挽救生命的奇跡:DNA修復(fù)機(jī)制的發(fā)現(xiàn)及研究歷程
2016-09-05 18:10:25宋晶晶陳波
化學(xué)教學(xué) 2016年7期
宋晶晶 陳波
摘要:介紹了2015年的諾貝爾化學(xué)獎(jiǎng)DNA修復(fù)機(jī)制的研究歷程,包括其修復(fù)機(jī)理和修復(fù)途徑方面的理論與實(shí)踐研究以及DNA修復(fù)機(jī)制所展現(xiàn)的治癌新契機(jī),給生物醫(yī)學(xué)領(lǐng)域帶來(lái)的新的發(fā)展方向,為人類(lèi)的抗癌事業(yè)做出的巨大貢獻(xiàn)。
關(guān)鍵詞:諾貝爾化學(xué)獎(jiǎng);DNA修復(fù)機(jī)制;DNA修復(fù)途徑;抗癌
文章編號(hào):1005–6629(2016)7–0090–07 中圖分類(lèi)號(hào):G633.8 文獻(xiàn)標(biāo)識(shí)碼:B
2015年的諾貝爾化學(xué)獎(jiǎng)授予托馬斯·林達(dá)爾(瑞典)、保羅·莫德里奇(美)和阿奇茲·桑賈爾(土耳其)三位科學(xué)家,以表彰他們?cè)贒NA修復(fù)的細(xì)胞機(jī)制方面的研究。瑞典皇家科學(xué)學(xué)院在授予他們?cè)摌s譽(yù)的頒獎(jiǎng)詞中提到:三人在分子領(lǐng)域繪制出了細(xì)胞是如何完成DNA修復(fù)及保護(hù)其遺傳信息的,他們的研究發(fā)現(xiàn)為活細(xì)胞功能的認(rèn)知提供了基礎(chǔ)知識(shí),其研究成果在未來(lái)甚至可以為癌癥治療的發(fā)展提供極大的幫助。這一科學(xué)研究領(lǐng)域的成果有望為廣大癌癥患者帶來(lái)福音。那么,這一科學(xué)研究是如何被一步步發(fā)現(xiàn)的?又為何會(huì)獲得諾貝爾化學(xué)獎(jiǎng)這一崇高榮譽(yù)?其科學(xué)意義和應(yīng)用前景又會(huì)是怎樣?對(duì)我們又有何啟發(fā)?
1 DNA修復(fù)機(jī)制的發(fā)現(xiàn)
人們可能認(rèn)為DNA像是布滿灰塵的模板,DNA分子伴隨著人類(lèi)出生而出現(xiàn),死亡時(shí)便消失。我們知道DNA(即脫氧核糖核酸)是由核苷酸重新排列的一類(lèi)具有長(zhǎng)鍵的生物高分子,它是由兩條主鏈形成的雙螺旋結(jié)構(gòu)。在細(xì)胞核中,它是組成遺傳基因信息的一類(lèi)重要功能分子。然而DNA不僅如此,它還能夠真實(shí)地作為一個(gè)細(xì)胞內(nèi)部的計(jì)算機(jī),直接培育形成,并對(duì)細(xì)胞自身修復(fù)受損部位。簡(jiǎn)言之,這是維持人類(lèi)長(zhǎng)壽的關(guān)鍵因子,DNA有能力改變和修復(fù)人體受損組織,如癌癥和其他突變引起的損傷一樣能夠由DNA的自身修復(fù)功能來(lái)完成。
DNA是一種化學(xué)分子,必然在生物體內(nèi)會(huì)發(fā)生各種化學(xué)反應(yīng),則發(fā)生化學(xué)反應(yīng)的分子是如何修復(fù)的呢?基于此,科學(xué)家們一直對(duì)DNA的修復(fù)機(jī)制的研究充滿了興趣。
諾貝爾化學(xué)獎(jiǎng)的得主托馬斯·林達(dá)爾、保羅·莫德里奇和阿齊茲·桑賈爾各自獨(dú)立地闡明了與人類(lèi)相關(guān)的若干DNA修復(fù)過(guò)程。托馬斯·林達(dá)爾一直從事癌癥方面的研究,自然少不了對(duì)DNA修復(fù)機(jī)制的研究;保羅·莫德里奇早在杜克大學(xué)教授生物化學(xué)時(shí),就專注于研究DNA的錯(cuò)配修復(fù);而阿奇茲·桑賈爾專門(mén)從事DNA修復(fù)、細(xì)胞周期檢查點(diǎn)和生物鐘方面的研究,他花費(fèi)了較長(zhǎng)的時(shí)間研究光解和光激活的機(jī)制,直接觀察到了光解酶修復(fù)胸腺嘧啶二聚體的過(guò)程(即下面即將提到的直接修復(fù))。
“DNA到底有多穩(wěn)定?”,托馬斯·林達(dá)爾早在20世紀(jì)60年代末就對(duì)這個(gè)問(wèn)題深感好奇。那時(shí),科學(xué)界相信作為生命基礎(chǔ)的DNA分子必須極度堅(jiān)實(shí),沒(méi)有別的可能性。生物體要生長(zhǎng)必然要經(jīng)歷演化的過(guò)程,而生物演化需要突變的存在,但每一代的突變都是有限的。如果遺傳信息太過(guò)不穩(wěn)定,多細(xì)胞生物就無(wú)法存在了。在美國(guó)普林斯頓大學(xué)當(dāng)博士后的時(shí)候,林達(dá)爾研究RNA—一種與DNA相近的分子。研究很不順利。在實(shí)驗(yàn)中,他必須加熱RNA,但這個(gè)過(guò)程不可避免地會(huì)導(dǎo)致RNA快速降解。如果RNA在受熱時(shí)都被毀滅得如此迅速,具有相似性的DNA真的能一輩子保持穩(wěn)定嗎?
幾年后,林達(dá)爾進(jìn)入瑞典斯德哥爾摩的卡羅林斯卡學(xué)院。一些實(shí)驗(yàn)驗(yàn)證了他的懷疑:DNA其實(shí)會(huì)發(fā)生緩慢但可觀測(cè)的降解。據(jù)林達(dá)爾估計(jì),基因組每天會(huì)發(fā)生數(shù)千起災(zāi)難性的潛在損傷,人類(lèi)能在地球上得以延續(xù),這些損傷顯然不可能真的發(fā)生。他的結(jié)論是,肯定有分子機(jī)制負(fù)責(zé)對(duì)這些DNA缺陷進(jìn)行修復(fù)。本著這個(gè)思路,托馬斯·林達(dá)爾打開(kāi)了通向嶄新研究領(lǐng)域的大門(mén)。
和人的DNA一樣,細(xì)菌DNA同樣由核苷酸的四種堿基腺嘌呤(A)、鳥(niǎo)嘌呤(G)、胞嘧啶(C)和胸腺嘧啶(T)組成[1]。托馬斯·林達(dá)爾開(kāi)始利用細(xì)菌DNA來(lái)尋找修復(fù)酶。DNA的化學(xué)弱點(diǎn)之一,它的胞嘧啶(C)很容易丟失一個(gè)氨基,這可能導(dǎo)致遺傳信息改變。在DNA的雙螺旋中,C總是與G配對(duì),但當(dāng)氨基丟失后,受損的堿基往往與A配對(duì)[2]。因此,如果允許這個(gè)缺陷繼續(xù)存在,下一次DNA復(fù)制時(shí)就會(huì)發(fā)生一次突變。林達(dá)爾意識(shí)到,細(xì)胞對(duì)此必存在某種防范機(jī)制。隨后,他成功發(fā)現(xiàn),有一種細(xì)菌酶能去除DNA里受損的胞嘧啶。1974年,他發(fā)表了這一發(fā)現(xiàn)。
之后35年的成功工作就此展開(kāi),在此期間,托馬斯·林達(dá)爾發(fā)現(xiàn)并研究了許多細(xì)胞用來(lái)修復(fù)DNA所用的工具蛋白。在20世紀(jì)80年代初,他因緣際會(huì)來(lái)到英國(guó),在倫敦他接受了帝國(guó)癌癥研究基金會(huì)(Imperial Cancer Research Fund)的一個(gè)職位。1986年,他成為了新成立的劍橋大學(xué)克萊爾學(xué)堂實(shí)驗(yàn)室(Clare Hall Laboratory)主任,這個(gè)實(shí)驗(yàn)室隨后以其科學(xué)創(chuàng)造力而聞名遐邇。
一點(diǎn)一滴,林達(dá)爾拼起了“堿基切除修復(fù)(base excision repair)”是如何作用的分子圖景,證實(shí)了糖基化酶(glycosylases)正是DNA修復(fù)過(guò)程中的第一步,這種酶很類(lèi)似他在1974年發(fā)現(xiàn)的那個(gè)細(xì)菌的酶。堿基切除修復(fù)也發(fā)生在人類(lèi)身上,1996年,托馬斯·林達(dá)爾還在體外設(shè)法重建了人類(lèi)的修復(fù)過(guò)程。
對(duì)于托馬斯·林達(dá)爾而言,關(guān)鍵的一點(diǎn)是意識(shí)到了DNA不可避免地會(huì)發(fā)生變化,哪怕是當(dāng)分子位于細(xì)胞的保護(hù)性環(huán)境中時(shí)也不例外。于是,他完成了“堿基切除修復(fù)”的拼圖[4]。
但是,我們?cè)缫阎繢NA會(huì)因環(huán)境因素如UV輻射而受到損傷。林達(dá)爾并沒(méi)有發(fā)現(xiàn)并闡述這一受損過(guò)程是如何被修復(fù)的,而多數(shù)細(xì)胞用于修復(fù)紫外線傷害的機(jī)制“核苷酸切除修復(fù)”是被阿齊茲·桑賈爾闡明的。
桑賈爾出生于土耳其的薩武爾,活躍于美國(guó)研究界。他對(duì)于生命分子開(kāi)發(fā)的癡迷是在伊斯坦布爾攻讀醫(yī)學(xué)學(xué)位的時(shí)候培養(yǎng)起來(lái)的。畢業(yè)之后,他在土耳其的鄉(xiāng)下當(dāng)了幾年的外科醫(yī)師,但1973年時(shí),他決定學(xué)習(xí)生物化學(xué)。他的興趣是被一個(gè)特別的現(xiàn)象所激發(fā)的:細(xì)菌暴露在致命的紫外線照射下之后,如果再用可見(jiàn)藍(lán)光照射,它們突然就能死里逃生。桑賈爾對(duì)這近乎魔法的反應(yīng)感到非常好奇,這是一種怎樣的化學(xué)功能?
一位名叫克勞德·魯珀特(Claud Rupert)的美國(guó)人當(dāng)時(shí)在研究這一現(xiàn)象。于是阿齊茲·桑賈爾加入了他在達(dá)拉斯的德克薩斯州大學(xué)實(shí)驗(yàn)室。1976年,使用當(dāng)時(shí)的粗糙工具,他成功地克隆出能修復(fù)被紫外線損傷的DNA的酶——光解酶的基因,并成功讓細(xì)菌批量生產(chǎn)這種酶。這成為了他的博士論文,但在當(dāng)時(shí),并沒(méi)有引起很多反響。他申請(qǐng)了三次博士后職位,卻都遭到了駁回。對(duì)于光解酶的研究也被擱置了。為了繼續(xù)對(duì)DNA修復(fù)進(jìn)行研究,阿齊茲·桑賈爾在耶魯大學(xué)醫(yī)學(xué)院(這是業(yè)內(nèi)領(lǐng)先地位的機(jī)構(gòu))找了個(gè)實(shí)驗(yàn)室技術(shù)員的工作。在這里,他的工作最終讓他獲得了諾貝爾化學(xué)獎(jiǎng)。
當(dāng)時(shí),人們已經(jīng)知道細(xì)菌有兩套修復(fù)紫外線損傷的機(jī)制:一條系統(tǒng)是依賴光的作用的“光修復(fù)”,需要光解酶;另一個(gè)系統(tǒng)則可以在暗處發(fā)揮作用。阿齊茲·桑賈爾在耶魯大學(xué)的新同事們從二十世紀(jì)六十年代中葉就開(kāi)始研究暗修復(fù)系統(tǒng),研究對(duì)象是三個(gè)對(duì)紫外線敏感的細(xì)菌突變系,這三個(gè)細(xì)菌系中分別有不同的基因發(fā)生了突變,分別被稱為UVRa、UVRb與UVRc[5]。
就像此前對(duì)光修復(fù)的研究一樣,桑賈爾開(kāi)始探索暗修復(fù)的分子機(jī)制。只花了幾年的時(shí)間,他就鑒定、分離與描述了這三個(gè)基因編碼的酶。在突破性的體外實(shí)驗(yàn)中,他證明了這些酶可以發(fā)現(xiàn)紫外線傷害的位點(diǎn),然后在DNA鏈上切開(kāi)兩個(gè)切口,分別發(fā)生在紫外線損害位點(diǎn)兩側(cè)。一段12~13個(gè)堿基對(duì)的片段,包括損傷位點(diǎn),就這樣被切掉了。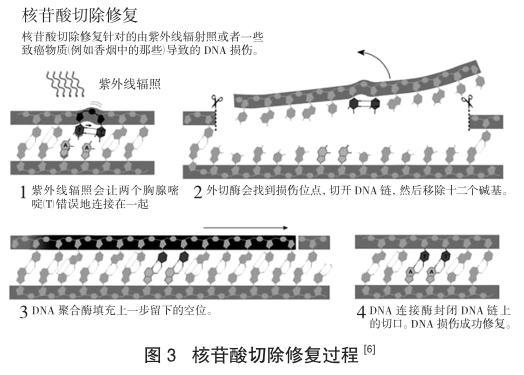
阿齊茲·桑賈爾從分子水平的細(xì)節(jié)中獲取對(duì)這一途徑認(rèn)知的能力改變了整個(gè)研究領(lǐng)域。他在1983年發(fā)表了他的研究。這樣的成就讓他獲得了北卡羅來(lái)納大學(xué)教堂山分校的副教授職位。在那里,他又一次漂亮地完成了對(duì)核苷酸切除修復(fù)機(jī)制的下一階段的研究工作。桑賈爾與其他研究人員——包括托馬斯·林達(dá)爾——同時(shí)進(jìn)行著人類(lèi)中的核苷酸切除修復(fù)的研究。在人體中,紫外線損傷修復(fù)的機(jī)制遠(yuǎn)比細(xì)菌中的程序復(fù)雜,但是從化學(xué)上來(lái)說(shuō),所有有機(jī)體里的核苷酸切除修復(fù)都是類(lèi)似的[7]。
然后,桑賈爾最初感興趣的光修復(fù)又怎樣了呢?最后他還是回到了光解酶,找到了它“復(fù)活”細(xì)菌的機(jī)制。另外,他還證明,人類(lèi)體內(nèi)相應(yīng)的酶幫助我們建立了生物鐘。
接下來(lái)就是保羅·莫德里奇的工作了。他也是從關(guān)于基因修復(fù)機(jī)制的模糊概念出發(fā),從中雕琢出了美妙的分子細(xì)節(jié)。
保羅·莫德里奇在美國(guó)新墨西哥州北部的一個(gè)小城鎮(zhèn)長(zhǎng)大。當(dāng)?shù)囟喾N多樣的廣袤風(fēng)景激發(fā)了他對(duì)大自然的興趣。但是有一天,他教生物的父親說(shuō):“你應(yīng)該研究一下DNA那套東西。”那是在1963年,詹姆斯·沃森和弗朗西斯·克里克因發(fā)現(xiàn)DNA結(jié)構(gòu)而獲得諾貝爾獎(jiǎng)的第二年。
幾年后,“DNA那套東西”真的成為了保羅·莫德里奇生活的中心。在他研究生涯的早期,在斯坦福大學(xué)做博士研究生、在哈佛大學(xué)做博士后以及在杜克大學(xué)做助理教授的時(shí)候,他檢驗(yàn)了一系列作用于DNA的酶:DNA連接酶,DNA聚合酶和限制性內(nèi)切酶。隨后,當(dāng)他在20世紀(jì)70年代末將注意力轉(zhuǎn)移到Dam甲基化酶時(shí)候,他不小心碰到了另一件“DNA那套東西”上,占據(jù)了他科學(xué)生涯的一大部分。
Dam甲基化酶能夠給DNA加上甲基基團(tuán)。保羅·莫德里奇證明,這些甲基基團(tuán)可以像路標(biāo)一樣起作用,幫助某個(gè)特定的限制性內(nèi)切酶在正確的位置切斷DNA鏈。但是,就在幾年前,馬修·梅塞爾森(Matthew Meselson),一位哈佛大學(xué)的分子生物學(xué)家,認(rèn)為DNA甲基基團(tuán)還有另一種信號(hào)作用。
利用一些分子生物學(xué)技巧,梅塞爾森改造出了一種噬菌體,它的DNA中有幾個(gè)不匹配位點(diǎn)。譬如,A(腺嘌呤)對(duì)面可能是C(胞嘧啶),而不是它本來(lái)應(yīng)該配對(duì)的T(胸腺嘧啶)。讓這些噬菌體感染細(xì)菌,細(xì)菌就會(huì)修復(fù)錯(cuò)誤的配對(duì)。沒(méi)人知道細(xì)菌為什么有這個(gè)功能,但是在1976年,梅塞爾森提出,這可能是因?yàn)榧?xì)菌自己本身存在的修復(fù)機(jī)制,來(lái)修復(fù)這些有時(shí)在DNA復(fù)制時(shí)出現(xiàn)的錯(cuò)誤。如果推測(cè)是正確的,梅塞爾森繼續(xù)推測(cè),可能DNA上的甲基基團(tuán)幫助細(xì)菌確認(rèn),糾正的時(shí)候哪個(gè)是原來(lái)的模板,哪個(gè)是后來(lái)的錯(cuò)誤。帶有錯(cuò)誤的新DNA鏈還沒(méi)有甲基化,可能這就是它被識(shí)別并修正的方式?
在DNA甲基化的過(guò)程中,保羅·莫德里奇和馬修·梅塞爾森的道路相交了。兩人合作構(gòu)建了一個(gè)具有一系列DNA錯(cuò)配的病毒。這一次,莫德里奇的Dam甲基化酶也被用于在其中一條DNA鏈上添加甲基。當(dāng)這些病毒感染細(xì)菌時(shí),細(xì)菌持續(xù)地修正那些未經(jīng)甲基化的、錯(cuò)配的DNA。莫德里奇和梅塞爾森的結(jié)論是,DNA的錯(cuò)配修復(fù)是一個(gè)天然的過(guò)程,在DNA復(fù)制過(guò)程中,以未甲基化為標(biāo)志來(lái)識(shí)別出錯(cuò)的DNA鏈。
保羅·莫德里奇的這個(gè)發(fā)現(xiàn)引發(fā)了長(zhǎng)達(dá)十年的系統(tǒng)性工作——克隆和測(cè)定錯(cuò)配修復(fù)過(guò)程中一個(gè)又一個(gè)酶。到20世紀(jì)80年代末,他已經(jīng)可以體外重建這套復(fù)雜的分子修復(fù)機(jī)理,并且深入了解它的細(xì)節(jié)。這項(xiàng)工作在1989年發(fā)表[8]。
和托馬斯·林達(dá)爾、阿齊茲·桑賈爾一樣,保羅·莫德里奇也研究了人類(lèi)版本的修復(fù)系統(tǒng)。今天我們知道,在人類(lèi)基因組復(fù)制時(shí)產(chǎn)生的所有錯(cuò)誤中,只有千分之一逃過(guò)了錯(cuò)配修復(fù)的法眼。但是,在人類(lèi)錯(cuò)配修復(fù)中,我們?nèi)匀徊荒艽_切地知道怎么判斷哪條鏈?zhǔn)窃镜逆湣NA甲基化在我們的基因組中有與微生物中不同的功能,所以,一定是什么別的東西在掌管究竟應(yīng)該修復(fù)哪條鏈——這個(gè)東西是什么還有待查明。
除了堿基切除修復(fù)、核苷酸切除修復(fù)和DNA錯(cuò)配修復(fù)這三種,還有其他多種機(jī)制維護(hù)著我們的DNA。每天,它們修復(fù)幾千起因?yàn)槿照铡⑽鼰熁蚱渌z傳毒性物質(zhì)導(dǎo)致的DNA損傷;它們不斷抵抗著DNA的自發(fā)改變。而且,每一次細(xì)胞分裂,錯(cuò)配修復(fù)都會(huì)糾正幾千個(gè)錯(cuò)配。沒(méi)有這些修復(fù)機(jī)制,我們的基因組將會(huì)崩潰。其中哪怕只有一個(gè)機(jī)制失靈了,遺傳信息就會(huì)很快改變,致癌風(fēng)險(xiǎn)也會(huì)增加。比如,DNA錯(cuò)配修復(fù)如果存在缺陷,就會(huì)增加患遺傳性結(jié)腸癌的風(fēng)險(xiǎn)。
事實(shí)上,在很多類(lèi)癌癥中,就是上述一到多個(gè)修復(fù)體系部分地或者全部地被關(guān)閉了。這使得癌細(xì)胞的DNA變得不穩(wěn)定,這也是癌細(xì)胞經(jīng)常突變且能夠抵抗化療的原因之一。
同時(shí),這些生病的細(xì)胞會(huì)更加依賴還在正常工作的修復(fù)體系:沒(méi)有剩下的這些修復(fù)體系,它們的DNA受到的損傷會(huì)過(guò)于嚴(yán)重,細(xì)胞也會(huì)死亡。研究者試圖利用這種弱點(diǎn)來(lái)研發(fā)新的抗癌藥物。通過(guò)抑制殘留的修復(fù)體系,人們可以減緩或者完全阻止癌癥的生長(zhǎng)。譬如,奧拉帕尼(olaparib)就是一種可以抑制修復(fù)體系的抗癌藥物。
我們?nèi)梭w的分子裝置一直受到周?chē)h(huán)境的攻擊。活性化學(xué)物質(zhì)使其腐敗,高溫使其折疊結(jié)構(gòu)改變而變性。紫外線則會(huì)使其分解,多數(shù)情況下,它們被損壞而無(wú)法正常地發(fā)揮作用。細(xì)胞可以無(wú)情地拋棄受損的蛋白質(zhì),卻承擔(dān)不起拋棄DNA的代價(jià)。DNA必須維持完美的構(gòu)型,因?yàn)樗?fù)載著遺傳信息,指導(dǎo)著細(xì)胞的生命進(jìn)程并遺傳給下一代,為確保這一遺傳信息不丟失,細(xì)胞具備了多種不同的方式保護(hù)DNA免于受損,并在其受損時(shí)修復(fù)它們[9]。
隨著研究的推進(jìn),我們對(duì)修復(fù)機(jī)制和修復(fù)過(guò)程有了更為深入系統(tǒng)的了解。在包括人類(lèi)在內(nèi)的哺乳動(dòng)物中,細(xì)胞內(nèi)不同的DNA受損主要通過(guò)六種方式修復(fù):①直接修復(fù)(direct repair,DR):直接修復(fù)DNA的烷基化損傷;②堿基切除修復(fù)(base excision repair,BER):主要修復(fù)內(nèi)源性的氧化、烷基化和脫氨基的堿基損傷;③核苷酸的切除修復(fù)(nucleotide excision repair,NER):主要修復(fù)外源性輻射、化學(xué)物質(zhì)及蛋白質(zhì)形成的DNA加成物;④錯(cuò)配修復(fù)(mismatch repair,MMR):修復(fù)自發(fā)產(chǎn)生的和堿基脫氨基、堿基氧化及堿基甲基化等導(dǎo)致的堿基錯(cuò)配;⑤同源重組修復(fù)(homologous recombination repair,HR)和⑥非同源末端連接(non-homologous end-joining repair,NHEJ):均修復(fù)DNA雙鏈斷裂。故而,要想研究細(xì)胞內(nèi)的DNA修復(fù)機(jī)制是如何運(yùn)行的,首先就要對(duì)DNA修復(fù)的六種途徑進(jìn)行分析[10]。
2 DNA修復(fù)途徑
細(xì)胞的DNA不斷遭受內(nèi)源性和外源性物質(zhì)的襲擊:內(nèi)源性物質(zhì)引起的損害包括正常代謝中產(chǎn)生的活性氧引起的氧化作用;來(lái)自環(huán)境的外源性損傷物質(zhì)包括化學(xué)物質(zhì)、致癌物質(zhì)、紫外線和化學(xué)治療藥物及電離輻射等。DNA損傷的類(lèi)型決定DNA修復(fù)的途徑,從圖5的脈絡(luò)中我們可以看到于DNA修復(fù)的六種途徑,以下我們主要介紹兩種。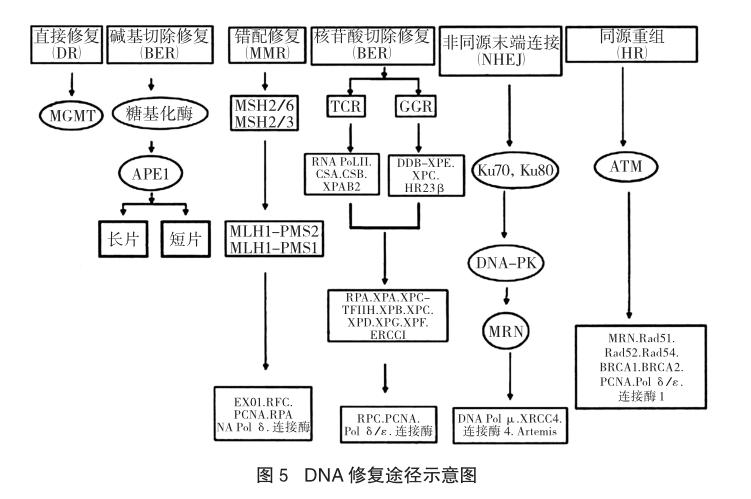
2.1 直接修復(fù)
有時(shí)稱為“直接逆轉(zhuǎn)”,該修復(fù)途徑是針對(duì)DNA由于外源性的損傷物質(zhì)(比如紫外線)而引起的損傷修復(fù)機(jī)制。通過(guò)微電子顯微鏡可以觀察到長(zhǎng)期受紫外線照射會(huì)使得DNA中相鄰的兩個(gè)嘧啶堿基通過(guò)共價(jià)鍵橋接起來(lái)造成損傷,這樣的雙嘧啶鍵結(jié)構(gòu)與DNA自身的雙螺旋結(jié)構(gòu)是不匹配的,如果不將其除去就無(wú)法完成遺傳因子的復(fù)制。細(xì)胞的直接修復(fù)可移除DNA分子中受損堿基的改變部位,而非切除受損堿基。所以,該途徑是通過(guò)DNA光解酶催化來(lái)完成的,將雙嘧啶鍵解分為兩個(gè)正常的單體,使DNA恢復(fù)原貌,恢復(fù)為正常的雙螺旋。
在可見(jiàn)光(300~600nm)活化之下,由光復(fù)活酶催化胸腺嘧啶二聚體分解為單體的修復(fù)機(jī)制如圖6顯示,這也是阿奇茲·桑賈爾研究發(fā)現(xiàn)的。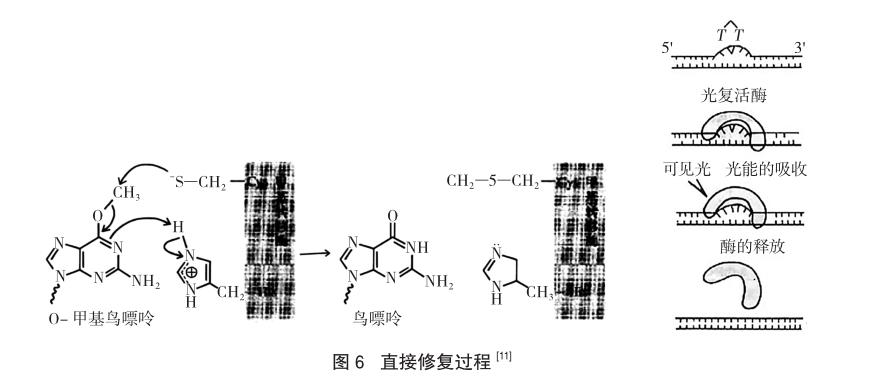
2.2 堿基切除修復(fù)(BER)
構(gòu)成DNA的腺嘌呤和胞嘧啶會(huì)發(fā)生自然的脫氨基化,生成次黃嘌呤和尿嘧啶,引起損傷。因此,堿基切除修復(fù)途徑主要是修復(fù)單個(gè)堿基受損的情況,包括:N-烷基化嘌呤;5-羥基胞嘧啶;尿素及其他加合物。DNA糖基化酶能識(shí)別,并將它們與糖苷鍵切斷,切除受損的堿基,脫氧核糖上留下無(wú)堿基位點(diǎn),啟動(dòng)修復(fù)過(guò)程。出現(xiàn)無(wú)堿基位點(diǎn)時(shí),利用核酸內(nèi)切酶將空位的磷酸鍵移除,形成空缺,最后通過(guò)DNA聚合酶和酶的作用完成修復(fù)過(guò)程[12]。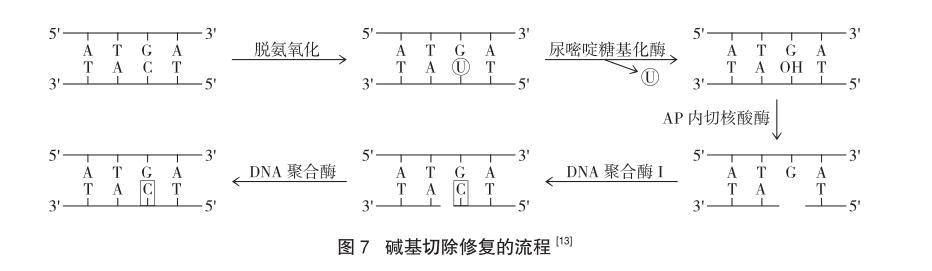
3 DNA修復(fù)機(jī)制的前景
隨著研究的深入,對(duì)于DNA修復(fù)機(jī)制的應(yīng)用也越來(lái)越廣泛。科學(xué)家用實(shí)際的探索向世界證實(shí)了:DNA修復(fù)并非空談。在了解了DNA修復(fù)機(jī)制的修復(fù)過(guò)程后,針對(duì)DNA修復(fù)系統(tǒng)進(jìn)行抗腫瘤藥物的研發(fā),近幾年來(lái)發(fā)展迅速。
細(xì)胞中這么多DNA,全都是從一開(kāi)始受精卵里那兩米長(zhǎng)的DNA復(fù)制來(lái)的。所有化學(xué)進(jìn)程都是不精確的,這幾十億次的復(fù)制之后早就應(yīng)該錯(cuò)得沒(méi)邊兒了,何況細(xì)胞還在每天承受活性分子和輻射帶來(lái)的損傷。但是我們大部分人都活得還挺好,基因也沒(méi)有變成一堆亂碼。我們體內(nèi)有一群蛋白質(zhì)專門(mén)負(fù)責(zé)看管DNA。它們持續(xù)不斷地校對(duì)基因組,發(fā)現(xiàn)損傷就立刻著手修復(fù)[14]。2015年的諾貝爾化學(xué)獎(jiǎng)表彰的就是發(fā)現(xiàn)這一修復(fù)機(jī)制的化學(xué)家。生命要想存活下去,遺傳物質(zhì)必須相對(duì)穩(wěn)定。最早時(shí)候,人們覺(jué)得DNA本身就特別結(jié)實(shí),根本就不需要修護(hù)。林達(dá)爾發(fā)現(xiàn),DNA其實(shí)沒(méi)有人想象的那么結(jié)實(shí),你要把DNA單獨(dú)拿出來(lái),它很容易壞,但是在人體內(nèi)卻沒(méi)壞,就證明它肯定是有什么特殊的機(jī)制在起作用。從他才開(kāi)始意識(shí)到我們需要這么一個(gè)機(jī)制。
事實(shí)上,許多類(lèi)型的癌癥就要?dú)w結(jié)于這些機(jī)制的失靈,但得是部分失靈。全部機(jī)制都完好的話,新的錯(cuò)誤就很難產(chǎn)生,癌癥就不容易發(fā)展;但如果所有機(jī)制都?jí)牧耍?xì)胞就承受不了錯(cuò)誤,會(huì)很快死掉。許多癌癥藥物都是以破壞癌細(xì)胞殘存修復(fù)機(jī)制為目標(biāo)的。因此,2015年諾貝爾化學(xué)獎(jiǎng)不但增進(jìn)了我們對(duì)細(xì)胞的了解,還可能成為許多拯救生命藥物的來(lái)源。
準(zhǔn)確地說(shuō),很多種癌癥發(fā)病的一個(gè)重要因素就是某些修復(fù)機(jī)制壞了,為什么會(huì)有癌癥?癌癥是因?yàn)檎5募?xì)胞出現(xiàn)了突變,或DNA出了錯(cuò),它不按照本來(lái)正常規(guī)定工作去運(yùn)行,修復(fù)機(jī)制完好的時(shí)候,它也不是萬(wàn)能的,之前說(shuō)一般它的修復(fù)率可能一千個(gè)里面會(huì)漏掉一個(gè),它也會(huì)犯錯(cuò)誤。但是在修復(fù)機(jī)制完好的時(shí)候,癌癥是比較難出現(xiàn)的,一旦眾多修復(fù)機(jī)制有一兩個(gè)壞掉了,這時(shí)候犯錯(cuò)誤的概率大大增加,即細(xì)胞失靈突變成癌細(xì)胞的概率大大增加了。修復(fù)機(jī)制也是隨機(jī)突變導(dǎo)致的,但是現(xiàn)在我們知道,它壞了可以想辦法修復(fù)它,或者是就反過(guò)來(lái)讓它壞得更徹底,讓癌細(xì)胞徹底死掉,這是對(duì)待癌癥思路之一。由此可見(jiàn),能夠找到適用的DNA修復(fù)機(jī)制就是找到了修復(fù)“生命密碼”,癌癥不再是無(wú)法戰(zhàn)勝的。
4 意義和啟發(fā)
2015年諾貝爾化學(xué)獎(jiǎng)的三位得主分別發(fā)現(xiàn)了三種DNA修復(fù)機(jī)制,這些基礎(chǔ)研究不僅加深了我們對(duì)于自身運(yùn)轉(zhuǎn)方式的理解,而且有助于繼續(xù)研發(fā)可以拯救生命的治療方法。如果我們回過(guò)頭來(lái)總結(jié)一下,就會(huì)發(fā)現(xiàn),他們的身上都具有一些相同的特質(zhì),這就是好奇心、不迷信權(quán)威和堅(jiān)持到底的精神。
與其同時(shí)代的研究人員都認(rèn)為DNA如此穩(wěn)定、如此強(qiáng)大,完全不需要任何修復(fù)。但托馬斯·林達(dá)爾在做關(guān)于RNA的分子實(shí)驗(yàn)時(shí),由于發(fā)現(xiàn)它特別不穩(wěn)定,很容易分解,卻能聯(lián)想到它的近親DNA是否也會(huì)具有相似的性質(zhì)?秉持著這份有依據(jù)的好奇心,他開(kāi)始了一次次的實(shí)驗(yàn),終于檢測(cè)到DNA也不像普遍認(rèn)為的那樣強(qiáng)大。而阿齊茲·桑賈爾的研究實(shí)際上也是從一個(gè)失敗開(kāi)始的,但他一旦決定了DNA的研究方向,就會(huì)對(duì)實(shí)驗(yàn)中的錯(cuò)誤和失敗展開(kāi)鍥而不舍的追問(wèn)和探究,并最終取得了豐碩的研究成果,也獲得了世人的肯定和激賞。
我們現(xiàn)在也已知,RNA和DNA的修復(fù)機(jī)制是多種多樣的,這也是科學(xué)家們?cè)谇叭搜芯康幕A(chǔ)上不斷摸索前行,開(kāi)拓新領(lǐng)域的結(jié)果。或許,諾貝爾獎(jiǎng)獲得者,不論其研究領(lǐng)域?qū)儆谀膫€(gè)方向,他們的科研經(jīng)歷和共通的品質(zhì)特性,或多或少,都會(huì)對(duì)我們的學(xué)習(xí)有所啟迪!
參考文獻(xiàn):
[1][3][12] 柴國(guó)林,朱衛(wèi)國(guó). DNA的損傷與修復(fù)[J].中華腫瘤雜志,2005,(10):577~580.
[2][5] P. Somnath, B. Nilanjana, C. Aditi, et al. Arsenicinduced Promoter Hypomethylation and Over-expression of ERCC2 Reduces DNA Repair Capacity in Humans by Nondisjunction of the ERCC2-Cdk7 Complex [J]. Metallomics,2014,(4):864~873.
[4] T. Lindahl, R. D. Wood. Quality Control by DNA repair [J]. Science,1999,(286):1897~1905.
[6][10][14] N. Shrivastav, D. Li, and J. M. Essigmann. Chemical Biology of Mutagenesis and DNA Repair: Cellular Responses to DNA Alkylation [J]. Carcinogenesis,2009,(31):59~70.
[7] Dragony Fu, Jennifer A, Calvo, Leona D, Samson. Balancing Repair and Tolerance of DNA Damage Caused by Alkylating Agents [J]. Nature Reviews Cancer, 2012,(12):104~120.
[8] A. Sancar, L. A. Lindsey-Boltz, K. Unsal-Kacmaz, S. Linn. Molecular Mechanisms of Mammalian DNA Repair the DNA Damage Checkpoints [J]. Annual Review of Biochemistry,2004,(73):39~85.
[9][美]古德賽爾著.王新國(guó)譯. 生命與死亡——《圖解生命》[M].中國(guó)青年出版社,2013:121~129.
[11] A. Sancar. Structure and Function of DNA Photolyase and Cryptochrome Blue-light Photoreceptors [J]. Chemical Reviews,2003,(103):2203~2237.
[13] P. Modrich, R. Lahue. Mismatch Repair in Replication Fidelity, Genetic Recombination and Cancer Biology[J]. Annual Review of Biochemistry,1996,(65):101~133.
