組蛋白去乙酰化酶抑制劑對膠質瘤細胞增殖及E2F1表達的影響
2016-09-06 08:57:04袁忠民梁琳唐曉梅
新醫學 2016年8期
關鍵詞:檢測
袁忠民 梁琳 唐曉梅
?
·基礎研究論著·
組蛋白去乙酰化酶抑制劑對膠質瘤細胞增殖及E2F1表達的影響
袁忠民梁琳唐曉梅
目的探討組蛋白去乙酰化酶抑制劑(HDACIs)對膠質瘤細胞增殖及E2F1表達的影響。方法體外培養膠質瘤細胞U251,用2個E2F1干擾質粒she2f1a和she2f1b轉染細胞,空載體BS/U6為對照,噻唑藍比色法及核染色法檢測細胞增殖率;TSA處理U251細胞2 h、4 h、8 h,及多種HDACIs包括曲古菌素A(TSA)TSA、伏立諾他(SAHA)、M344、丁酸鈉(NaBu)NaBu、丙戊酸(VPA)、苯甲酰胺(M344)處理細胞8 h,二甲基亞砜(DMSO)處理為對照,蛋白免疫印跡法檢測E2F1表達;用TSA處理8、12、24 h,DMSO處理為對照,噻唑藍比色法分析細胞增殖率;采用多種HDACIs(TSA,SAHA, NaBu, VPA,M344)處理細胞24 h,噻唑藍比色法分析細胞增殖率。結果與BS/U6比較,2個E2F1干擾質粒she2f1a和she2f1b都能沉默E2F1表達和抑制細胞增殖(P<0.05);與對照組比較,TSA處理細胞2 h和4 h未見抑制E2F1表達,處理8 h能抑制E2F1表達,其它HDACIs均能抑制E2F1表達;與對照組相比,TSA處理8 h未影響細胞增殖率(P>0.05),但TSA處理12 h能抑制細胞增殖(P<0.05),處理24 h抑制效應更強(P<0.05)。而且,其它HDACIs均能抑制細胞增殖(P<0.05)。 結論HDACIs能抑制膠質瘤細胞E2F1表達及其介導的細胞增殖。
神經膠質瘤;E2F1;組蛋白去乙酰化酶抑制劑
神經膠質瘤是中樞神經系統最常見的原發性腫瘤,惡性增殖是其主要特點之一,亦是造成高復發率的首要原因[1]。基于分子生物學和遺傳學的研究表明,在高級別(WHO分級為Ⅲ和Ⅳ級)的膠質瘤樣本中,調控細胞周期的關鍵蛋白的表達和功能出現明顯異常[2]。如約25%的腫瘤樣本中發現抑癌基因Rb缺失,約15%的樣本中發現細胞周期素依賴性蛋白激酶 (CDKs)家族成員CDK4或CDK6活性異常增強,而抑癌基因p16在多達50%~70%的樣本中缺失等[3-4]。這些信號的異常使得下游E2F蛋白家族(共有8個成員,E2F1~8)中關鍵分子轉錄因子E2F1的活性失去控制,失控的E2F1驅動細胞持續分裂,惡性增殖[5]。本研究采用小分子干擾、噻唑藍比色法、蛋白免疫印跡法、轉染等技術,檢測多種組蛋白去乙酰化酶抑制劑(HDACIs)包括曲古柳菌素(TSA)、伏立諾他(SAHA)、丁酸鈉(NaBu)、丙戊酸(VPA)、苯甲酰胺(M344)能否靶向影響E2F1表達及其功能。
材料與方法
一、U251細胞培養
神經膠質瘤細胞株U251的培養參考已發表文章進行[6]。U251細胞購自中科院上海細胞庫。培養基為高糖DMEM(美國Invitrogen公司)加10%胎牛血清(美國Hyclone公司)。
二、U251細胞轉染
轉染參照Thermo公司的說明書,當U251細胞生長至融合度70%以上時,用Turbofect Transfection Reagent分別將質粒BS/U6、she2f1a、she2f1b轉染到U251細胞內,于37℃恒溫箱培養48 h。轉染細胞分為3組,轉染BS/U6作為對照組,2個E2F1干擾質粒為she2f1a組和she2f1b組(24孔板,每孔0.5 μg)。質粒BS/U6、she2f1a,she2f1b的靶序列參照已發表文章設計[5]。
三、細胞加藥處理
參考已發表文章進行[6]。用胰酶消化U251細胞,種入24孔板,當細胞生長至融合度為50%~60%時,將細胞分為對照組[二甲基亞砜(DMSO, Sigma)處理]和組蛋白去乙酰化酶抑制劑組(分別用TSA、M344、NaBu、VPA處理),分別處理8、12、24 h后進行噻唑藍比色法分析或Hoechst 33258核染色后計數細胞數。
四、蛋白免疫印跡
參考已發表文章進行[7]。為了檢測曲古菌素A處理U251細胞后對E2F1的抑制作用是否發生在抑制細胞增殖之前,我們用曲古菌素A處理U251細胞2 h、4 h、8 h,用細胞裂解液(50 mmol/L Tris pH 7.4、150 mmol/L NaCl、2 mmol/L EGTA、1 mmol/L EDTA、1% Triton X-100、2.5 mmol/L Sodium pyrophosphate、1 mmol/L Glycerophosphate、1 mmol/L Na3VO4、1 mmol/L PMSF、1 μg/ml Leupeptin、5 μg/ml Aprotinin)裂解細胞檢測E2F1的表達。同時,用多種HDACIs如0.5 μmol/L TSA、6 mmol/L NaBu、6 mmol/L VPA、1 μmol/L SAHA、1 μmol/L M344處理U251細胞8 h后裂解細胞檢測E2F1表達。用針對E2F1的小分子干擾質粒轉染U251細胞48 h后裂解細胞,檢測E2F1表達。采用4%集成膠和10%分離膠,200 V電壓電泳45 min,100 V電壓轉膜60 min。脫脂奶粉(5%)室溫封閉1 h。一抗(anti-E2F1 1∶1 000,anti-GAPDH 1∶5 000,所有抗體均購自美國Cell Signaling Technology公司)孵育過夜(4℃),辣根過氧化物酶(HRP)標記的二抗(美國Jackson Immuno Research公司)室溫孵育1 h。增強化學發光后曝光。
五、噻唑藍比色法(MTT)分析
參考已發表文章進行[7]。用胰酶消化U251細胞,種入96孔板,每個處理為6個平行孔。只加培養液不加細胞作空白對照。為防止MTT分析時有邊緣效應,板周邊的孔不加細胞。當U251細胞生長至融合度為50%~60%時,轉染質粒BS/U6、she2f1a、she2f1b共48 h,每孔加入MTT溶液(5 g/L)20 μl培養4 h,棄培養液,然后每孔加入DMSO 150 μl,振蕩10 min,選擇490 nm波長,在酶聯免疫檢測儀上測定全部孔包括平行孔的吸光度A值。
六、統計學處理
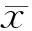
結 果
一、沉默E2F1抑制U251細胞增殖
與對照相比,2個E2F1干擾質粒she2f1a和she2f1b均能抑制E2F1表達和細胞增殖(BS/U6: 增殖率=100%; she2f1a:增殖率=78±6,n=3,P=0.02;she2f1b:增殖率=83±5, n=3, P=0.03)(圖1 A, B)。核染色后統計細胞數,與對照組比較,干擾E2F1組的細胞減少(BS/U6: 100%; she2f1a:增殖率=79±2, n=3,P=0.007;she2f1b:增殖率=77±6,n=3, P=0.01)(圖1 C)。
二、HDACIs抑制U251細胞E2F1表達
與對照組比較,TSA處理細胞8 h能抑制U251細胞E2F1表達(圖2 A)。而且,與TSA一樣,NaBu、VPA、SAHA和M344均能抑制E2F1表達(圖2 B)。

圖1 沉默E2F1抑制U251細胞增殖
A:用BS/U6、she2f1a、she2f1b轉染U251細胞48 h,蛋白免疫印跡法檢查E2F1表達,GAPDH為標準化內參;B:BS/U6、she2f1a、she2f1b轉染U251細胞48 h后MTT分析;C:BS/U6、she2f1a、she2f1b轉染U251細胞48 h, Hoechst 33258染核后,熒光顯微鏡拍照(×200)

圖2 HDACIss抑制U251細胞E2F1表達
A:0.5 μmol/L TSA處理U251細胞2 h、4 h和8 h,裂解細胞,蛋白免疫印跡法檢測E2F1蛋白表達;B:用多種HDACIs包括0.5 μmol/L TSA、6 mmol/L NaBu、6 mmol/L VPA、1 μmol/L SAHA和1 μmol/L M344分別處理U251細胞12 h,DMSO作為溶劑對照(CTL),裂解細胞,蛋白免疫印跡法檢測E2F1表達
三、HDACIs抑制U251細胞增殖
多組間比較差異有統計學意義(F=29.39,P=0.01)。與對照組比較,TSA處理細胞8 h不影響增殖率(P>0.05),但處理12 h抑制細胞增殖(P<0.05),處理24 h抑制效應更明顯(P<0.05),見表1。
HDACIs包括NaBu、VPA、SAHA、M344處理U251細胞24 h均能抑制細胞增殖,各組與DMSO相比,差異均有統計學意義(P均<0.05),見表2。
討 論
我們的研究顯示,沉默E2F1抑制神經膠質瘤細胞株U251;TSA抑制E2F1表達發生在抑制細胞增殖之前;所使用的HDACIs均能抑制E2F1表達和腫瘤細胞增殖。筆者查閱文獻發現,這是首次報道HDACIs能靶向抑制E2F1表達及其介導的膠質瘤細胞增殖。

表1 TSA處理細胞不同時間對U251細胞增殖的影響±s)
注:DMSO為對照組;增殖率=TSA處理組細胞數/DMSO組細胞數×100%(特定時間點)

表2 HDACIs處理U251細胞24 h對細胞增殖的影響
注:aDMSO對照組與各個HDACIs處理組比較
E2F1是第一個被鑒定的具有轉錄活性的E2F家族成員,其活性受上游CDKs/Rb等分子精確調控,在調節細胞周期由G1期進入S期中發揮主導作用[8-9]。由于p16、Rb、p21等抑癌基因在高級別膠質瘤樣本中發生高頻缺失,E2F1對膠質瘤細胞增殖的作用在過去的研究中得到格外關注[10-11]。多家實驗室包括我們均觀察到利用小分子干擾RNA或反義核苷酸片段抑制E2F1表達抑制了膠質瘤細胞增殖。而且,多個微小RNA(miRNA)包括miRNA-329、miRNA-106a、miRNA-136、miRNA-138等均通過抑制E2F1表達抑制膠質瘤細胞增殖[12-15]。可見,在上游抑制信號缺失的情況下,E2F1的持續激活是造成膠質瘤細胞惡性增殖的關鍵原因之一,因而其是一個理想的分子靶點。
HDACIs的抗癌作用緣于其突出的促進腫瘤細胞凋亡或抑制細胞周期效應,而這些效應在正常細胞中的敏感性明顯下降[16]。有文獻報道HDACIs通過上調E2F1進而上調Bim等BH3-only蛋白促發線粒體依賴的細胞凋亡[17]。但在膠質瘤細胞株U251中,無論加大HDACIs濃度或是延長處理時間,我們都只檢測到E2F1表達被抑制,而沒有檢測到E2F1被上調。可見,HDACIs抑制E2F1表達也是重要的抗癌機制之一。
總之,我們發現了HDACIs抑制E2F1表達進而抑制膠質瘤增殖的新機制。我們的研究一方面進一步支持了E2F1是治療膠質瘤的理想靶點,另一方面提示HDACIs可能是的治療膠質瘤的理想藥物。未來我們的工作將進一步明確HDACIs調控E2F1表達的分子機制。
[1]陳正和,陳忠平. 腦膠質瘤的治療進展. 新醫學,2015,46(7):417-422.
[2]Lewis KM, Petritsch C. Asymmetric cell division: implication for glioma dvelopment and treatment. Transl Neurosi, 2013,4(4):484-503.
[3]Furnari FB, Fenton T, Bachoo RM, Mukasa A, Stommel JM, Stegh A, Hahn WC, Ligon KL, Louis DN, Brennan C, Chin L, DePinho RA, Cavenee WK. Malignant astrocytic glioma: genetics, biology, and paths to treatment. Gene Dev, 2007,21(21):2683-2710.
[4]Fischer M, Grossmann P, Padi M, DeCaprio JA. Integration of TP53, DREAM, MMB-FOXM1 and RB-E2F target gene analyses identifies cell cycle gene regulatory networks. Nucleic Acids Res, 2016 Jun 8. pii: gkw523. [Epub ahead of print].
[5]Johnson J, Thijssen B, McDermott U, Garnett M, Wessels LF, Bernards R. Targeting the RB-E2F pathway in breast cancer. Oncogene, 2016 Feb 29.[Epub ahead of print].
[6]袁忠民,曾敏靈,唐曉梅,伍燕娜,何國振. 組蛋白去乙酰化酶抑制劑對膠質瘤細胞增殖及MKK7表達的影響. 腫瘤防治研究, 2016, 43(1): 1-5.
[7]He W, Wu Y, Tang X, Xia Y, He G, Min Z, Li C, Xiong S, Shi Z, Lu Y, Yuan Z. HDAC inhibitors suppress c-Jun/Fra-1-mediated proliferation through transcriptionally downregulating MKK7 and Raf1 in neuroblastoma cell. Oncotarget,2016, 7(6):6727-6747,
[8]Udayakumar T, Shareef MM, Diaz DA, Ahmed MM, Pollack A. The E2F1/Rb and p53/MDM2 pathways in DNA repair and apoptosis: understanding the crosstalk to develop novel strategies for prostate cancer radiotherapy. Semin Radiat Oncol, 2010,20(4):258-266.
[9]Meng P, Ghosh R. Transcription addiction: can we garner the Yin and Yang functions of E2F1 for cancer therapy? Cell Death Dis, 2014,5:e1360.
[10]袁忠民,李明昌,杜漢強,陸永建. 肝癌缺失基因-1不同亞型在人神經膠質瘤組織中的表達及意義. 新醫學,2010,41(2):98-10.
[11]Lladó V, López DJ, Ibarguren M, Alonso M, Soriano JB, Escribá PV, Busquets X Regulation of the cancer cell membrane lipid composition by NaCHOleate: effects on cell signaling and therapeutical relevance in glioma. Biochim Biophys Acta, 2014,1838(6):1619-1627.
[12]Xiao B, Tan L, He B, Liu Z, Xu R. MiRNA-329 targeting E2F1 inhibits cell proliferation in glioma cells. J Transl Me, 2013,11:172.
[13]Yang G, Zhang R, Chen X, Mu Y, Ai J, Shi C, Liu Y, Shi C, Sun L, Rainov NG, Li H, Yang B, Zhao S. MiR-106a inhibits glioma cell growth by targeting E2F1 independent of p53 status. J Mol Med, 2011,89(10):1037-1050.
[14]Chen W, Yang Y, Chen B, Lu P, Zhan L, Yu Q, Cao K, Li Q. MiR-136 targets E2F1 to reverse cisplatin chemosensitivity in glioma cells. J Neuro-Oncol, 2014,120(1):43-53.
[15]Qiu S, Huang D, Yin D, Li F, Li X, Kung HF, Peng Y. Suppression of tumorigenicity by microRNA-138 through inhibition of EZH2-CDK4/6-pRb-E2F1 signal loop in glioblastoma multiforme. Biochim Biophys Acta,2013,1832(10):1697-707.
[16]姚毅武, 姚和權, 蔣晟, 薛曉文. 組蛋白去乙酰化酶抑制劑抗腫瘤臨床研究進展. 中國新藥雜志, 2013,22(3):294-300.
[17]Gogada R, Yadav N, Liu J, Tang S, Zhang D, Schneider A, Seshadri A, Sun L, Aldaz CM, Tang DG, Chandra D. Bim, a proapoptotic protein, up-regulated via transcription factor E2F1-dependent mechanism, functions as a prosurvival molecule in cancer. J Biol Chem, 2013,288(1):368-381.
(本文編輯:洪悅民)
Effect of histone deacetylase inhibitor on the proliferation of glioma cells and expression of E2F1
Yuan Zhongmin,Liang Lin,Tang Xiaomei.
Department of Neurosurgery, the Second Affiliated Hospital of Guangzhou Medical University, Guangzhou 510260, China Corresponding author,Yuan Zhongmin,E-mail:yzm@gzhmu.edu.cn
ObjectiveTo investigate the proliferation effect of histone deacetylase inhibitors (HDACIs) on the proliferation of glioma cells and expression of E2F1. MethodsThe glioma cell line U251 were transfected with two plasmids of she2f1a or she2f1b, empty vector BS/U6 was regarded as a control, the cell proliferation rate was detected by tetrazoliumsalt (MTT) assays and nuclear staining method. U251 cells were treated with TSA for 2 h, 4 h and 8 h and treated with kinds of HDACIs including TAS, SAHA, M344, NaBu, VPA for 8 h. Dimethyl sulfoxide(DMSO) treatment was as control. Western blot was performed to test E2F1 expression. The glioma cell line U251 was treated with TSA for 8 h, 12 h and 24 h, DMSO treatment was regarded as control, and the cell proliferation rate was analyzed by MTT assays. U251 cells were treated with kinds of HDACIs including TSA, SAHA, M344, NaBu and VPA for 24 h, and then MTT assays were performed to determine cell proliferation rate. ResultsCompared with BS/U6, both she2f1a and she2f1b plasmids can silence E2F1 expression and inhibit cell proliferation (P<0.05). Compared with control, no significant inhibition of E2F1 expression was found in TSA treated cells at 2 h and 4 h, and the E2F1 expression was significantly inhibited at 8 h with TSA treatment, and the other HDACIs can significantly suppress E2F1 expression as well. MTT assays showed that TSA treatment 8 h samples did not remarkably affected cell proliferation rate (P>0.05), however, TSA treatment 12 h samples significantly inhibited cell proliferation. (P<0.05) and induced more suppression effects when lasting up to 24 h (P<0.05). Moreover, the other HDACIs can significantly inhibit cell proliferation (P<0.05). ConclusionHDACIs can suppress E2F1 expression of glioma cell and cell proliferation meditated by E2F1. Result
Glioma;E2F1;Histone deacetylase inhibitors

10.3969/j.issn.0253-9802.2016.08.003
廣東省自然科學基金(2015A030313475);廣州市科信局基金 (201510010180)
510260 廣州,廣州醫科大學附屬第二醫院神經外科(袁忠民,唐曉梅);510515 廣州,南方醫科大學第一臨床醫學院(梁琳)
,袁忠民,E-mail: yzm@gzhmu.edu.cn
2016-05-25)
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
