋白質組學技術是一種用來確定新肽的更綜合的方法,不僅能在蛋白質水平上驗證基因表達和基因精細模式,還能改進蛋白質序列數據庫[6]。對于蛋白質來說,為了測定蛋白質生物標志物的含量,常用免疫斑點技術測定分析肌肉樣品中的大量蛋白質。
1.2.3轉錄組學對于轉錄物來說,在基因組中能夠挑選出至少3000個涉及肌肉生物過程或肉品質量的基因(相當于13409個寡核苷酸探針),而目前蛋白質組學與轉錄組學的研究或科學出版物都集中在DNA芯片上[7]。DNA芯片又稱為基因芯片,基因芯片作為一種新技術,不僅準確、快速、高效,而且具有高通量等特點。基因芯片的測序原理是雜交測序方法,即通過與一組已知序列的核酸探針雜交進行核酸序列測定的方法。具體見圖1,在一塊基片表面固定了序列已知的靶核苷酸的探針,當溶液中帶有熒光標記的核酸序列TATGCAATCTAG,與基因芯片上對應位置的核酸探針產生互補匹配時,通過確定熒光強度最強的探針位置,獲得一組序列完全互補的探針序列,據此可重組出靶核酸的序列[8]。
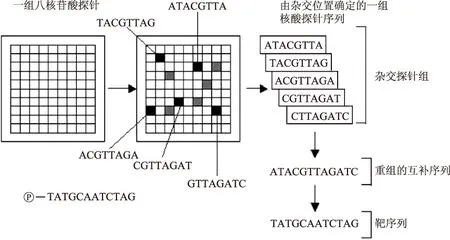
圖1 基因芯片的測序原理[8]Fig.1 The sequencing principle of gene chip[8]
1.2.4代謝組學代謝組學是以組群指標分析為基礎,以高通量檢測和數據處理為手段,以信息建模與系統整合為目標的系統生物學的一個分支,是繼基因組學、轉錄組學、蛋白質組學后系統生物學的另一重要研究領域,它是研究生物體系受外部刺激所產生的所有代謝產物變化的科學,所關注的是代謝循環中分子量小于1000的小分子代謝物的變化,反映的是外界刺激或遺傳修飾的細胞或組織的代謝應答變化[3,6]。
2 利用組學技術鑒定豬肉質量特性的生物標志物
同其他物種一樣,豬肉的感官及質量特性也同樣取決于動物的遺傳因素、飼養和屠宰條件以及豬肉在加工過程中發生的復雜的物理化學反應變化。盡管許多生產廠商在生產及加工過程中盡可能的避免這些影響豬肉質量的因素,但豬肉及其制品的質量特性的差異性仍然很高。通過在胴體上或者分割加工后的生物標志物可以提前來預測以后的肉品質量,因此可以更好的對產品進行分級,降低損失,提高豬肉生產商的利益。目前,對于影響肉品質量的特征如蒸煮損失,肌間脂肪含量,PSE肉以及剪切力的生物標志物的鑒定已經得到了廣泛的研究[1-3,9]。最近,一種基于實驗設計誘導一個或多個肉品質量性狀同時產生較高的個體差異的方法已經研制成功并且得到了廣泛的應用[10]。在豬肉中,對背最長肌的大部分研究已經完成,而一些研究已經開始考慮對半膜肌的研究[8-11]。
2.1PSE肉的生物標志物
PSE(pale,soft,and exudative)肉是一種常見的豬肉的質量缺陷,它主要是由遺傳或非遺傳因素所導致產生的一種異常肉,PSE發生會使肉的生理和生化機制發生了一定改變,使豬肉產生不良的質量特性,比如表面滲水過多、變色、質地變軟等,因此通過鑒別PSE的生物標志物可以避免肉品加工行業的經濟損失。Damon 等人[12]分析出PSE肉內糖酵解途徑中酶的基因編碼的上調也可以作為PSE肉的生物標志物。Laville 等人[13]通過蛋白質組學的研究發現,在PSE肉中肌原纖維蛋白的溶解度較低,肌原纖維蛋白會產生一定程度的降解并產生小熱休克蛋白,這些也可以作為PSE肉的生物標志物。有一種遺傳因子叫氟烷基因(hal gene),常用N表示顯性,用n表示隱性。當該基因型為nn時(即正常豬),豬極易出現應激綜合癥(PSS),是導致PSE肉形成的主要原因[14]。所以氟烷基因也可以作為PSE肉的生物標志物。但是Laville等人[15]進一步研究發現與顯性純合體(NN)的豬相比,隱性純合子(nn)遺傳型的豬的半膜肌在蘭尼受體基因(RyR1)基因座上也可以檢測出其蛋白質溶解度的降低、小熱休克蛋白以及與氧化代謝有關系的蛋白質的增多,這也說明了PSE缺陷可能與遺傳因素有關,該研究可以得出小熱休克蛋白可被作為宰后初期的有PSE缺陷的豬肉的生物標志物。因此,就目前研究而言,發現的PSE肉的生物標志物主要有糖酵解途徑中酶的基因編碼的上調,小熱休克蛋白,氟烷基因等。
2.2肉的嫩度的生物標志物
一般來講,通過力學測量例如剪切應力(SF)可以評定肌肉嫩度,或者由經過培訓的專業人員進行感官評分來確定最適的肉的嫩度,但是目前常用的方法是感官評定與力學測量所得的結果結合起來進行綜合的評定[16]。隨著基因組學、蛋白質組學、代謝組學、計算生物學、以及生物化學等數門學科研究的深入,為了得到感官質量高的豬肉,可以將上述組學進行有機的結合,通過鑒定其生物標志物研究影響豬肉嫩度因素的內部機制,通過轉錄物、蛋白質或者代謝物的含量高低來分析肌肉組織嫩度的大小。
Picard等人[6]對三種豬的半膜肌和背最長肌進行嫩度的測定,并利用得到的結果建立了預測嫩度的方程。Picard等人[6]首先分別從半鍵肌和背最長肌中提取出蛋白質,將提取出的蛋白質與靶向蛋白中的特異性抗體進行雜交。隨后加入帶有標記物的抗體,使標記通過抗體和相應抗體的結合間接地交聯于纖維素膜上。最后加入標記物與相應的底物后,標記物與底物作用形成不溶性產物,加入熒光染色劑后會呈現斑點狀著色,從而判定結果。而在這之前,抗體的特異性以及使用的最佳條件需要通過蛋白質印跡法(Western-Blot)來確定[7],接著用測得的蛋白質相對含量建立方程式來預測肌肉的嫩度。Damon 等人[12]根據轉錄組學分析宰后20 min的豬肉大腿肌樣品,鑒別出豬肉宰后初期嫩度的生物標志物為肌原纖維蛋白的基因編碼的上調,并且肌原纖維蛋白的基因編碼的上調也涉及到肌動蛋白-肌球蛋白之間的相互作用以及肌節的完整性,因此肌節的完整性也與嫩度存在一定的相關性。Picard 等人[17]又對兩組嫩度不同的豬肉樣品中起主要作用的蛋白質的差異進行了研究,將兩組嫩度不同的豬肉樣品中的蛋白質通過二維凝膠電泳進行分離,進而通過統計實驗進行對比得到24 種蛋白質可能是嫩度的潛在生物標志物。這些蛋白質在糖酵解、氧化能量代謝、鈣代謝、肌肉的微觀結構、肌肉的收縮、肌肉的氧化、細胞凋亡、細胞的保護中起到了一定的作用,進而影響了肉的嫩度。此結論在其他文獻中也得到了證實,例如,Jia 等人[18]證實了Park7和PRDX6為嫩度的生物標志物,Hollung 等人[19]也證明了小清蛋白(parvalbumin)和膜聯蛋白(Annexin V)與鈣的代謝有關。Bernard等人[20]挑選了大約100 頭豬作為樣品,并利用基因芯片分析了飼養方式不同的同種豬的兩個肌肉部分。結果證明了 DNAJA1基因可能是豬肉韌性的生物標志物。他們還發現,有一組基因與豬肉的嫩度及剪切力之間存在著聯系,這組基因屬于熱休克蛋白族,或存在于能量代謝或脂肪代謝途徑中。Lomiwes等人[11]認為,一組屬于熱休克蛋白族的4 種基因可以用來解釋一些肉的嫩度變化,但是這些嫩度標記物或許只是對于某一群體而言。許多的研究小組已經開始使用基因芯片技術來對豬肉或者牛肉的質量進行預測或者研究其食用性質的影響[9]。
因此,就目前研究而言,發現的肉的嫩度的生物標志物主要有肌原纖維蛋白的基因編碼的上調,肌節的完整性,Park7,PRDX6,Parvalbumin和Annexin V。
2.3肌間脂肪的含量以及肉的剪切應力的生物標志物
肌間脂肪的含量在豬肉的食用品質及可接受性中起著重要的作用,而對于肌間脂肪含量的生物標記物的了解和鑒別,主要涉及了轉錄組學、蛋白質組學和代謝組學的相關知識。D’Alessandro 等人[21]結合差異蛋白質組學與定量代謝組學對肌間脂肪含量不同的兩個物種的 Casertana 豬和大白豬進行分析時,發現那些參與糖酵解反應支路的酶,如甘油-3-磷酸脫氫酶(為甘油合成提供基質)會在背最長肌中加速脂肪的合成。Liu 等人[22]研究來自相同遺傳背景的兩種豬,對比了100 kg背最長肌的肌間脂肪含量為1.36%和4.58%,這一研究結果表明了mRNA以及蛋白質在基因上表達水平的差異對葡萄糖、脂質、蛋白質的代謝過程,細胞間的信息傳遞,以及應激反應都有一定的影響。Hamill等人[23]利用轉錄組學對不同實驗組間肌間脂肪含量的差異進行了研究,結果表明了脂肪和蛋白質代謝、蛋白質的合成,細胞的結構和細胞的功能都與肌間脂肪含量有所關聯,更加確立了肌間脂肪含量與各種代謝和細胞途徑有關。
Liu 等人[22]同樣對肌間脂肪含量很高的豬肉的背最長肌進行了測定,得出了小清蛋白含量的減少會對背最長肌中脂肪的合成會有一定消極的影響,并證實了脂肪細胞的發育對肌間脂肪含量變化的具有一定的重要性。同時,Damon 等人[12]研究發現大量的 FABP4蛋白質(fatty acid binding protein 4,adipocyte)與肌肉脂肪細胞的數量或者肌間脂肪含量具有較高的正相關性。這表明FABP4蛋白質可以在豬肉中作為肌間脂肪含量的生物標志物。此外,Laville 等人[24]研究表明與脂代謝有關的FABP4蛋白質也在肌間脂肪含量較低但是具有較高剪切應力(SF)的豬肉中被發現,這與肌間脂肪含量與剪切應力之間呈現負相關性的結論相一致。Hamill 等人[23]的研究表明在豬肉的背最長肌中,蛋白質合成基因表達的減少以及參與蛋白質降解的基因表達的增多同樣與SF的降低有關。就目前研究而言,發現的肌間脂肪含量和剪切力生物標志物主要有甘油-3-磷酸脫氫酶,mRNA以及蛋白質在基因上表達水平的差異,小清蛋白的含量,FABP4蛋白質等。
3 組學技術鑒別出的生物標志物預測豬肉的質量特性
許多研究為了同時分析不同物種或是生長環境中不同的豬的許多質量特性之間存在的差異性,結合生物學知識,利用生物標志物來研究多種豬肉質量特性之間的相關性及差異性。近期Rohart 等人[25]研究表明,不同于肉品質量特性,一些潛在的經濟效益特性(如瘦肉率,平均日采食量)可以在測試結束期(當豬的權重約為110 kg體重)時通過動物在生長期(當豬的體重約60 kg)的代謝數據很好的進行預測。
對于肉品的質量特征,Lebret 等人[26]實驗發現將兩種豬(法國本地的Basque和大白豬)飼養在不同的生長環境中,研究結果表明不同的生長環境同樣會影響到豬肉的品質。其研究關于背最長肌基因芯片轉錄分析結果顯示,不同物種之間的肌肉生理機能和肉品質量性狀的不同與代謝過程、骨骼肌結構和組織、細胞外基質、溶酶體以及蛋白水解作用的差異有關。Lebret 等人[26]在 Basque 這個物種的豬中進行了進一步的研究,結果表明,相對于傳統飼養,現今飼養體系導致了參與控制肌肉結構以及熱反應(小熱休克蛋白)的基因的過度表達。聯系到其質量特性的差異,上述這些數據可以作為鑒別豬肉特性的生物標志物進而預測豬肉的質量特性。
Lebret B 等人[27]從更廣范圍選取不同物種和不同生長環境的50頭豬,鑒別與它們的加工特性和感官質量包括極限pH、滴水損失、亮度(L*)、紅度(a*)、色相(h°)、肌間脂肪含量、剪切應力(SF)和嫩度等相聯系的生物標志物。研究結果表明基因芯片表達與肉品質量特性之間建立了多達數千種聯系,其中與a*值的聯系有140種左右,與嫩度的聯系達到2892種。之后,Lebret B等人[27]從中挑選出40個具有高度相關系數值或者與生物過程相關的基因。RNA的逆轉錄(RT)和cDNA的聚合酶鏈式擴增反應(PCR)相結合的技術,即RT-PCR技術是更為精確的一種方法來檢測同一樣品中它們之間的聯系的分子生物學技術,因此Damon等人[28]進一步利用RT-PCR技術鑒別出113種與質量特性之間存在的聯系的轉錄物,通過建立多元線性回歸模型進行分析,分析結果表明,3~5個基因可以用來解釋多達59%的肉品質量特性的變化,因此這3~5個基因可以作為上述質量特性的生物標志物。Lebret等人[29]挑選了100只商業杜洛克豬、長白豬、約克夏豬進行研究,結果表明檢測到19 項轉錄物與質量特性之間的聯系(R2≤0.94)包括6 項與極限pH相聯系的轉錄物,5項與L*相聯系的轉錄物,4項與滴水損失相聯系的轉錄物,1 項與肌間脂肪含量相聯系的轉錄物,1項與與嫩度相聯系的轉錄物,其中在這些生物標志物中CA3(碳酸酐酶III)與肌間脂肪含量成反比,FOS(FBJ小鼠骨肉瘤病毒癌基因同源物)涉及到鈣的傳輸及轉錄,并且與極限pH、滴水損失、L*、h°呈負相關性,因此這些轉錄物皆可作為特定質量特性的生物標志物。Pierzchala等人[30]與同樣選取和Lebret等人[29]的實驗一致的商業杜洛克豬、長白豬和約克夏豬來進行豬肉質量的生物標志物的外部鑒定實驗。但在鑒定生物標志物之前,通過轉錄組學對比實驗設計,測定了新鮮豬肉的工藝品質。得到的結果為2項與滴水損失相聯系的轉錄物,3項與極限pH相聯系的轉錄物,1 項與a*相聯系的轉錄物,2項與L*相聯系的轉錄物,但是他們的研究中同一質量特性涉及的基因與Lebret等人[29]的實驗結果有所不同,因此生物標志物不同。
這些和質量特性相關的肌肉蛋白組被考慮用來作為宰后初期的豬的生物標志物。tePas等人[30]通過分析背最長肌樣品中的蛋白質組,發現分析結果中的峰或峰組與肉品質量特征之間存在一定的關系,尤其是滴水損失和極限pH與峰或峰組之間存在著最高的相關性。這一研究結果表明,測定蛋白質組生物標志物來預測肉品質量的預測精度要高于測定單一的蛋白質的生物標志物。然而,te Pas等人[31]認為,如果要想進一步測定蛋白質的基礎峰值模式,需要開發出更方便,成本更低,更加快速的方法(例如試紙條)來量化這些生物標志物。
4 前景及展望
目前組學技術具有高通量、操作簡便、特異性強、靈敏度高和可預測性等諸多優點,現有研究已經證明了可以通過蛋白質組學,轉錄組學和代謝組學等方法將與豬肉質量特性有關的生物標志物鑒別出來。利用生物標志物對肉制品質量的預測可以更好的發展豬肉產業和更好的控制產品質量,但仍需進一步改善其預測能力(轉錄組標記)或量化方法(蛋白質組標記)。因此,今后的研究應致力于基于驗證和鑒定肉品質量等級(例如豬肉質量等級)和感官質量的轉錄組生物標志物的發展,這樣可以對評定肉質等級提供更為方便、快速、價廉的方法。而對生物標志物鑒定的最終的目的是在對胴體進行分級或是屠宰初期對胴體進行感官和加工質量的預測,從而提高豬肉生產商的利益,減少損失。
[1]孔保華,韓建春. 肉品科學與技術[M]. 北京:中國輕工業出版社,2011,16(3):8-10.
[2]殷朝敏,雷靖行. 基因表達系列分析技術在真菌功能基因組學中的應用[J]. 北京:生物技術通報,2011,33(1):25-28.
[3]D’Alessandro A,Rinalducci S,Marocco C,et al. An Omics window on the pork tenderness network[J]. Journal of Proteomics,2012,84(1):4360-4380.
[4]謝志修. 生物技術在食品檢測方面的應用[M]. 生物技術通報,2010,33(15):412-417.
[5]吳曉薇,黃國城. 生物標志物的研究進展[J]. 廣東畜牧獸醫科技,2008,30(12):3405-3409.
[6]Picard B,Micol D,Cassar-Malek I,et al. Meat and fish flesh quality with proteomic applications[J]. Animal Frontiers,2012,2(4):18-25.
[7]Hocquette J F,Bernard-Capel C,Vidal V,et al. The GENOTEND chip:A new tool to analyse gene expression in muscles of pig for pork quality prediction[J]. BMC Veterinary Research,2012,32(3):18-35.
[8]鄒宗亮,王志清,王升啟. 基因芯片技術研究進展[J]. 高技術通訊,2000,24(16):314-318.
[9]Hiller B,Hocquette J F,Cassar-Malek I,et al. Dietaryn-3 fatty acid effects on gene expression in pig longissimus muscle as assessed by microarray/q RT-PCR methodology[J]. British Journal of Nutrition,2012,10(8),858-863.
[10]Guillemin N,Cassar-Malek I,Hocquette J,et al. La maitrise de la tendreté de la viande pig:identification de marqueurs biologiques[J]. INRA Productions Animal,2009,22(4):331-344.
[11]Lomiwes D,Farouk M M,Wiklund E. Small heat shock proteins and their role in meat tenderness:A review[J]. Meat Science,2014,96(1):26-40.
[12]Damon M,Louveau I,Lefaucheur L,et al. Number of intramuscular adipocytes and fatty acid binding protein-4 content are significant indicators of intramuscular fat level in crossbred Large White X Duroc pigs[J]. Journal of Animal Science,2006,8(4):1083-1092.
[13]Laville E,Sayd T,Sante-Lhoutellier V,et al. Characterisation of PSE zones in semimembranosus pig muscle[J]. Meat Science,2005,7(10):167-172.
[14]董佩佩,劉志軍,賈俊靜. 影響豬肉品質的基因及研究進展[J]. 飼料研究,2010,(01):46-48.
[15]Laville E,Sayd T,Terlouw C,et al. Differences in pig muscle proteome according to HAL genotype:Implication for meat quality defects[J]. Journal of Agricultural and Food Chemistry,2009,5(7):4913-4923.
[16]Chriki S,Renand G,Picard B,et al. Meta-analysis of the relationships between pork tenderness and muscle characteristics[J]. Livestock Science,2013,15(5):424-434.
[17]Picard B,Gagaoua M,Micol D,et al. Inverse relationships between biomarkers and pig tenderness according to contractile and metabolic properties of the muscle[J]. Journal of Agricultural and Food Chemistry,2014,62(40):9808-9818.
[18]Jia X,Veiseth K E,Grove H,et al. Peroxiredoxin-6-A potential protein marker for meat tenderness in pork longissimus thoracis muscle[J]. Journal of Animal Science,2009,87(7):2391-2399.
[19]Hollung K,Veiseth E,Jia X,et al. Application of proteomics to understand the molecular mechanisms behind meat quality[J]. Meat Science,2007,7(7):97-114.
[20]Bernard C,Cassar-Malek I,Le Cunff M,et al. New indicators of pork sensory quality revealed by expression of specific genes[J]. Journal of Agricultural and Food Chemistry,2007,55(13):5229-5237.
[21]D’Alessandro A,Marocco C,Zolla V,et al. Meat quality of the longissimus lumborum muscle of Casertana and Large White pigs:Metabolomics and proteomics intertwined[J]. Journal of Proteomics,2011,7(5):610-627.
[22]Liu J,Damon M,Guitton N,et al. Differentially-expressed genes in pig Longissimus muscles with contrasting levels of fat,as identified by combined transcriptomic,reverse transcription PCR,and proteomic analyses[J]. Journal of Agricultural and Food Chemistry,2009,5(7):3808-3817.
[23]Hamill R M,McBryan J,MCGee C,et al. Functional analysis of muscle gene expression profiles associated with tenderness and intramuscular fat content in pork[J]. Meat Science,2014,9(2):440-450.
[24]Laville E,Sayd T,Terlouw C,et al. Comparison of sarcoplasmic proteomes between two groups of pig muscles selected for shear force of cooked meat[J]. Journal of Agricultural and Food Chemistry,2007,15(5):5834-5841.
[25]Rohart F,Paris A,Laurent B,et al. Phenotypic prediction based on metabolomic data for growing pigs from three main European breeds[J]. Journal of Animal Science,2012,9(10):4729-4740.
[26]Lebret B,Denieul K,Damon M. Muscle transcriptome profiles highlight biomarkers of pig production system and high meat quality[J]. Journal of Animal Science,2014,1(8):44-49.
[27]Lebret B,Ecolan P,Bonhomme K,et al. Influence of production system in local and conventional pig breeds on stress indicators at slaughter,muscle and meat traits and pork eating quality[J]. Journal of Animal Science,2014,1(3):34-41.
[28]Damon M,Wyszynska-Koko J,Vincent A,et al. Comparison of muscle transcriptome between pigs with divergent meat quality phenotypes identifies genes related to muscle metabolism and structure[J]. Journal of Animal Science,2012,7(3):33-39.
[29]Lebret B,Denieul K,Vincent A,et al. Identification by transcriptomics of biomarkers of pork quality[J]. Meat Science,2015,9(1):44-50.
[30]Pierzchala M,Hoekman A J,Urbanski P,et al. Validation of biomarkers for loint meat quality(M. longissimus)of pigs[J]. Journal of Animal Breeding and Genetics,2014,131(3):258-270.
[31]te Pas M F,Kruijt L,Pierzchala M,et al. Identification of proteomic biomarkers in M. Longissimus dorsias potential predictors of pork quality[J]. Meat Science,2013,9(5):679-687.
Application of omics techniques in identification and predication of biomarkers of quality attributes of pork:a review
LIU Hao-tian,LI Yuan-yuan,KONG Bao-hua*
(College of Food Science,Northeast Agricultural University,Harbin 150030,China)
Biomarkers which are identified by omics techniques has received some applications in technological and sensory attributes of pork. In this paper,the definition and basic method of identification of biomarkers were introduced briefly,and its identification in pork samples were reviewed in analyzing pale soft exudative(PSE)meat,tenderness,intramuscular fat(IMF)content and meat shear force. The prediction of biomarkers was introduced briefly in pork qualities. Finally,applications of biomarkers in further meat research were prospected and this technology may play a greater role in the field of product quality.
genomics;pork quality;biomarkers
2015-12-16
劉昊天(1992-),男,碩士研究生,研究方向:畜產品加工,E-mail:18246142997@163.com。
孔保華(1963-),女,博士,教授,研究方向:畜產品加工, E-mail:kongbh@163.com。
國家自然科學基金(31471599);黑龍江省應用技術研究與開發重點計劃(GA15B302)。
TS251.1
A
1002-0306(2016)13-0381-05
10.13386/j.issn1002-0306.2016.13.070
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
航空世界(2020年10期)2020-01-19 14:36:20
中國生殖健康(2019年2期)2019-08-23 08:12:08
產品可靠性報告(2017年7期)2017-09-05 09:49:12
