不同培養條件對三角褐指藻生長及其生物活性成分積累的影響
2016-08-23 09:27:57張文源高保燕李愛芬張成武
海洋科學 2016年5期
關鍵詞:生長
張文源,高保燕,李愛芬,張成武
(暨南大學 水生生物研究中心生態學系,廣州 510632)
?
不同培養條件對三角褐指藻生長及其生物活性成分積累的影響
張文源,高保燕,李愛芬,張成武
(暨南大學 水生生物研究中心生態學系,廣州 510632)
以三角褐指藻(Phaeodactylum tricornutum)為研究材料,比較了NaNO3、NH4HCO3和CO(NH2)2為氮源的兩種培養基(mL1和ASW培養基)對其生長和生物活性成分(巖藻黃素、金藻昆布糖和二十碳五烯酸(C20:5,EPA))時相積累的影響,同時分析了脂肪酸組成和總脂含量的變化。結果表明:以mL1培養基培養時,三角褐指藻的生物質質量濃度明顯高于ASW培養基培養時的生物質質量質量濃度,尿素優于其他兩種氮源,最大生物質質量質量濃度為3.7 g/L。不同培養條件下巖藻黃素含量的時相變化規律一致,均隨著培養時間的延長呈現先增加后減少的趨勢,其最高積累量分別為:13.27 mg/g (NaNO3)、13.23 mg/g(CO(NH2)2)和 13.89 mg/g(NH4HCO3)(mL1);13.2 mg/g(NaNO3)、14.92 mg/g (CO(NH2)2)和13.6 mg/g(NH4HCO3)(ASW),由此可知氮源對巖藻黃素積累量影響不大。金藻昆布糖含量隨著培養時間延長逐漸增加,其最大積累量分別為9.82 mg/g(NH4HCO3)(mL1)和8.59 mg/g(NaNO3)(ASW)。不同培養條件下其總脂含量變化不顯著,均在培養末期達到最大值,分別為24.18%(NH4HCO3)(mL1)和23.79%(NaNO3)(ASW);其主要脂肪酸組成為:豆蔻酸(C14:0)、棕櫚酸(C16:0)、棕櫚油酸(C16:1)、硬脂酸(C18:0)、油酸(C18:1)、亞油酸(C18:2)、花生一烯酸(C20:1)、木焦油酸(C24:0)和EPA,其中,EPA含量隨著培養時間延長逐漸下降,尿素最有利于EPA的積累。
三角褐指藻;生物質質量濃度;巖藻黃素;金藻昆布糖;二十碳五烯酸
doi:10.11759/hykx20150706002
近年來微藻培養受到越來越多的關注,一方面是因為微藻來源豐富,生長迅速;另一方面是由于微藻具有生產高價值代謝產物的潛能,且能通過改變培養條件以提高細胞內生物活性物質的含量。其中海洋微藻的利用較為成熟,在保健食品、藥物、飼料、化妝品、生物農藥、廢水治理等方面具有較廣泛的應用前景[1]。而海洋微藻中種群數量最大的是硅藻,其作為海洋生態系統的初級生產者,生物多樣性高,生物量巨大,占海洋初級生產量的40%左右[2],是天然活性產物的潛在來源[3]。在海洋硅藻中,三角褐指藻的研究較為廣泛,被認為是巖藻黃素、金藻昆布糖和二十碳五烯酸(EPA)等生物活性物質的潛在生產者。另外,三角褐指藻可利用天然海水進行培養,不會競爭淡水資源,與其他硅藻相比三角褐指藻對硅的需求量較低,使其更易成為商業化生產的原料[2]。
巖藻黃素屬于類胡蘿卜素中的葉黃素類,是一種偏極性類胡蘿卜素,它可通過甲醇、乙醇及丙酮等有機溶劑提取,再經過硅膠柱層析的純化方法獲得,亦可通過CO2超臨界萃取的方法獲得。研究表明巖藻黃素具有抗炎、抗氧化、抗腫瘤、抗肥胖和抗糖尿病等多種生理活性[4]。目前已有不同類型的巖藻黃素營養保健品在市場中銷售,而天然的巖藻黃素主要來源于褐藻和硅藻。由于三角褐指藻能適應弱光強生長,其細胞色素組成中巖藻黃素所占的比例較高,因此,它被認為是天然巖藻黃素的最佳來源。
金藻昆布糖是一類水溶性多糖,主要由葡萄糖通過β-1,3糖苷鍵主鏈和少量的β-1,6糖苷鍵支鏈連接而成的葡聚糖[5]。研究表明金藻昆布糖是硅藻細胞中主要的儲存性碳水化合物,它能為細胞代謝循環提供能量,也能轉變成其他有機化合物[6],比如氨基酸、葡萄糖等。Kusaikin等[7]發現尖針桿藻(Synetra acus)中的金藻昆布糖具有明顯的抗腫瘤活性。Chan等[8]發現β-葡聚糖具有抗輻射、降血脂血糖等功效,常被稱作生物活性調節劑。
EPA屬于ω-3系列多不飽和脂肪酸,為人體必需脂肪酸。EPA已被證實具有多種生理功效,可預防冠狀動脈心血管疾病;可降低甘油三酯的含量,從而預防高甘油三酯血癥;可防止血小板凝集,降低血液中膽固醇含量,從而降低動脈硬化炎癥和各種瘤癥的風險[9]。EPA主要來源于魚油,但從魚油中獲取高純度高含量的EPA面臨一些問題,因為其EPA含量明顯低于10%,并混有其他多種脂肪酸[9]。而許多微藻富含EPA,其中三角褐指藻是EPA的一種潛在來源,并具有兩種優勢,一是三角褐指藻生長迅速,二是三角褐指藻細胞中二十二碳六烯酸(DHA)和花生四烯酸(AA)的含量相對較低,從而能大大增加EPA的回收率[10]。
因此,本文以海洋硅藻三角褐指藻為研究材料,比較以NaNO3、NH4HCO3和(NH2)2CO為氮源的兩種培養基對三角褐指藻生長及其細胞中巖藻黃素、金藻昆布糖和多不飽和脂肪酸積累的影響,以期為探求三角褐指藻的最適生長條件和生物活性成分積累的最佳條件提供依據。
1 材料和方法
1.1 實驗藻種與培養基
本實驗所用的三角褐指藻藻株來源于中國科學研究院海洋研究所,于暨南大學水生生物研究中心微藻生物資源技術實驗室保存。
1.2 培養條件與方法
對處于生長對數期的三角褐指藻進行接種培養。采用φ6 cm×60 cm的柱狀光生物反應器進行通氣培養,攪拌氣體為含1%二氧化碳的壓縮空氣,通氣速率為0.3 vvm,單側連續光照,光照強度為4000~4500 lx,培養溫度為25℃±1℃。初始接種密度控制在0.4~0.6。
以NaNO3、NH4HCO3和(NH2)2CO為氮源的mL1 和ASW的培養基進行培養,其中氮濃度設定為14.36 mmol/L,每種條件下設定3個平行,連續培養15天。每天從各管反應器中取10 mL藻液,用于生物量測定;每隔3天再取一定量的藻液,3500 r/min離心收集藻細胞,冷凍干燥后獲得凍干藻粉,儲存于-20℃的冰箱 中,用于各生化組分含量測定。
1.3 生物質質量濃度測定
采用重量法。混合纖維濾膜(上海市新亞凈化器件廠,孔徑0.45μm,尺寸50mm)于106℃電熱恒溫鼓風干燥箱(上海一恒科技有限公司)中干燥過夜至恒重M1,每天各管均勻取樣10mL藻液進行抽濾,并用0.5 mol/L的甲酸銨進行洗滌,測定抽濾后藻粉與濾膜總恒重值M2,則計算藻每天生物質質量濃度為(g/L)=1000×(M2-M1)/10。
1.4 巖藻黃素含量測定
采用有機溶劑提取法。取各條件下第0、3、6、9、12和15天的凍干藻粉,精確稱取10 mg,分別加入5 mL甲醇,于4℃黑暗條件下磁力攪拌提取12 h,3500 r/min離心5 min,取上清經0.45 μm微空有機濾膜過濾。過濾后所得色素提取物進行高效液相色譜分析,并與巖藻黃素標準曲線進行對照,計算出各時相巖藻黃素的含量。
1.5 金藻昆布糖含量測定
采用苯酚-硫酸法。稱取50 mg凍干藻粉,加入50 mmol/L的稀硫酸5 mL于60℃水浴提取1h。提取物離心后收集上清,提取后的藻渣再用蒸餾水洗滌2次,合并上清定容于50 mL容量瓶中。從容量瓶中吸取1 mL樣品,加入1 mL苯酚(6%)和5 mL濃硫酸充分搖勻,室溫靜置30 min后,于490 nm下測定吸光度值。結合葡萄糖的標準曲線,計算出金藻昆布糖含量。
1.6 脂肪酸組成分析
采用氣相色譜-質譜法(GC-MS)測定。精確稱取25 mg凍干藻粉,加入2 mL 2%的硫酸甲醇甲苯溶液(90︰10,v/v),同時加入十七烷酸作為內標,充氬氣后,置于80℃恒溫水浴鍋中攪拌1.5 h。反應結束后再在上述原位轉甲酯化樣品溶液中加入1 mL蒸餾水和1 mL正己烷,3500 r/min離心3 min,吸取上清至小瓶中,吹干后再加入200 μL正己烷密封,留存上樣,以測定脂肪酸組成及EPA的相對含量。
1.7 總脂含量的測定
總脂含量測定采用重量法。參考Khozi-Goldberg I等方法稍加修改。稱取80~100mg凍干藻粉,加入2mL二甲基亞砜-甲醇(1︰9,v/v)混合溶液,于50℃恒溫水浴中磁力攪拌1.5h,再在冰浴中磁力攪拌1.5 h,3000 r/min離心5 min,吸取上清至小玻璃瓶中。剩余藻渣再加入4 mL乙醚-正己烷溶液(1︰1,v/v),冰浴磁力攪拌1.5 h,3000 r/min離心5 min,轉移上清至上述對應小玻璃瓶中。以上操作重復一遍。在裝有上清液的小玻璃瓶中加入4 mL蒸餾水以靜置分層過夜。吸取上層相于離心管中,3000 r/min離心5 min,取上清于潔凈干燥小瓶中,吹干后用乙醚洗滌濃縮轉移至預先稱重的EP管中(管質量為m1),用氮氣吹至恒質量(m2)。得總脂含量(占藻粉干質量百分比,%)為:(m2-m1)/藻粉質量。
1.8 數據處理
運用Excel軟件進行平均值及標準差值的計算。運用origin 8.5軟件進行繪圖。
2 結果與分析
2.1 不同氮源的mL1和 ASW培養基對三角褐指藻生長的影響
為了測定不同氮源的mL1 and ASW培養基對三角褐指藻生長的影響,在同一種氮濃度(14.36 mmol/L)下,以生物質質量濃度為檢測指標。得到的生長曲線如圖1所示。由圖1可知,在3種形式氮源的mL1培養基和ASW培養基中,三角褐指藻的生物質質量濃度均逐漸增加,且尿素態氮和銨態氮的增長速率明顯優于硝態氮,最高生物質質量濃度均在第15天獲得,且均以尿素態氮條件下生物質質量濃度最高。mL1培養基條件下最高生物質質量濃度為3.7 g/L,ASW培養基條件下最高生物質質量濃度為3.3 g/L。
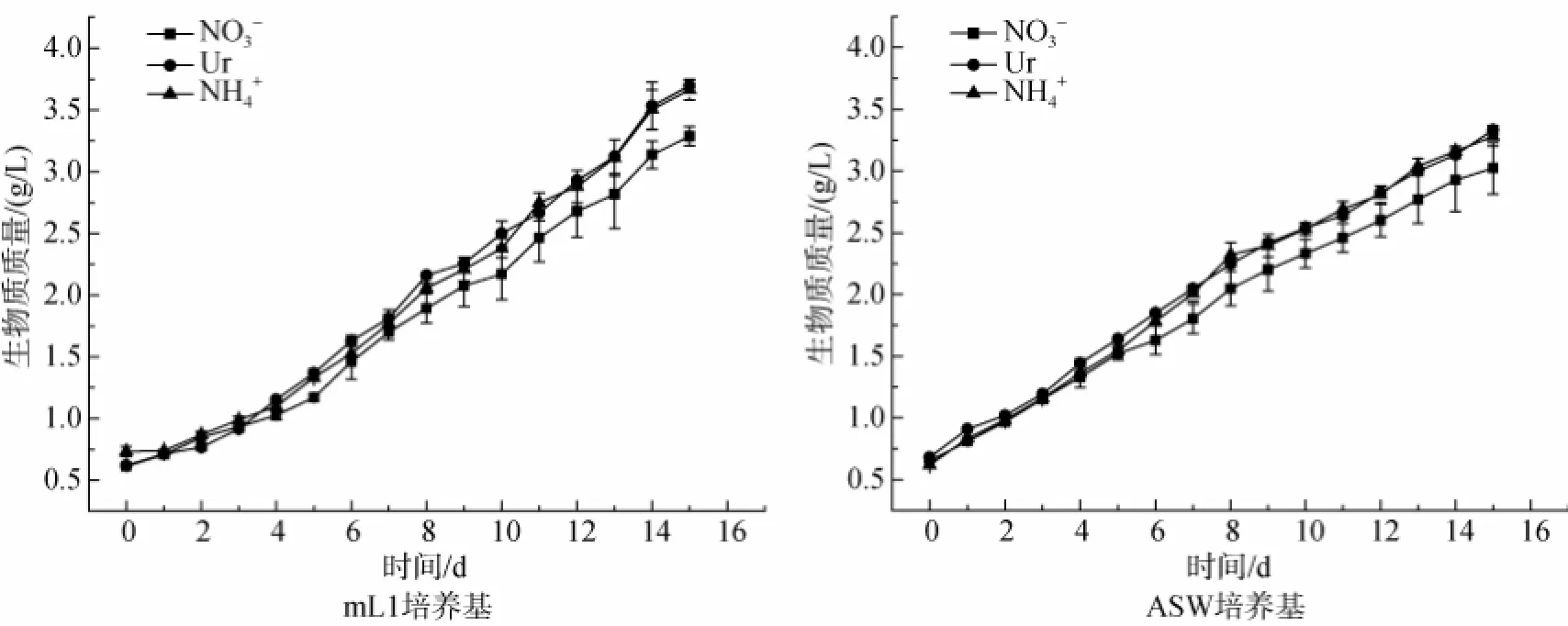
圖1 不同氮源的mL1和ASW培養基培養三角褐指藻時生物質質量濃度的時相變化Fig.1 Time-course changes in the biomass concentration of Phaeodactylum trirornutum grown in mL1 or ASW media with different nitrogen sources
2.2 不同氮源的mL1和 ASW培養基對三角褐指藻總脂積累的影響
圖2所示為三角褐指藻在不同氮源的mL1和ASW培養基培養時油脂積累的時相變化。由圖可知,油脂積累量隨著培養時間的延長呈現逐漸上升的趨勢,第15天達到最大值。在整個培養周期內,油脂積累速率均比較平緩,mL1培養基培養時油脂積累量略高于ASW培養基培養時油脂積累量,其油脂最高積累量分別為24.18%(mL1)和23.79%(ASW)。同一種培養基不同氮源條件下油脂積累規律基本相同,最大積累量相差不顯著。
2.3 不同氮源的mL1和ASW培養基對三角褐指藻巖藻黃素含量的影響
圖3和圖4分別顯示了三角褐指藻在mL1和ASW培養基培養時主要色素組成及巖藻黃素的時相變化。由圖可知,三角褐指藻中的主要色素組成有巖藻黃素、硅甲藻黃素、葉綠素a及β-胡蘿卜素等,其中巖藻黃素含量最高。在mL1培養基中,巖藻黃素含量隨著培養時間的延長呈現先增加后減少的趨勢,在第12天達到最大積累量,分別為:13.27 mg/g(NaNO3)、13.23 mg/g(CO(NH2)2)、13.89 mg/g(NH4HCO3)。在ASW培養基中,巖藻黃素呈現出相同的積累規律,在第9天達到最大積累量,分別為:13.2 mg/g(NaNO3)、14.92 mg/g(CO(NH2)2)、13.6 mg/g(NH4HCO3)。由此可知,兩種培養基對巖藻黃素的積累量影響不大,3種氮源中尿素氮源更有利于巖藻黃素的積累。

圖2 不同氮源的mL1和ASW培養基培養三角褐指藻時總脂含量的時相變化Fig.2 Time-course changes in the total lipid content of Phaeodactylum trirornutum grown in mL1 or ASW media with different nitrogen sources

圖3 不同氮源的mL1培養基培養三角褐指藻時巖藻黃素的時相變化Fig.3 Time-course changes in the pigment composition and fucoxanthin content of Phaeodactylum trirornutum grown in mL1 or ASA media with different nitrogen sources
2.4 不同氮源的mL1和 ASW培養基對三角褐指藻金藻昆布糖尿含量和單位體積產率的影響
金藻昆布糖是三角褐指藻中主要的儲藏性碳水化合物。如圖5所示,培養基類型對金藻昆布糖的積累有一定的影響,采用mL1培養基培養時金藻昆布糖的積累量更高,最高達到9.82%。3種氮源對金藻昆布糖的積累影響不顯著,mL1培養基中金藻昆布糖積累量分別為 8.25 mg/g(NaNO3)、9.47 mg/g (CO(NH2)2)、9.82 mg/g(NH4HCO3),ASW培養基中金藻昆布糖的積累量分別為:8.59 mg/g(NaNO3)、8.13 mg/g (CO(NH2)2)、7.73 mg/g(NH4HCO3)。
圖6比較了不同培養條件下金藻昆布糖的單位體積產率。在mL1培養基中,3種氮源的最高單位體積產率分別為:18.12 mg/(L·d),23.37 mg/(L·d)和23.97 mg/(L·d),在ASW培養基中,3種氮源的最高單位體積產率分別為17.3 mg/(L·d),18.06 mg/(L·d)和16.91 mg/(L·d)。

圖4 不同氮源的ASW培養基培養三角褐指藻時巖藻黃素的時相變化Fig.4 Time-course changes in the pigment composition and fucoxanthin content of Phaeodactylum trirornutum grown in 1 mL or ASA media with different nitrogen sources

圖5 不同氮源的mL1和ASW培養基培養三角褐指藻時金藻昆布糖含量的變化Fig.5 Time-course changes in the chrysolaminaran content of Phaeodactylum tricornutum grown in mL1 or ASW media with different nitrogen sources

圖6 不同氮源的mL1和ASW培養基培養三角褐指藻時金藻昆布糖的單位體積產率Fig.6 Volumetric production of chrysolaminaran by Phaeodactylum tricornutum grown in mL1 or ASW media with different nitrogen sources
2.5 不同氮源的mL1和 ASW培養基對三角褐指藻脂肪酸組成的影響
圖7所示為三角褐指藻在不同氮源的mL1和ASW培養基培養時脂肪酸的組成分析,由圖可知,6種培養條件下,三角褐指藻的脂肪酸組成基本相同,主要有豆蔻酸(C14:0)、棕櫚酸(C16:0)、棕櫚油酸(C16:1)、硬脂酸(C18:0)、油酸(C18:1)、亞油酸(C18:2)、花生一烯酸(C20:1)、木焦油酸(C24:0)和二十碳五烯酸(C20:5),且從第3天到第15天,C16:1、C18:0、C24:0含量增加,C18:1、C20:5含量減少,其他脂肪酸含量變化不顯著,表明這兩種培養基及3種不同氮源對脂肪酸組成的影響不大。

圖7 不同氮源的mL1和ASW培養基培養三角褐指藻的脂肪酸組成Fig.7 Fatty acid profiles of Phaeodactylum tricornutum on days 3 and 15 grown in mL1 or ASW media with different nitrogen sources
2.6 不同氮源的mL1和 ASW培養基對三角褐指藻二十碳五烯酸含量和單位體積產率的影響
圖8所示為三角褐指藻在兩種培養基3種不同氮源條件下培養時二十碳五烯酸(EPA)含量的變化。由圖8可知,二十碳五烯酸含量隨著培養時間延長而逐漸下降,以銨氮為氮源mL1培養基培養時二十碳五烯酸(EPA)含量下降的最為顯著,由22.14%下降到14.58%。而以尿素為氮源時,二十碳五烯酸(EPA)含量由19.97%下降至17.45%,因而最有利于EPA的積累。EPA占總脂肪酸的最大比例為22.14%。ASW培養基條件下,二十碳五烯酸(EPA)含量由19.58%(NaNO3)、20.78%(CO(NH2)2)、20.77% (NH4HCO3)下 降 至 16.75%(NaNO3)、17.44% (CO(NH2)2)、17.71%(NH4HCO3),EPA占總脂肪酸的最大比例為20.78%。

圖8 不同氮源的mL1和ASW培養基培養三角褐指藻時EPA含量的變化Fig.8 Time-course changes in the eicosapentaenoic acid(EPA)content of Phaeodactylum tricornutum grown in mL1 or ASW media with different nitrogen sources
3種氮源的mL1培養基培養三角褐指藻時,EPA的最大單位體積產率分別為:2.56 mg/(L·d)(NaNO3),2.31 mg/(L·d)(CO(NH2)2)和2.77 mg/(L·d)(NH4HCO3)。另外,3種氮源的ASW培養基培養三角褐指藻時,EPA的最大單位體積產率分別為:2.98 mg/(L·d)(NaNO3),3.09 mg/(L·d)(CO(NH2)2)和3.19 mg/(L·d)(NH4HCO3)。見圖9。
綜上所述,三角褐指藻分別在mL1和ASW培養基培養時,二十碳五烯酸(EPA)的含量變化規律基本相同,在以尿素為氮源時更有利于二十碳五烯酸(EPA)的積累。
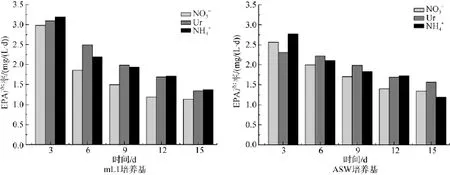
圖9 不同氮源的mL1和ASW培養基培養三角褐指藻時EPA的單位體積產率Fig.9 Volumetric production of eicosapentaenoic acid(EPA)by Phaeodactylum tricornutum grown in mL1 or ASW media with different nitrogen sources
3 討論
藻類的生長、油脂和色素的積累、多糖含量的變化及脂肪酸組成與含量均受各種環境因子和營養因子的影響,比如藻類的生長階段[11]、培養基的組成[12]、光照和溫度等,氮源類型對微藻的生長、油脂和色素的積累及多糖含量與脂肪酸組成也有著較為顯著的影響[13]。三角褐指藻在不同培養條件下的生長狀況已廣泛研究,但均只比較其中的某一種活性物質(多為二十碳五烯酸)的差異變化。本實驗比較了不同氮源、不同培養基對三角褐指藻的生長影響,并同時研究了其對巖藻黃素、金藻昆布糖和二十碳五烯酸3種活性物質含量變化的影響。
培養基提供了藻類生長所需的各種營養元素[14]。mL1培養基和ASW培養基均含有三角褐指藻生長所需的基本元素N、P、S、Si及Zn、Cu、Co、Mn等微量元素,但部分元素濃度存在差異。mL1培養基中,S的濃度是ASW培養基的10倍,NaCl的濃度相差不大,有研究報道NaCl濃度增加不會抑制三角褐指藻的生長,但會顯著影響脂肪酸的組成,當培養基中NaCl濃度增加時,EPA含量也隨之增加[15]。本實驗的研究結果顯示mL1培養基比ASW培養基更利于三角褐指藻的EPA積累。mL1培養基和ASW培養基均添加了硅藻生長所需的維生素,但Droop等[16]證實維生素對三角褐指藻中色素的積累沒有特別的影響。色素的含量隨著三角褐指藻的生長周期而發生變化[17]。mL1培養基和ASW培養基對巖藻黃素的積累量影響不大,且均隨著培養時間的延長呈現先增加后減少的趨勢,進一步印證前人的研究。
與mL1培養基相比,ASW培養基含有更少量的Si元素。但三角褐指藻在兩種培養基中均能生長,表明Si元素對三角褐指藻的生長不是必需的,但添加適量的Si元素能更有利于其生長。Andreas等[18]認為三角褐指藻雖是硅藻,但對Si元素沒有嚴格的需求,而Stigolsen等[19]發現Si元素對硅藻假微型海鏈藻影響較大,其能在淡水培養基生長,只要添加了Si元素。與mL1培養基相比,ASW培養基缺乏Se、Ni、Cr等微量金屬元素,表明這些微量元素對三角褐指藻的生長也有一定作用。
氮是微藻生長所必需的基本元素之一,是構成藻體內蛋白質、核酸及色素的重要元素,對藻類的生長發育有著重要的作用[20]。大量研究表明,微藻能夠利用多種不同形式的氮源供自身生長[21]。三角褐指藻能利用銨態氮、硝態氮、尿素態氮及其他無機氮源。Yongmanitchai等[15]研究NaNO3、CO(NH2)2和NH4Cl對三角褐指藻生長和脂肪酸組成的影響,得出以尿素為氮源進行培養時能得到最大生物量,且最利于二十碳五烯酸(EPA)的積累。Andreas等[18]以KNO3、CO(NH2)2和NH4Cl為氮源培養三角褐指藻,得出尿素最佳氮源。本實驗以NaNO3、CO(NH2)2和NH4HCO3為3種不同氮源,得出尿素為最適合三角褐指藻生長的氮源,這與多數研究結果相符。通常,微藻能夠利用氨氮進行生長,但隨著NH4+被利用,培養液的pH逐漸下降,從而抑制了微藻的生長[22]。而硝氮被藻體吸收后,轉變成NH4+,使培養液的pH增加,從而不利于微藻的生長。以尿素為氮源時,不會引起培養液pH的變動。另外從經濟角度,尿素氮源成本低,且1摩爾尿素可提供2摩爾的N原子。
綜上所述,(1)mL1培養基與ASW培養基相比,含有三角褐指藻培養所需的Si元素和Se、Ni、Cr、V等微量金屬元素,含有更高濃度的S元素,因而更利于三角褐指藻的生長及金藻昆布糖、二十碳五烯酸(EPA)的積累,但對總脂、巖藻黃素的積累影響不顯著。(2)尿素態氮與硝態氮、銨態氮相比,更利于三角褐指藻的生長、巖藻黃素和二十碳五烯酸的積累,因此尿素為最佳氮源。但氮源類型對金藻昆布糖的積累影響較小。
[1] 高亞輝.海洋微藻分類生態及生物活性物質研究[J].廈門大學學報:自然科學版,2001,40(2):566-573.
Gao Yahui.Studies on taxonomy,ecology and bioactive products of marine microalgae[J].Journal of Xiamen University:Natural Science,2001,40(2):566-573.
[2] Benavides A M S,Torzillo G,Kopecky J,et al.Productivity and biochemical composition of Phaeodactylum tricornutum (Bacillariophyceae)cultures grown outdoors in tubular photobioreactors and open ponds[J].Biomass and Bioenergy,2013,54:115-122.
[3] 周成旭,田甜,陳海敏,等.海洋微藻β-葡聚糖的生物活性分析[J].海洋與湖沼,2014,45(1):66-72.
Zhou Chengxu,Tian Tian,Chen Haimin,etal.Biatoactivities of β-glucan in six marine microalgae[J].Oceanologia et Limnologia Sinica,2014,45(1):66-72.
[4] Peng J,Yuan J P,Wu C F,et al.Fucoxanthin,a marine carotenoid present in brown seaweeds and diatoms:metabolism and bioactivities relevant to human health[J].Marine drugs,2011,9(10):1806-1828.
[5] Xia S,Wan L,Li A,et al.Effects of nutrients and light intensity on the growth and biochemical composition of a marine microalga Odontella aurita[J].Chinese Journal of Oceanology and Limnology,2013,31:1163-1173.
[6] Myklestad S M.Production,ChemicalStructure, Metabolism,and Biological Function of the(1→3)-Linked,β3-D-Glucans in Diatoms[J]. Biological oceanography,1989,6(3-4):313-326.
[7] Kusaikin M I,Ermakova S P,Shevchenko N M,et al.Structural characteristics and antitumor activity of a new chrysolaminaran from the diatom alga Synedra acus[J].Chemistry of natural compounds,2010,46(1):1-4.
[8] Chan G C,Chan W K,Sze D M.The effects of beta-glucan on human immune and cancer cells[J].J Hematol Oncol,2009,2(25):1-11.
[9] Giménez A G,González M J I,Medina A R,et al.Downstream processing and purification of eicosapentaenoic(20:5n-3)and arachidonic acids(20:4n-6)from the microalga Porphyridium cruentum[J].Bioseparation,1998,7(2):89-99.
[10]Yongmanitchai W,Ward O P.Screening of algae for potential alternative sources of eicosapentaenoic acid[J].Phytochemistry,1991,30(9):2963-2967.
[11]Huerlimann R,De N R,Heimann K.Growth,lipid content,productivity,and fatty acid composition of tropical microalgae for scale-up production[J].Biotechnology and bioengineering,2010,107(2):245-257.
[12]Valenzuela-Espinoza E,Millán-Nú?ez R,Nú?ez-Cebrero F.Protein,carbohydrate,lipid and chlorophyll a content in Isochrysis aff.galbana(clone T-Iso)cultured with a low costalternative to the f/2 medium[J].Aquacultural Engineering,2002,25(4):207-216.
[13]Huang C C,Hung J J,Peng S H,et al.Cultivation of a thermo-tolerant microalga in an outdoor photobioreactor:influences of CO2and nitrogen sourceson the accelerated growth[J].Bioresource technology,2012,112:228-233.
[14]Chisti Y.Biodiesel from microalgae[J].Biotechnology advances,2007,25(3):294-306.
[15]Yongmanitchai W,Ward O P.Growth of and omega-3 fatty acid production by Phaeodactylum tricornutum under differentculture conditions[J].Applied and environmental microbiology,1991,57(2):419-425.
[16]DroopM R.Optimum relativeandactualionic concentrations for growth of some euryhaline algae[J].Verh Int Verein Theor,Angew Limnol,1958,13:722-730.
[17]Carreto J I,Catoggio J A.Variations in pigment contents of the diatom Phaeodactylum tricornutum during growth[J].Marine biology,1976,36(2):105-112.
[18]Meiser A,Schmid-Staiger U,Tr?sch W.Optimization of eicosapentaenoic acid production byPhaeodactylum tricornutumin the flat panel airlift(FPA)reactor[J].Journal of applied phycology,2004,16(3):215-225.
[19]Olsen S,Paasche E.Variable kinetics of silicon-limited growth in Thalassiosira pseudonana(Bacillariophyceae)in response to changed chemical composition of the growth medium[J].British phycological journal,1986,21(2):183-190.
[20]胡章喜,安時,段舜山,等.不同氮原對布朗葡萄藻生長、總脂和總烴含量的影響[J].生態學報,2009,29(6):3288-3294.
Hu Zhangxi,An Min,Duan Shunshan,et al.Effects of nitrogen sources on the growth,contents of total lipids and total hydrocarbons of Botryococcus braunii[J].Acta Ecologica Sinica,2009,29(6):3288-3294.
[21]Liang Y,Beardall J,Heraud P.Effects of nitrogen source and UV radiation on the growth,chlorophyll fluorescence and fatty acid composition of Phaeodactylum tricornutum and Chaetoceros muelleri (Bacillariophyceae)[J].Journal of Photochemistry and Photobiology B:Biology,2006,82(3):161-172.
[22]蔣漢明,高坤山.氮源及其濃度對三角褐指藻生長和脂肪酸組成的影響[J].水生生物學報,2004,28(5):545-551.
Jiang Hanming,Gao Kunshan.Effects of nitrogen sources and concentrations on the growth and fatty acid composition of Phaeodactylum tricornutum[J].Acta Hydrobiologica Sinica,2004,28(5):545-551.
(本文編輯:梁德海)
Effects ofdifferentculture conditions on growth and accumulation ofbioactivecompoundsby Phaeodactylum tricornutum
ZHANG Wen-yuan,GAO Bao-yan,LI Ai-fen,ZHANG Cheng-wu
(Department of Ecology&Research Center for Hydrobiology,Jinan University,Guangzhou 510632,China)
Nov.26,2015
Phaeodactylum tricornutum;biomass concentration;fucoxanthin;chrysolaminaran;eicosapentaenoic acid
In this study,the effects of mL1 and ASW media containing NaNO3,CO(NH2)2,or NH4HCO3on the growth and accumulation of bioactive compounds[fucoxanthin,chrysolaminaran,and eicosapentaenoic acid(EPA)]by the marine diatom Phaeodactylum tricornutum were investigated;and the total lipid content and fatty acid composition were analyzed.The results showed that maximum biomass concentration in mL1 medium with urea as the nitrogen source was 3.7 g/L.Fucoxanthin content initially increased and then decreased during culture under different conditions,and the highest quantities that accumulated were 13.27 mg/g(NaNO3),13.23 mg/g(CO(NH2)2),and 13.89 mg/g(NH4HCO3)in mL1 medium and 13.2 mg/g(NaNO3),14.92 mg/g(CO(NH2)2),and 13.6 mg/g (NH4HCO3)in ASW medium,indicating that the nitrogen source had a weak effect on fucoxanthin accumulation.Chrysolaminaran content increased gradually during the culture period,and the highest contents were 9.82 mg/g (NH4HCO3)in mL1 and 8.59 mg/g(NaNO3)in ASW on day 15.The total lipid content changed slightly under the different conditions,and the maximum values were 24.18%(NH4HCO3)in mL1 and 23.79%(NaNO3)in ASW,which were observed during the stationary phase.The fatty acid profiles were myristic acid(C14:0),palmitic acid (C16:0),palmitoleic acid(C16:1),stearic acid(C18:0),oleic acid(C18:1),linoleic acid(C18:2),eicosaenoic acid(C20:1),tetracosanoic acid(C24:0),and EPA(C20:5).The EPA content decreased gradually as culture time was extended.Urea was the optimal nitrogen source to accumulate EPA.
Q4.3
A
1000-3096(2016)05-0057-09
2015-11-26;
2016-02-06
國家“863”項目(2013AA065805);國家自然科學基金項目(31170337);廣東省低碳專項(2011-051);珠海市科技重大項目(PB20041018);珠海市科技攻關項目(PC20081008)
[Foundation:NationalHigh Technology Research andDevelopment Program ofChina (863Program),No.2013AA065805;NaturalScience Foundation of China,No.31170337;the Special Program for Low-Carbon,Reform and Development Commission of Guangdong Province,NO.2011-051;Scientific and Technological Key Program of Zhuhai,No.PB20041018;Scientific and TechnologicalProjectZhuhai,No.PC20081008]
張文源(1990-),女,湖南邵陽人,碩士研究生,主要從事海洋微藻生物活性物質的研究,E-mail:13397085106@163.com;張成武,通信作者,男,教授,博士生導師,主要從事微藻生物資源技術方面的研究,E-mail:tzhangcw@jnu.edu.cn
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
