甲型副傷寒沙門氏菌cdtB亞基的原核表達及對巨噬細胞IL-6、IL-8、TNF-α分泌的影響
2016-08-25 01:51:09陳鴻鵠吳圓圓梅玲玲
中國人獸共患病學報 2016年6期
關鍵詞:信號
陳鴻鵠,吳圓圓,占 利,梅玲玲
?
甲型副傷寒沙門氏菌cdtB亞基的原核表達及對巨噬細胞IL-6、IL-8、TNF-α分泌的影響
陳鴻鵠1,吳圓圓2,占利1,梅玲玲1
目的研究甲型副傷寒沙門氏菌感染過程中,cdtB對宿主巨噬細胞分泌促炎細胞因子的影響。NF-κB信號通路阻斷劑對cdtB誘導的巨噬細胞分泌細胞因子的影響。方法對甲型副傷寒沙門氏菌cdtB亞基進行原核表達,制備并模型純化重組蛋白,建立其刺激人THP-1巨噬細胞模型,ELISA檢測THP-1分泌IL-6,IL-8和TNF-α等細胞因子。在共培養體系中加入NF-κB 信號通路阻斷劑,ELISA檢測THP-1分泌IL-6,IL-8和TNF-α等細胞因子。結果成功構建甲型副傷寒沙門氏菌cdtB原核表達系統,表達并純化重組cdtB蛋白,與空白對照相比,受到cdtB刺激的THP-1細胞上清中的IL-6,IL-8和TNF-α濃度顯著上升,而在THP-1細胞培養基中加入NF-κB信號通路阻斷劑SN50可以顯著抑制重組cdtB誘導的IL-6、IL-8、TNF-α分泌。結論甲型副傷寒沙門氏菌cdtB能夠通過NF-κB信號通路誘導巨噬細胞分泌IL-6、IL-8和TNF-α,在甲型副傷寒相關的炎癥反應中發揮促進作用。
甲型副傷寒沙門氏菌; cdtB;細胞因子;巨噬細胞
甲型副傷寒沙門氏菌(SalmonellaparatyphiA)引起的甲型副傷寒是常見的腸道傳染病,被列為國家重點監測的13種傳染病之一。近年來,傷寒和副傷寒的總發病率一直保持穩定,但是其中甲副傷寒的發病比例顯著增高[1-2]。 可見,甲型副傷寒沙門氏菌已替代傷寒沙門氏菌成為優勢致病菌。在感染急性期,傷寒和甲副傷寒病人血清中炎癥因子IL-6,IL-8和TNF-α顯著升高[3]。但是,甲型副傷寒沙門氏菌或甲型副傷寒沙門氏菌的組分如何導致機體產生炎癥的機理還有待闡明。
細胞致死性腫脹毒素(cytolethal distending toxin, CDT)是首先發現于空腸彎曲桿菌的一種外毒素,重組的空腸彎曲桿菌CDT具有類DNA酶的作用,能損傷巨噬細胞DNA,而使得細胞周期停滯,導致巨噬細胞腫脹、凋亡。CDT由cdtA、 cdtB和cdtC 3種多肽組成,其中,cdtB引起染色體DNA降解,cdtA和cdtC運送cdtB進入宿主細胞內。 傷寒沙門氏菌只表達cdtB,不表達cdtA和cdtC,由于傷寒沙門氏菌可以被巨噬細胞內吞到胞內,傷寒沙門氏菌可以直接在胞內致死宿主細胞[4]。伴放線桿菌CDT可以直接介導巨噬細胞釋放炎癥細胞因子[5]。巨噬細胞是病原菌引發的炎癥反應的核心,通過抗原提呈和釋放炎癥細胞因子激活宿主固有免疫系統,巨噬細胞在宿主控制和清除沙門氏菌感染的過程中發揮了重要作用[6]。甲型副傷寒沙門氏菌攜帶編碼cdtB的基因cdtB(GenBank accession No.: NC_006511.1),不攜帶cdtA和cdtC。本研究對甲型副傷寒沙門氏菌cdtB亞基進行原核表達,制備并模型純化了重組蛋白并建立其刺激人THP-1巨噬細胞模型。利用ELISA等檢測細胞因子的分泌水平,從細胞水平上研究甲副傷寒沙門氏菌cdtB亞基對巨噬細胞分泌IL-6,IL-8和TNF-α的影響。
1 材料與方法
1.1菌株來源和培養甲型副傷寒沙門菌參考標準株(CMCC 50001)由本實驗室保存,采用營養肉湯(Oxoid)培養。
1.2cdtB基因原核表達系統的構建及鑒定采用細菌基因組DNA制備試劑盒(Axygen)提取甲型副傷寒沙門菌CMCC 50001株基因組DNA,分光光度法測定其濃度。PCR擴增全長cdtB基因(GenBank accession No. NC_006511.1),引物序列如下:上游 5′-CGC CAT ATG (NdeI) AAAAAACCTGTTTTTTTCCT-3′,下游 5′-CGC CTC GAG (XhoI) ACAGCTTCGTGCCAAAAAGG-3′。反應條件為:94 ℃ 5 min,94 ℃ 30 s、52 ℃ 30 s、72 ℃ 90 s,30個循環;72 ℃ 10 min。采用1 μg/mL溴乙錠預染色的1.5%瓊脂糖凝膠電泳檢測擴增產物,采用T-A克隆試劑盒(TaKaRa)cdtB基因擴增產物克隆入pMDl9-T形成重組質粒pMDl9-TcdtB,委托上海Invitrogen公司測序。pMDl9-TcdtB和表達載體pET42a(Novagen)分別用NdeI(TaKaRa)和XhoI(TaKaRa)雙酶切,瓊脂糖電泳分離酶切片段后切膠回收目的片段。在T4 DNA連接酶(TaKaRa)作用下,將cdtB基因片段與線性化pET42a連接后轉化入表達宿主菌大腸桿菌BL21DE3 (Novagen)形成E.coliBL21DE3pET42a-cdtB表達重組cdtB。用10%分離膠的SDS-PAGE聯合Bio-Rad凝膠圖像分析系統檢查重組cdtB的表達情況,Ni-NTA親和層析柱(BioColor)提純重組cdtB。
1.3去除重組cdtB中的LPS為了避免大腸桿菌表達得到的重組cdtB中可能出現的大腸桿菌LPS污染,本研究通過Detoxi-Gel內毒素去除膠(Thermo Scientific, USA)去除重組cdtB中的LPS。為了檢測LPS是否完全去除,我們利用鱟試劑檢測重組cdtB,重組cdtB稀釋到不同濃度,各取100 μL加入等量鱟試劑,37 ℃孵育1 h。倒置試管,內容物凝固不能從管壁滑落為陽性結果;內容物從管壁滑落為陰性結果。本實驗中,無熱原水作為陰性對照,大腸桿菌LPS (Invivogen, USA)作為陽性對照。
1.4人單核分化巨噬細胞THP-1的分化與培養THP-1細胞接種于6孔培養板中(1×106細胞/孔,2 mL每孔),于37 ℃、 5%CO2條件下培養過夜使成單層細胞。THP-1細胞在實驗前加入終濃度為50 ng/mL佛波酯(PMA)刺激分化48 h,使之分化為巨噬細胞。
1.5重組cdtB蛋白刺激人單核分化巨噬細胞THP-1在經過PMA處理的THP-1(105/mL)的培養基中分別加入終濃度為0.1,1和10 μg/mL的重組cdtB,置于37 ℃、5% CO2培養箱中培養24 h后,收集上清,ELISA(ebioscience)檢測上清中IL-6、IL-8、TNF-α的濃度。本實驗中,不外加重組cdtB的THP-1細胞培養上清作為空白對照。
1.6NF-κB信號通路阻斷研究 為了研究NF-κB信號通路阻斷對重組cdtB誘導巨噬細胞IL-6、IL-8、TNF-α分泌的影響,20 μmol/L NF-κB信號通路阻斷劑SN50 (Tocris Bioscience, USA) 加入經過PMA預處理的THP-1細胞(2×10/孔)培養基中,1 h后用1 μg/mL重組cdtB刺激THP-1細胞,置于37 ℃、5% CO2培養箱中培養24 h后,收集上清,ELISA檢測培養上清中的IL-6、IL-8和TNF-α的濃度。本實驗中,不外加NF-κB信號通路阻斷劑SN50和重組cdtB的THP-1細胞培養上清作為空白對照,重組cdtB刺激的THP-1細胞培養上清作為陽性對照。
1.7統計學處理采用SPSS 18.0統計軟件對數據進行卡方檢驗。
2 結 果
2.1重組表達載體的構建和鑒定甲型副傷寒沙門菌CMCC 50001株基因組DNA中可以擴增出預期大小的cdtB基因片段(圖1)。測序及序列分析結果顯示,與GenBank中甲型副傷寒沙門菌ATCC26695株cdtB基因核苷酸和氨基酸序列相似性分別為97.62%和99.82%。
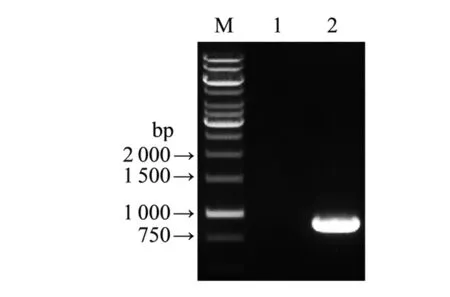
M: DNA marker; Lane 1: Blank control; Lane 2: Amplification fragments of cdtB gene from S.para CMCC 50001 strains.圖1 甲型副傷寒桿菌CMCC 50001株cdtB基因PCR結果Fig.1 PCR result of cdtB gene from S.para CMCC 50001 strains
2.2重組cdtB的表達及提純效果在IPTG誘導下,E.coliBL21DE3pET42a-cdtB能夠高效表達重組cdtB,該重組蛋白以可溶的形式存在。利用Ni-NTA親和層析法提純重組cdtB蛋白,SDS-PAGE結果顯示,重組cdtB蛋白在膠上表現為單一的蛋白條帶(圖2)。

M: protein marker; Lane 1: Blank control (wild-type pET42a). Lane 2 and 3: The expressed and purified SPA-rCdtB protein, respectively.圖2 甲型副傷寒桿菌CMCC 50001株重組CdtB表達和提純效果Fig.2 Effects of rCdtB expression and purification effects of S.para CMCC 50001 strains
2.3重組cdtB對巨噬細胞IL-6、IL-8、TNF-α分泌的影為了研究重組cdtB對巨噬細胞的促炎作用,我們用不同濃度的重組cdtB(0.1 μg/mL,1 μg/mL和10 μg/mL)刺激THP-1細胞。24 h后,ELISA檢測培養上清中的IL-6、IL-8和TNF-α。結果顯示,與沒有受到重組cdtB刺激的對照組相比,受到cdtB刺激的THP-1細胞上清中IL-6,IL-8和TNF-α濃度上升,并呈現劑量依賴效應(t=4.13,P<0.05)。不同刺激時間(h)對上清中IL-6,IL-8和TNF-α濃度影響的研究結果顯示,上清中的IL-6濃度在6~12 h間顯著上升(t=4.38,P<0.05),約在24~48 h時到達峰值。IL-8的分泌在2~6 h開始顯著升高(t=6.07,P<0.05),約在24~48 h時到達峰值。TNF-α的表達在2~6 h間顯著上升(t=3.78,P<0.05),約在12~24 h時到達峰值(圖3)。
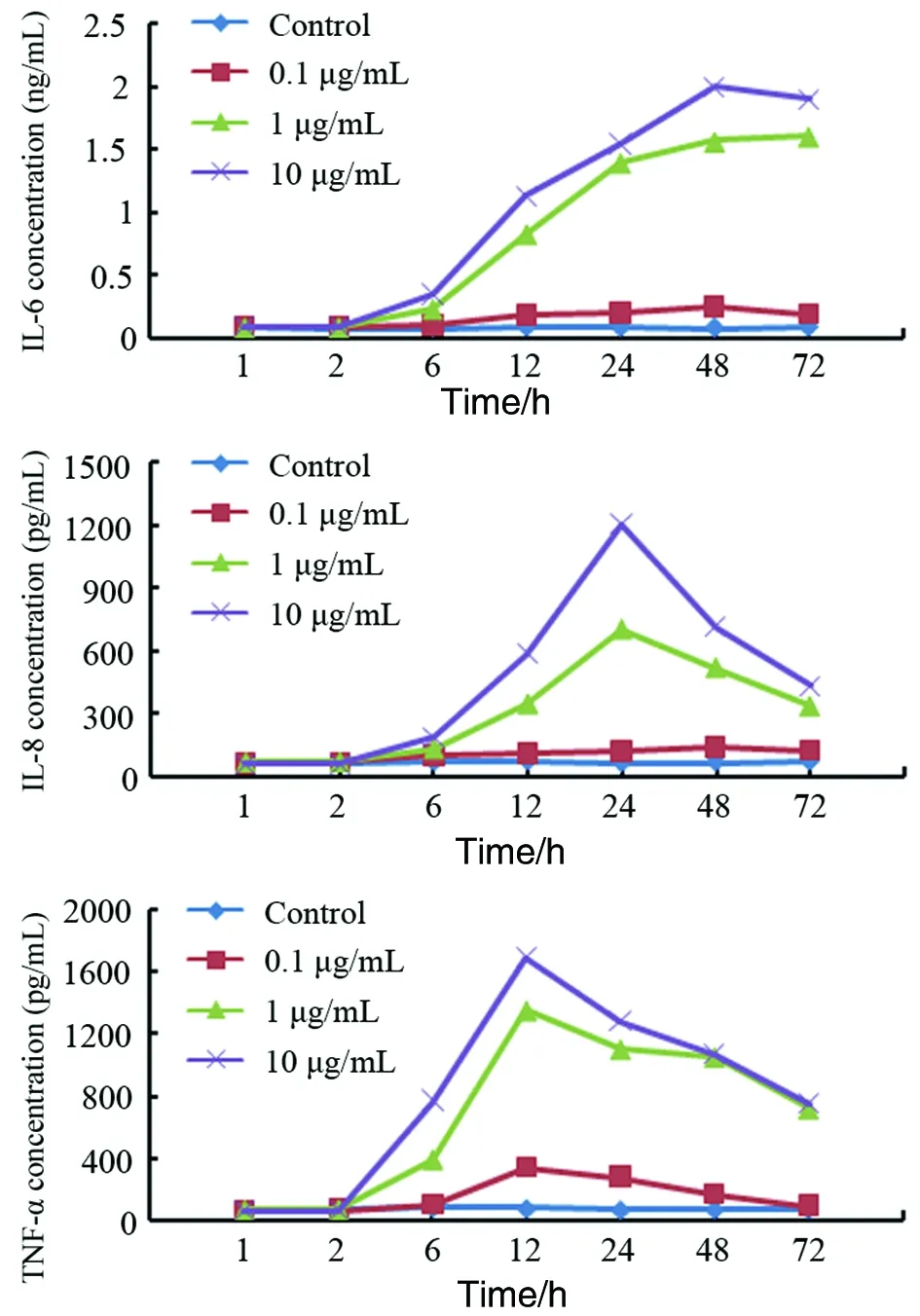
圖3 不同濃度重組CdtB對THP-1細胞因子分泌的影響Fig.3 Ability of rcdtB proteins to induce IL-6, IL-8 and TNF-α in THP-1 cells
2.4NF-κB信號通路阻斷對重組cdtB誘導巨噬細胞IL-6、IL-8、TNF-α分泌的影響在THP-1細胞培養基中加入NF-α信號通路阻斷劑SN50,1 h后用重組cdtB刺激THP-1細胞,24 h后,ELISA檢測培養上清中的IL-6、IL-8和TNF-α。結果顯示,與沒有加入NF-κB信號通路阻斷劑的陽性對照組相比,在THP-1細胞培養基中加入NF-κB信號通路阻斷劑SN50可以顯著抑制重組cdtB誘導的IL-6、IL-8、TNF-α分泌(t=2.41,P<0.05)(圖4)。
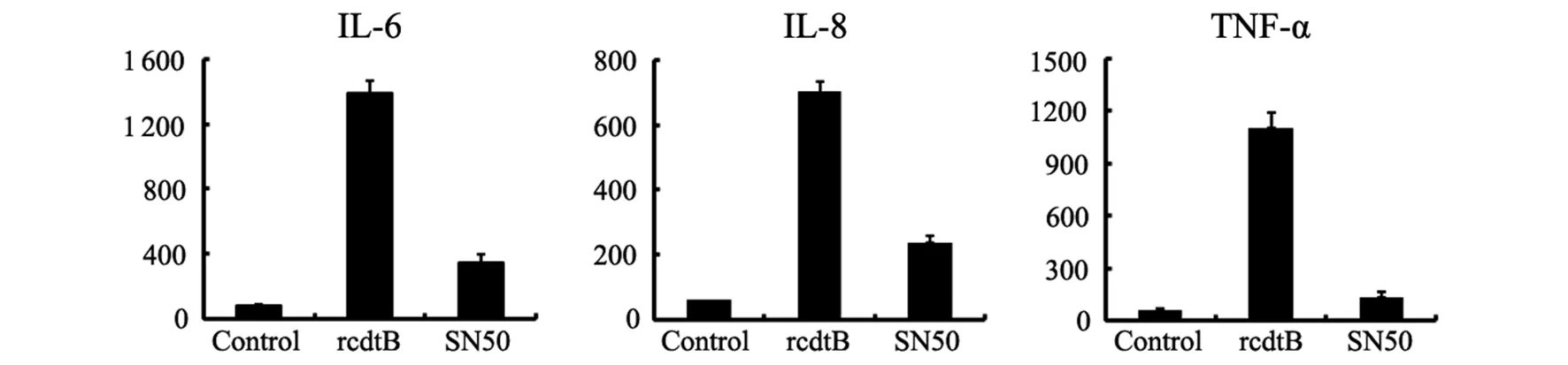
圖4 NF-κB信號通路阻斷對重組cdtB誘導THP-1分泌細胞因子的影響Fig.4 Influence of NF-κB signaling pathways inhibitor in rcdtB inducing IL-6, IL-8 and TNF-α
3 討 論
宿主對抗傷寒或甲型副傷寒沙門氏菌感染的特點是釋放促炎細胞因子,LPS被認為是傷寒和甲型副傷寒沙門氏菌的主要促炎組分。但是大多數傷寒病人血清中不能檢測到內毒素的存在,同時,傷寒沙門氏菌LPS耐受的動物模型在受到傷寒沙門氏菌攻擊時,還是表現傷寒熱的癥狀,說明傷寒和甲型副傷寒沙門氏菌LPS不是唯一促炎組分。傷寒和甲型副傷寒沙門氏菌的其他組分,如鞭毛蛋白,菌毛等也能誘導宿主細胞分泌促炎因子[7-8]。本研究發現,甲型副傷寒沙門氏菌重組cdtB能促使巨噬細胞分泌IL-6、IL-8和TNF-α,通過使用Detoxi-Gel內毒素去除膠處理重組cdtB,這些促炎細胞因子的釋放可以排除LPS污染的可能性。TNF-α是早期反應的促炎細胞因子,IL-6和IL-8是炎癥反應中重要的趨化因子,可以趨化和激活多種特異和非特異性的免疫細胞,從而激活宿主固有免疫系統[9-10]。這與以往的李鐵民報道的傷寒沙門氏菌能使人巨噬細胞分泌TNF-α的結果是一致的[11]。
NF-κB信號通路是調控促炎細胞因子基因表達的主要信號通路,NF-κB信號通路通過級聯信號放大和NF-rd3的表達,可以輔助促炎細胞因子的表達[12]。本研究通過NF-κB信號通路阻斷劑,證明甲型副傷寒沙門氏菌重組cdtB主要通過NF-κB信號通路誘導的巨噬細胞分泌IL-6、IL-8和TNF-α。但是,NF-κB信號通路阻斷劑不能完全阻斷甲型副傷寒沙門氏菌重組cdtB誘導的巨噬細胞分泌IL-6、IL-8和TNF-α,這結果說明,甲型副傷寒沙門氏菌重組cdtB可能還通過其他信號通路誘導巨噬細胞分泌促炎細胞因子。
我們的研究結果證明,甲型副傷寒沙門氏菌致死性腫脹毒素能夠通過NF-κB信號通路誘導人巨噬細胞分泌IL-6、IL-8和TNF-α,在甲型副傷寒相關的炎癥反應中發揮了促進作用。
[1] Wang HQ, Pan JC, Ge YM, et al. Molecular characteristics ofSalmonellatyphiandSalmonellaparaophia isolates in Hangzhou[J]. Chin J Microbiol Immunol, 2012, 32(8): 711-715. (in Chinese)
汪皓秋, 潘勁草, 葛玉梅, 等. 杭州地區傷寒及甲型副傷寒沙門菌流行菌株分子特征的研究[J]. 中華微生物和免疫學雜志, 2012,32(8):711-715.
[2] Yan MY, Liang WL, Li W, et al. Epidemiological analysis of typhoid and paratyphoid cases in China during 1995 to 2004[J]. Dis Ctrl, 2005, 20(8): 40l-403. (in Chinese)
閆梅英, 梁未麗, 李偉, 等. 1995—2004年全國傷寒副傷寒的流行分析[J]. 疾病監測, 2005, 20(8):401-403.
[3] Keuter M, Dharmana E, Gasem MH, et al. Patterns of proinflammatory cytokines and inhibitors during typhoid fever[J]. J Infect Dis, 1994, 169(6): 1306-1311. DOI:10.1093/infdis/169.6.1306
[4] Jinadasa RN, Bloom SE, Weiss RS, et al. Cytolethal distending toxin: a conserved bacterial genotoxin that blocks cell cycle progression, leading to apoptosis of a broad range of mammalian cell lineages[J]. Microbiology, 2011, 157(Pt 7): 1851-1875. DOI:10.1099/mic.0.049536-0
[5] Haghjoo E, Galan JE.Salmonellatyphiencodes a functional cytolethal distending toxin that is delivered into host cells by a bacterial-internalization pathway[J]. Proc Natl Acad Sci U S A, 2004, 101(13): 4614-4619. DOI:10.1073/pnas.0400932101
[6] Ando-Suguimoto ES, da Silva MP, Kawamoto D, et al. The cytolethal distending toxin ofAggregatibacteractinomycetemcomitansinhibits macrophage phagocytosis and subverts cytokine production[J]. Cytokine, 2014, 66(1): 46-53. DOI: 10.1016/j.cyto.2013.12.014
[7] Wyant TL, Tanner MK, Sztein MB.Salmonellatyphiflagella are potent inducers of proinflammatory cytokine secretion by human monocytes[J]. J Infect Dis, 1999, 67(7): 3619-3624.
[8] Wang FB, Zhang XL. Type IVB pili ofSalmonellatyphiinduce interleukin-6 expression in THP-1 cells by activating PKC signal pathway[J]. Immunol J, 2005, 21(3): 67-69. (in Chinese)
汪付兵, 章曉聯. 傷寒桿菌IVB型菌毛激活PKC信號通路誘導THP-1細胞IL-6表達 [J]. 免疫學雜志, 2005, 21(3):67-69.
[9] Amber KT, Bloom R, Mrowietz U, et al. TNF-alpha: a treatment target or cause of sarcoidosis[J]. J Eur Acad Dermatol, 2015, 29(11): 2104-2111. DOI: 10.1111/jdv.13246
[10] Nielsen AR, Pedersen BK. The biological roles of exercise-induced cytokines: IL-6, IL-8, and IL-15[J]. Appl Physiol Nutr Metab, 2007, 32(5): 833-839. DOI: 10.1139/H07-054
[11] Li TM, Xu H, Jiangqi XX. Research on mechanisms of mononuclear phagocyte in response toSalmonellatyphistrains[J]. Chin J Public Health, 2003, 19(1): 51-52. (in Chinese)
李鐵民, 徐浩, 江崎孝行.單核吞噬細胞應答傷寒沙門菌的實驗研究[J]. 中國公共衛生, 2003,19(1): 51-52
[12] Idzko M, Ferrari D, Eltizschig HK. Nucleotide signalling during inflammation[J]. Nature, 2014, 509(7500): 310-317. DOI: 10.1038/nature13085
Cloning and expression of recombinantSalmonellaparatyphiA cytolethal distending toxin proteins and its effect on cytokine production by human monocyte-derived macrophages
CHEN Hong-hu1, WU Yuan-yuan2, ZHAN Li1, MEI Ling-ling1
(1.DepartmentofMicrobiology,ZhejiangProvincialCenterforDiseaseControlandPrevention,Hangzhou310051,China;2.DepartmentofClinicalLaboratory,theChildren'sHospital,ZhejiangUniversitySchoolofMedicine,Hangzhou310003,China)
To investigate the role ofSalmonellaparatyphiA cytolethal distending toxin (CDT) in pro-inflammatory cytokines induction in macrophages, we cloned and expressed cdtB, a subunit of CDT fromSalmonellaparatyphi(S.paratyphi) A inEscherichiacoli, and purified the recombinant cdtB proteins (SPA-rcdtB) to homogeneity by Ni-NTA affinity chromatography. By co-cultured SPA-rcdtB with human monocyte-derived macrophages (THP-1) and detected IL-6, IL-8 and TNF-α in supernatant by ELISA, we determined if SPA-rcdtB were able to induce human monocyte-derived macrophages to produce cytokines. In addition, NF-κB inhibitor SN50 was used to research the effect of NF-κB signaling pathways in SPA-rcdtB induced macrophage IL-6, IL-8 and TNF-α secretion. In this study, we proved that the cdtb gene segment amplified fromS.paratyphiA CMCC 50001 strains showed high nucleotide and amino acid sequence identities (97.62% and 99.82%) compared withcdtbgene in the GenBank (GenBank accession No. NC_006511.1). And the SPA-rcdtB were able to induce the synthesis by THP-1 of interleukin-6, IL-8 and TNF-α, and the IL-6, IL-8 and TNF-α induction by SPA-rCdtB was significantly suppressed by NF-κB inhibitor. This result indicated that SPA-rcdtB plays a promoting role inflammatory response associated with SPA infection.
SalmonellaparatyphiA; cytolethal distending toxin; cytokines; macrophages.
Supported by the grants from the Zhejiang Provincial Center for Disease Control and Provention (No. 2013-11)
Mei Ling-ling, Email: llmei@cdc.zj.cn
梅玲玲,Email:llmei@cdc.zj.cn
1. 浙江省疾病預防控制中心微生物所,杭州310051;2. 浙江大學醫學院附屬兒童醫院實驗檢驗中心,杭州310003
R378.2
A
1002-2694(2016)06-0535-04
2015-11-13;
2016-03-30
DOI:10.3969/j.issn.1002-2694.2016.06.006
浙江省疾病預防控制中心青年科技項目(No.2013-11)資助
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
