斑石鯛早期發(fā)育的異速生長模式
2016-08-23 09:27:55王雨福肖志忠劉清華翟介明龐尊方馬文輝馬道遠(yuǎn)徐世宏肖永雙
海洋科學(xué) 2016年5期
王雨福,肖志忠,劉清華,翟介明,龐尊方,馬文輝,馬道遠(yuǎn),徐世宏,肖永雙,李 軍
(1.中國科學(xué)院 海洋研究所實(shí)驗(yàn)海洋生物學(xué)重點(diǎn)實(shí)驗(yàn)室,山東 青島 266071;2.中國科學(xué)院大學(xué),北京100049;3.青島海洋科學(xué)與技術(shù)國家實(shí)驗(yàn)室 海洋生物學(xué)與生物技術(shù)功能實(shí)驗(yàn)室,山東 青島 266237;4.萊州明波水產(chǎn)有限公司,山東 煙臺 261418)
?
斑石鯛早期發(fā)育的異速生長模式
王雨福1,2,3,肖志忠1,3,劉清華1,3,翟介明4,龐尊方4,馬文輝4,馬道遠(yuǎn)1,3,徐世宏1,3,肖永雙1,3,李 軍1,3
(1.中國科學(xué)院 海洋研究所實(shí)驗(yàn)海洋生物學(xué)重點(diǎn)實(shí)驗(yàn)室,山東 青島 266071;2.中國科學(xué)院大學(xué),北京100049;3.青島海洋科學(xué)與技術(shù)國家實(shí)驗(yàn)室 海洋生物學(xué)與生物技術(shù)功能實(shí)驗(yàn)室,山東 青島 266237;4.萊州明波水產(chǎn)有限公司,山東 煙臺 261418)
為研究斑石鯛(Oplegnathus punctatus)早期發(fā)育階段的生長規(guī)律及重要器官的異速生長模式,作者運(yùn)用實(shí)驗(yàn)生態(tài)學(xué)方法,測定了斑石鯛仔稚幼魚(0~70日齡)全長和體質(zhì)量隨日齡的變化規(guī)律。結(jié)果顯示,全長、體質(zhì)量與日齡的關(guān)系均符合Cubic函數(shù),隨日齡的生長曲線表現(xiàn)為S型。斑石鯛全長和體質(zhì)量的生長可分為3個階段,不同階段的生長率存在顯著差異(P<0.05)。斑石鯛的頭長、眼徑、吻長、口裂、體高、尾鰭長等重要外部形態(tài)學(xué)指標(biāo)均存在異速生長規(guī)律,其生長拐點(diǎn)依次為35日齡(全長20.32 mm)、22日齡(全長13.17 mm)、29日齡(全長16.63 mm)、30日齡(全長15.50 mm)、32日齡(全長19.79 mm)、32日齡(全長19.79 mm)。相對于全長,頭長、吻長、口裂在拐點(diǎn)前后由正異速生長變?yōu)榈人偕L;眼徑、尾鰭由正異速生長轉(zhuǎn)為負(fù)異速生長;而體高在0~70日齡為正異速生長,但拐點(diǎn)之后已明顯變緩。研究表明斑石鯛通過異速生長,確保與其基本生存能力相關(guān)的功能器官優(yōu)先發(fā)育,以適應(yīng)復(fù)雜多變的生存環(huán)境,并相應(yīng)地提高了仔稚魚的存活率,對魚苗培育具有重要的指導(dǎo)意義。
斑石鯛(Oplegnathus punctatus);早期發(fā)育;功能器官;異速生長
doi:10.11759//hykx20141216005
異速生長(allometric growth)最早是由生物學(xué)家Huxley[1]等在研究招潮蟹(Uca pugnax)的鉗大小隨蟹身體變化時提出的,指生物體在成長過程中,其各部分器官相對于整體的生長速率表現(xiàn)出不同步的現(xiàn)象。異速生長律通常以冪函數(shù)Y=aXb表示,Y為被研究對象的屬性值(如待測器官的大小),X是生物的個體大小(通常以重量和長度來表示),a與b為常數(shù)[2-3],該函數(shù)式可以用來描述某器官大小相對于整體的變化情況[4]。
異速生長是生物界普遍存在的現(xiàn)象,在魚類早期生長階段,由于個體發(fā)育的生理、環(huán)境因素(水溫、光照、鹽度、餌料等)的影響,仔稚幼魚各器官(眼、口、體高、尾鰭等)的生長并不同步,如西伯利亞鱘(Acipenser baeri)[5], 施 氏 鱘 (Acipenserschrenckii)[6], 海 馬(Hippocam puskuda)[7],鮸(Miichthys miiuy)[8]和條石鯛(Oplegnathus fasciatus)[9]等。在早期發(fā)育過程中,仔稚幼魚的功能器官相對于整體的生長速度要快,直至相應(yīng)功能完善后,其生長才會減緩或等速于整體生長。從生態(tài)功能適應(yīng)性上講,異速生長確保不同發(fā)育階段中最重要的器官優(yōu)先生長[10],這提高了仔魚的成活率和種群的繁衍能力。因此,對魚類早期生長模式的研究可以了解其生活史,并人為創(chuàng)造相應(yīng)條件,促進(jìn)相應(yīng)器官的發(fā)育,降低死亡率和畸形率。
斑 石 鯛 (Oplegnathus punctatus),屬 鱸 形 目(Perciformes)、 石 鯛 科 (Oplegnathidae)、 石 鯛 屬(Oplegnathus),俗稱黑金鼓、斑鯛,是分布在中國、日本沿岸海域的溫帶魚類,是目前新興的水產(chǎn)品種。本實(shí)驗(yàn)對斑石鯛仔稚幼魚的異速生長情況進(jìn)行了較為系統(tǒng)的研究,探討了其早期的生長特性及生態(tài)學(xué)意義,并為斑石鯛規(guī)模化苗種繁育及養(yǎng)殖提供相應(yīng)的科學(xué)依據(jù)和基礎(chǔ)理論指導(dǎo)。
1 材料與方法
1.1 實(shí)驗(yàn)材料
本實(shí)驗(yàn)所用魚苗取自萊州明波水產(chǎn)有限公司。斑石鯛親魚經(jīng)人工誘導(dǎo)性腺發(fā)育成熟并自然產(chǎn)卵受精,收集懸浮受精卵后進(jìn)行孵化。孵出魚苗培育條件為水溫(22±1)℃,海水鹽度29±0.5,日換水1/2,持續(xù)充氣。以孵化當(dāng)天為 0日齡,3~17日齡仔魚投喂輪蟲(Brachionus plicatilis),15~32日齡增投鹵蟲(Artemia sinica),30日齡開始進(jìn)行飼料馴化。
1.2 實(shí)驗(yàn)方法
實(shí)驗(yàn)設(shè)3個平行組,取樣期間從0日齡開始,每天從各實(shí)驗(yàn)池隨機(jī)取樣10尾仔、稚幼魚,MS-222麻醉,用體視鏡(江南XTL-1型)觀察仔魚的早期形態(tài)發(fā)育,以光學(xué)顯微鏡(Olympus SZX16)和游標(biāo)卡尺(S·H)測定其全長、頭長、眼徑、口裂、吻長、體高、尾鰭長(mm),并用電子天平測定仔魚的體質(zhì)量(g)。
1.3 數(shù)據(jù)分析
用SPSS16.0軟件進(jìn)行數(shù)理統(tǒng)計,分析全長-日齡、體質(zhì)量-日齡以及各功能器官相對全長的生長關(guān)系,并以R2值最大的函數(shù)關(guān)系式作為最佳擬合生長模式。
異速生長以冪函數(shù)即異速生長方程y=axb表示,自變量x為全長,y為相應(yīng)的各器官長度,a為截距,b為異速生長指數(shù)。b>1,待測器官相對于全長為正異速生長;b<1,待測器官相對于全長成負(fù)異速生長;b=1待測器官與全長等速生長。異速生長模型中若存在不同的生長階段,以拐點(diǎn)分開,不同階段用不同的異速方程表示:y=a1xb1,y=a2xb2。對b1、b2是否存在顯著差異做t檢驗(yàn),并對b1、b2是否等于1做t檢驗(yàn)。用SPSS16.0軟件中的Nonlinear Regression過程,并結(jié)合OriginPro9.2軟件中的Nonlinear Curve Fit進(jìn)行分段擬合,以相關(guān)指數(shù)R2最大及殘差平方和最小作為曲線擬合標(biāo)準(zhǔn),求得拐點(diǎn)值。用SPSS16.0統(tǒng)計軟件進(jìn)行顯著性檢驗(yàn),P<0.05表示具有顯著性差異,描述統(tǒng)計量用平均值±標(biāo)準(zhǔn)差表示,并用Origin9.2軟件作圖。
2 結(jié)果
2.1 全長與體質(zhì)量的生長變化
2.1.1 全長與日齡的函數(shù)關(guān)系
以SPSS16.0軟件中的Curve Estimation對全長與日齡的關(guān)系進(jìn)行模型分析與參數(shù)估計(表1),選取R2值最大(0.994)的Cubic函數(shù)作為最佳模型,其關(guān)系式為Y=0.036X2-0.486X+5.932。

表1 全長與日齡的模型分析及參數(shù)估計Tab.1 Model summary and parameter estimates(total length and days)
斑石鯛初孵仔魚(0日齡)全長為3.49 mm±0.12 mm,經(jīng)過70 d的生長,其全長達(dá)到71.35 mm±3.02 mm。分析發(fā)現(xiàn),全長隨日齡的變化分為生長速率明顯不同的3個階段。如圖1所示,0~21日齡的全長生長速率為0.155 mm/d,21~35日齡的全長生長速率為0.794 mm/d,35~70日齡的全長生長率為1.455 mm/d,3個階段的全長生長率具有顯著差異(P<0.05)。
2.1.2 體質(zhì)量與日齡的函數(shù)關(guān)系
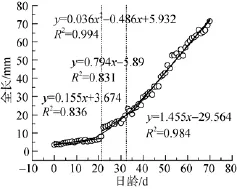
圖1 全長與日齡的最佳擬合曲線(Cubic)Fig.1 Best-fit curves between total length and days
以SPSS16.0軟件中的Curve Estimation對體質(zhì)量與日齡的關(guān)系進(jìn)行模型分析與參數(shù)估計(表2),選取R2值最大(0.994)的Cubic函數(shù)作為最佳模型,其關(guān)系式為Y=3.281×10-5X3-0.38X+0.006。
實(shí)驗(yàn)中,稱量了斑石鯛后期稚魚到幼魚的體質(zhì)量(31d~70 d),經(jīng)過70 d的生長,在70 d時體質(zhì)量達(dá)到8.01 g。體質(zhì)量隨日齡的變化分為生長速率顯著不同3個階段。如圖2所示,31~40日齡的體質(zhì)量生長率為0.038 g/d;40~50日齡的體質(zhì)量生長率為 0.145 g/d;50~70日齡的體質(zhì)量生長率為0.292 g/d,3個階段的體質(zhì)量生長率具有顯著性差異(P<0.05)。

表2 體質(zhì)量與日齡的模型分析及參數(shù)估計Tab.2 Model summary and parameter estimates(body weight and days)

圖2 體質(zhì)量與日齡的最佳擬合曲線(Cubic)Fig.2 Best-fit curves between body weight and days
2.2 各功能器官參數(shù)與全長的異速生長關(guān)系
如圖3a所示,全長為20.32 mm,35日齡時為頭長的生長的拐點(diǎn)。35日齡之前,b1=1.147,相對全長呈現(xiàn)正異速生長;35日齡之后,b2=1.028,與1差異不顯著(P>0.05),相對全長呈現(xiàn)等速生長。
如圖3b所示,全長為13.17mm,即22日齡時為眼徑的生長拐點(diǎn)。22日齡之前,b1=1.059,相對全長呈現(xiàn)正異速生長;22日齡之后,b2=0.846,相對全長呈現(xiàn)負(fù)異速生長。
如圖3c所示,全長為16.63mm,即29日齡時為吻長的生長拐點(diǎn)。29日齡之前b1=1.328,相對全長為正異速生長;29日齡之后,b2=0.997,與1差異不顯著(P>0.05),相對全長呈現(xiàn)出等速生長模式。
如圖3d所示,全長為15.50mm,即30日齡時為口裂的生長拐點(diǎn)。30日齡之前b1=1.062,相對全長為正異速生長;30日齡之后,b2=0.992,與1差異不顯著(P>0.05),相對全長呈現(xiàn)出等速生長模式。
如圖3e所示,全長為19.79mm,即32日齡時為體高的生長拐點(diǎn)。32日齡之前b1=1.508,為正異速生長;32日齡之后,b2=1.213,仍為正異速生長,但相對于32日齡之前已經(jīng)明顯變慢。
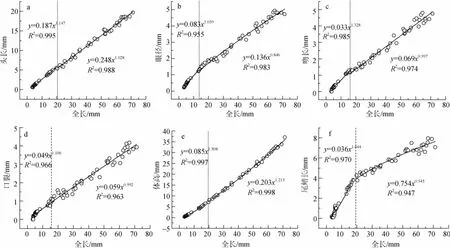
圖3 斑石鯛各功能器官與全長的異速生長關(guān)系Fig.3 Allometric growth of selected organs related to the total length of the spotted knifejaw
如圖3f所示,全長為19.79mm,即32日齡時為尾鰭的生長拐點(diǎn)。32日齡之前,b1=1.644,相對全長呈現(xiàn)正異速生長;32日齡之后,b2=0.545,相對全長呈現(xiàn)負(fù)異速生長。
3 討論
魚類在早期發(fā)育過程,饑餓和被捕食是影響仔稚魚成活率的重要原因[11-14]。因此,在長期進(jìn)化過程中,初孵仔魚營浮游生活之后,對生存起首要作用的攝食、消化及游泳器官進(jìn)入迅速生長階段。當(dāng)影響仔稚魚成活的重要器官功能指標(biāo)完善后,又開始轉(zhuǎn)為負(fù)異速或者等速生長階段[15-17]。在養(yǎng)殖過程中發(fā)現(xiàn),和其他硬骨魚類一樣,與斑石鯛攝食、游泳等密切相關(guān)的功能器官均表現(xiàn)出異速生長的模式。
3.1 全長與體質(zhì)量的生長變化
3.1.1 全長與日齡的生長關(guān)系
本實(shí)驗(yàn)中,斑石鯛仔稚魚全長與日齡的關(guān)系表現(xiàn)為Cubic函數(shù)形式,生長曲線為S型(圖1)。0~21日齡之間,由于仔稚魚主要以小輪蟲為營養(yǎng)來源,其視覺、游泳及攝食消化器官還未完善,因此維持在很低的生長速度0.155 mm/d。21~35日齡,影響斑石鯛生存的重要器官如口、視覺、鰭等已完成優(yōu)先發(fā)育,并處在功能逐步完善階段,捕食及游泳和躲避自殘的能力均大幅提高,故而生長速率提高到0.794 mm/d。35日齡之后,各項(xiàng)器官的功能指標(biāo)已完善,在33日齡左右完成飼料馴化,斑石鯛以較高的生長速度1.455 mm/d,進(jìn)入幼魚的快速生長階段。
3.1.2 體質(zhì)量與日齡的生長關(guān)系
本實(shí)驗(yàn)中,與全長類似,斑石鯛的體質(zhì)量隨日齡的變化也呈現(xiàn)出Cubic函數(shù)關(guān)系(圖2)。在30~40日齡之間,斑石鯛仔稚幼魚處于各功能器官的完善與適應(yīng)階段,體質(zhì)量增長很慢。40~50日齡,由于各器官已完成優(yōu)先發(fā)育,仔稚魚的捕食游泳能力大幅提高,體質(zhì)量開始增加明顯。50日齡之后,隨著飼料的追加和定期分苗,斑石鯛進(jìn)入了豐度增強(qiáng)期,體質(zhì)量快速增加。
3.2 各功能器官與全長的異速生長關(guān)系
3.2.1 頭長
斑石鯛仔稚魚的頭長相對于全長具有異速生長的特性,拐點(diǎn)為 35日齡,和條石鯛(Oplegnathus fasciatus)(38日齡)[9]類似,這是由于它們的外形比較接近產(chǎn)生的。頭部作為生物最重要的視覺、感覺信息的處理中心,也是攝食、呼吸器官賴以發(fā)育的基礎(chǔ),其結(jié)構(gòu)的完善與否對物種的生存能力有著極為重要的影響。斑石鯛頭部在短時間內(nèi)的快速生長,為眼、口、咽等器官的生長提供相應(yīng)的結(jié)構(gòu)基礎(chǔ)。
3.2.2 吻長、口裂
吻長和口裂是影響魚類攝食和吞咽食物顆粒的重要因素,在斑石鯛中它們的生長拐點(diǎn)比較接近,為29~30日齡,之后轉(zhuǎn)為等速生長。這和條石鯛(31日齡)[9]比較接近,但是比施氏鱘(Acipenser schrenckii)(8~9日齡)[6]、西伯利亞鱘(Acipenser baeri)(17~18日齡)[5]等要晚。由于西伯利亞鱘魚在14日齡左右,其口部結(jié)構(gòu)發(fā)育狀態(tài)已基本適應(yīng)人工飼料的投喂,而斑石鯛在32日齡之前主要以輪蟲、鹵蟲作為營養(yǎng)來源,30日齡左右開始進(jìn)行飼料馴化,故而斑石鯛的口部有著相對較長的生長期。口裂、吻長的生長拐點(diǎn)值為29~30日齡,確保在進(jìn)行投喂飼料之前,攝食器官已完備相應(yīng)的結(jié)構(gòu)和功能[18]。
3.2.3 體高
斑石鯛體高在70日齡之前,相對于全長均表現(xiàn)為正異速生長的模式,不同于條石鯛[9]的是32日齡之后雖然仍為正異速生長,但已經(jīng)顯著低于32日齡之前。這可能是種屬差異及寬扁的體型特征所決定的。此外,體高在較長時間段內(nèi)充分增加,為斑石鯛腹部內(nèi)臟器官的生長發(fā)育提供了結(jié)構(gòu)基礎(chǔ),也可能為背部肌肉的生長提供相應(yīng)的結(jié)構(gòu)基礎(chǔ)。
3.2.4 眼徑
自然生存狀態(tài)下,食物資源的競爭、躲避天敵及自殘的能力是影響仔稚魚成活的關(guān)鍵因素,而這些與視覺器官(眼睛)的發(fā)育狀況有重要的聯(lián)系。斑石鯛仔魚眼徑的生長拐點(diǎn)為22日齡,此后轉(zhuǎn)為負(fù)異速生長。眼徑的生長拐點(diǎn)類似于條石鯛[9]、鮸(Miichthys miiuy)[8]、鱸鯉(Percocypris pingi)[19],說明它們在早期發(fā)育過程中,對視覺器官的優(yōu)先程度是基本一致的。視覺器官結(jié)構(gòu)和功能在短時間內(nèi)的快速完善,對于仔稚魚迅速捕獲食物、爭奪飼料、躲避敵害有重要作用。
3.2.5 尾鰭
尾鰭是魚類早期發(fā)育階段最為重要的游泳器官,為幼魚游動提供前行動力和方向,對于魚類競爭食物資源,躲避種內(nèi)自殘等生存能力有重要影響作用[20]。斑石鯛尾鰭的生長拐點(diǎn)為32日齡,尾鰭的優(yōu)先發(fā)育時間要明顯長于條石鯛(15日齡)。這可能與條石鯛存在嚴(yán)重的自殘行為,故而要保持在較短(15日齡)時間內(nèi)實(shí)現(xiàn)尾鰭的迅速發(fā)育,以期躲避種群的自殘和對食物資源的快速爭奪。而斑石鯛溫和的特性,可以使得其運(yùn)動器官在較長時間內(nèi)(32日齡)實(shí)現(xiàn)發(fā)育。然而隨著鰾的充氣(16日齡左右)、臀鰭、腹鰭、胸鰭等游泳器官的完善,降低了對尾鰭的依賴程度,尾鰭的生長在后期明顯變慢。
通過對斑石鯛仔稚幼魚的早期生長發(fā)育研究,發(fā)現(xiàn)它與其他硬骨魚類似,多數(shù)器官也存在異速生長的現(xiàn)象。仔魚孵化后,影響仔稚幼魚存活的重要功能器官能在短時間內(nèi)優(yōu)先發(fā)育,是物種為適應(yīng)復(fù)雜多變的自然生存環(huán)境,提高種群成活率而長期進(jìn)化的結(jié)果。本研究表明,在苗種培育過程中,可以根據(jù)斑石鯛早期生長發(fā)育的特點(diǎn),人為創(chuàng)造有利條件,優(yōu)化飼料配比和投喂策略,確保魚苗的成活率。
[1] Huxley J S,Tessier G.Terminology of relative growth[J].Nature,1936,137:780-781.
[2] Peters R H.The ecological implications of body size[M].Cambridge:Cambridge University Press,1986.
[3] O'connell C P.Development of organ systems in the northern anchovy,Engraulis mordax,and other teleosts[J].American Zoology,1981,21(2):429-446.
[4] 韓文軒,方精云.冪指數(shù)異速生長機(jī)制模型綜述[J].植物生態(tài)學(xué)報,2008,32(4):951-960.
Han Wenxuan,F(xiàn)ang Jingyun.Review on the mechanism models of allometric scaling laws:3/4 vs.2/3 power[J].Journal of Plant Ecology,2008,32(4):951-960.
[5] 莊平,宋超,章龍珍,等.全人工繁殖西伯利亞鱘仔稚魚發(fā)育的異速生長[J].生態(tài)學(xué)雜志,2009,28(4):681-687.
Zhuang Ping,Song Chao,Zhang Lonzhen,et al.Allometric growth of artificial bred Siberian sturgeon Acipenser baerilarvae and juveniles[J].Chinese Journal of Econology,2009,28(4):681-687.
[6] 馬境,章龍珍,莊平,等.施氏鱘仔魚發(fā)育及異速生長模型[J].應(yīng)用生態(tài)學(xué)報,2007,18(12):2875-2882.
Ma Jing,Zhang Longzhen,Zhuang Ping,etal.Development and allometric growth patterns of larval Acipenser schrenckii[J].Chinese Journal of Applied Econology,2007,18(12):2875-2882.
[7] Choo C K,Liew H C.Morphological development and allometric growth patterns in the juvenile seahorse Hippocam puskudaBleeker[J].Journal of Fish Biology,2006,69(2):426-445.
[8] 單秀娟,竇碩增.鮸魚(Miichthys miiuy)仔、稚魚發(fā)育生長方式及其生態(tài)學(xué)意義[J].海洋與湖沼,2009,40(6):714-719.
Shan Xiujuan,Dou Shuozeng.Allometric growth of croakerMiichthys miiuylarvae and juveniles and its ecological implication[J].Qceanologia Et LimnologiaSinica,2009,40(6):714-719.
[9] 何滔,肖志忠,劉清華,等.條石鯛早期發(fā)育階段的生長模式[J].水產(chǎn)學(xué)報,2012,36(8):1242-1248.
He Tao,Xiao Zhizhong,Liu Qinghua,et al.Allometric growth in rock bream larvae(Oplegnathus fasciatus Temminck et Schlegel 1844)[J].Journal of Fisheries of China,2012,36(8):1242-1248.
[10]Rodriguez A,Gisbert E.Eye development and the role of vision duringSiberian sturgeonearly ontogeny[J].Journal of Applied Ichthyology,2002,18(4):280-285.
[11]Houde E D.Fish early life dynamics and recruitment variability[J].American Fisheries Society Symposium,1987,2:17-29.
[12]Bailey K M,Houde E D.Predation on eggs and larvae ofmarine fishesand the recruitmentproblem[J].Advances in Marine Biology,1989,25:1-83.
[13]Kamler E.Early Life History of fish:an energetics approach[M].London:Chapman and Hall,1992:267.
[14]Iguchi K,Mizuno N.Early starvation limits survival in amphidromous fishes[J].Journal of Fish Biology,1999,54(1):705-712.
[15]Sink G M J,Boogaart J G M,Osse J W M.Larval growth patterns in Cyprinus carpio and Clarias gariepinuswith attention to the fin fold[J].Journal of Fish Biology,1997,50(6):1339-1352.
[16]Herbing I H.Development of feeding structures in larval fish with different life histories:winter flounder and Atlantic cod[J].Journal of Fish Biology,2001,59(4):767-782.
[17]宋洪建,劉偉,王繼隆,等.大麻哈魚卵黃囊期仔魚異速生長及其生態(tài)學(xué)意義[J].水生生物學(xué)報,2013,37(2):329-335.
Song Hongjian,Liu Wei,Wang Jilong,et al.Allometric growth during yolk-sac larvae of chun salmon(Oncorhynchus keta)and consequent ecological significance[J].Acta Hydrobiologica Sinica,2013,37(2):329-335.
[18]Gisbert E.Early development and allometric growth patterns inSiberian sturgeon and their ecological significance[J].J Fish Biol,1999,54(4):852-862.
[19]何勇鳳,吳興兵,朱永久,等.鱸鯉仔魚的異速生長模式[J].動物學(xué)雜志,2013,48(1):8-15.
He Yongfeng,Wu Xingbing,Zhu Yongjiu,et al.Allometric growth pattern of Percocypris pingi larvae[J].Chinese Journal of Zoology,2013,48(1):8-15.
[20]吳水清,李加兒,區(qū)又君,等.斜帶石斑魚(♀)×鞍帶石斑魚(♂)雜交子代仔、稚魚的異速生長[J].中國水產(chǎn)科學(xué),2014,21(3):503-510.
Wu Shuiqing,Li Jiaer,Ou Youjun,et al.Allometric growth of hybrid grouper(Epinephelus coioides♀×E.lanceolatus♂)larvae and juveniles[J].Journal of Fishery Sciences of China,2014,21(3):503-510.
(本文編輯:譚雪靜)
Allometric growth pattern during early ontogeny of spotted knifejaw(Oplegnathus punctatus)
WANG Yu-fu1,2,3,XIAO Zhi-zhong1,3,LIU Qing-hua1,3,ZHAI Jie-ming4,PANG Zun-fang4, MAWen-hui4,MADao-yuan1,3,XU Shi-hong1,3,XIAO Yong-shuang1,3,LI Jun1,3
(1.Key Laboratory of Experimental Marine Biology,Institute of Oceanology,the Chinese Academy of Science,Qingdao 266071,China;2.University of Academy of Science,Beijing 100049,China;3.Laboratory for Marine Biology and Biotechnology,Qingdao National Laboratory for Marine Science and Technology,Qingdao 266237,China;4.Lai Zhou Ming Bo Aquatic Co.,Ltd.,Yantai 261418,China)
Dec.16,2014
Spotted knifejaw(Oplegnathus punctatus);early ontogeny;functional organs;allometry
Early ontogenesis of spotted knifejaw(Oplegnathus punctatus)was investigated using morphology and morphometrics to study the allometric growth pattern.Spotted knifejaw were reared at(24±0.3)℃ from hatching to 70 days post-hatching(d),and total length,body weight,and allometric growth of important organs were determined.Therefore,the increases in the total length and body weight were consistent with the cubic function and displayed an S-curve with three distinct stages.Allometric growth patterns were detected between different organs (head,mouth,eyes,and fins)and total length and featured inflection points.The growth characteristics of the head length(with an inflection point at 35 d),rostrum length(29 d),and mouth width(30 d)changed from positive allometry to isometry around the infection points,whereas eye diameter(22 d)and tail fin length(32 d)changed from positive to negative allometry.Body height(32 d)maintained positive allometric growth during 0-70 d,with a distinctly lower growth rate after the reflection point than that before.These results indicate that the spotted knifejaw develop the most important organs using an allometric strategy primarily to adjust themselves to the surroundings and achieve a higher survival rate.
Q954.3
A
1000-3096(2016)05-0043-06
2014-12-16;
2015-02-06
山東半島重要海產(chǎn)品良種生態(tài)養(yǎng)殖與高值化產(chǎn)業(yè)鏈?zhǔn)痉顿Y助項(xiàng)目(KFJ-EW-STS-060);山東省科技發(fā)展計劃資助項(xiàng)目(2014GHY115019);鰲山科技創(chuàng)新計劃(No.2015ASKJ02,2015ASKJ02-03-03)
[Foundation:Chinese Academy of Science and Technology Service Network Planning(No.KFJ-EW-STS-060);The Science Technology Program of Shandong Province(No.2014GHY115019);The Scientific and Technological Innovation Project Financially Supported by Qingdao Laboratory for Marine ScienceandTechnology(No.2015ASKJ02,2015ASKJ02-03-03)]
王雨福(1986-),男,山東臨沂人,碩士研究生;主要從事海水魚類遺傳發(fā)育研究;E-mail:wangyufu1989@163.com;李軍,通信作者,E-mail:junli@qdio.ac.cn;肖志忠,通信作者,E-mail:xzz@qdio.ac.cn
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
小讀者(2021年2期)2021-03-29 05:03:48
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
