基于穩定同位素分析不同蛋白源對大菱鲆生長的影響
2016-08-10 06:16:04李忠清夏斌張利民王際英李寶山孫永智3王明輝
水生生物學報 2016年4期
關鍵詞:生長
李忠清夏 斌張利民王際英李寶山孫永智, 3王明輝
(1. 上海海洋大學水產與生命學院, 上海 201306; 2. 山東省海洋資源與環境研究院, 煙臺 264006;3. 山東升索漁用飼料研究中心, 煙臺 265500)
基于穩定同位素分析不同蛋白源對大菱鲆生長的影響
李忠清1, 2夏 斌2張利民2王際英2李寶山2孫永智2, 3王明輝1, 2
(1. 上海海洋大學水產與生命學院, 上海 201306; 2. 山東省海洋資源與環境研究院, 煙臺 264006;3. 山東升索漁用飼料研究中心, 煙臺 265500)
實驗采用混合植物蛋白(玉米蛋白和大豆濃縮蛋白等氮1∶1混合)替代魚粉蛋白投喂大菱鲆, 通過測定不同飼料組和體組織中氮穩定同位素的值, 估算氮同位素的周轉率及不同蛋白源對大菱鲆生長的貢獻比例。實驗設計了4種配合飼料, 在等氮等能的基礎上, 以混合植物蛋白分別替代飼料中0、10%、30%和50%的魚粉蛋白, 即CS0、CS10、CS30和CS50。將初始體重為(57.72±0.25) g的大菱鲆分成4組, 每組設3個平行,實驗周期為56d。實驗開始后的7d、14d、28d、42d和56d采集并測定大菱鲆肌肉和肝臟中氮穩定同位素值(δ15N)。研究結果表明, 當替代量達到50%時, 大菱鲆攝食率和特定生長率顯著低于其他組(P<0.05), 而飼料系數顯著高于其他組(P<0.05)。不同實驗組間氮同位素的周轉率均存在顯著性差異(P<0.05), 對照組的周轉速度最快。各實驗組肝臟的周轉率均顯著快于肌肉(P<0.05)。大菱鲆肌肉中氮同位素的周轉半衰期(23.96—32.42d)顯著高于肝臟的(12.38—16.83d) (P<0.05)。生長作用對大菱鲆肌肉氮同位素周轉的貢獻比例為57.33%—73.33%, 而對肝臟的貢獻比例為29.17%—36.10%。基于時間的周轉模型估算氮同位素分餾系數(Δ)為1.83‰— 2.83‰。采用Isosource軟件計算顯示, CS10、CS30和CS50組中魚粉、玉米蛋白和SPC三種蛋白源對大菱鲆生長的貢獻比例分別為80%、7%和13%, 62%、11%和27%, 46%、13%和41%。研究結果表明:生長作用和代謝作用共同驅動了氮穩定同位素在大菱鲆體組織中的周轉代謝; 氮穩定同位素在肝臟組織中的周轉速度明顯快于肌肉的, 相應的周轉半衰期更短; 各實驗組中SPC對大菱鲆生長的貢獻均優于玉米蛋白。
大菱鲆; 氮穩定同位素; 生長; 代謝; 周轉模型
長期以來, 蛋白質營養一直是國內外動物營養學研究領域的重點和熱點之一[1]。蛋白質周轉作為機體營養代謝的直接反應, 對調節蛋白質在組織中的沉積, 去除細胞內的異常蛋白, 維持細胞內酶和結構蛋白的穩定狀態以及細胞內環境的相對穩定具有重要作用, 是形成動物產品的唯一生物學途徑和動物適應生存環境的基本生物學機制[2]。目前,定量揭示機體蛋白質周轉代謝規律, 從代謝水平上探求蛋白源替代及蛋白質節約效應已經成為水生動物營養學研究中的重要課題。
隨著穩定同位素技術及相應質譜檢測技術的發展, 在非添加標記物的情況下基于穩定同位素自然豐度差異及富集變遷的示蹤, 為機體蛋白質周轉代謝機理的研究提供了新方法, 并逐漸向特定組織器官和細胞分子水平發展, 其具有干擾因素少、靈敏度高及結果可靠等優點, 且能反應機體長時間蛋白質周轉代謝的變動情況。Gamboa-Delgdo等[3, 4]利用13C和15N的天然豐度并基于時間的周轉模型,研究了不同蛋白源在凡納濱對蝦(Litopenaeus vannamei)中的周轉代謝率及對其生長的貢獻比例。Gianini-Abimorad等[5]通過平衡飼料中的賴氨酸和蛋氨酸含量, 利用穩定同位素技術分析了細鱗鯧(Piaractus mesopotamicus)幼魚肌肉蛋白質的周轉代謝率和氨基酸利用效率。
大菱鲆(Scophthatmus maximus L.) 屬鰈形目(Pleuronectiformes), 鲆科(Bothidae), 菱鲆屬(Psetta),俗稱歐洲比目魚, 在中國稱“多寶魚”、“瘤棘鲆”,現已成為我國重要的海水養殖品種[6, 7]。然而, 魚粉資源的匱乏和價格不斷飆升, 大大增加了其養殖成本[8—10]。因此, 尋求其他廉價、高效的蛋白源來替代魚粉以應對日益發展的水產養殖業已勢在必行[11, 12]。本實驗采用玉米蛋白和大豆濃縮蛋白(Soy protein concentrate, SPC)按不同比例替代大菱鲆飼料中的魚粉, 通過測定肌肉和肝臟中氮穩定同位素值的變動情況, 計算氮同位素的周轉率及不同蛋白源對大菱鲆生長的貢獻比例。
1 材料與方法
1.1 實驗設計
以魚粉、玉米蛋白和SPC為蛋白源, 魚油和大豆卵磷脂為脂肪源, 按不同比例混合后配制成4種配合飼料。4種飼料在等氮等能的基礎上, 以混合植物蛋白(玉米蛋白和SPC等氮1∶1混合)分別替代飼料中0、10%、30%和50%的魚粉蛋白, 并相應命名為CS0、CS10、CS30和CS50(表 1)。將粉碎后的飼料原料過80目篩, 稱重并采用逐級擴大法混合均勻, 然后加入魚油和適量的蒸餾水二次混合均勻,將油脂顆粒搓散過網, 經螺旋擠壓機加工成直徑4 mm的硬顆粒飼料, 60℃恒溫烘干后冷藏備用。
1.2 飼養管理
實驗用大菱鲆幼魚購于山東省蓬萊宗哲養殖有限公司, 為同批健康魚苗。馴養2周后, 選取體質健壯、均重為(57.72±0.25) g的大菱鲆, 隨機分為4組, 每組設3個平行, 每個平行36尾魚, 分別放于圓柱形養殖桶內(直徑80 cm, 高70 cm), 養殖周期56d。實驗在微流水環境中進行, 24h充氣增氧, 溶氧>6.5 mg/L, 水溫(17.5±0.5)℃, 鹽度在28‰—32‰,pH 7.8—8.0, 亞硝酸氮、氨氮含量均<0.1 mg/L。實驗期間每天飽食投喂2次(8:00, 17:00), 日投喂量為魚體重的1.5%—2%, 根據攝食情況適當調整投喂量。投喂30min后, 從排水口將殘餌排出, 記錄殘餌量。
1.3 樣品采集與生長指標計算
實驗開始前, 隨機取10尾大菱鲆的背肌和肝臟作為初始樣。實驗開始后的7d、14d、28d、42d和56d, 分別從每桶中隨機取3尾魚, 稱重并取其背肌和肝臟, -80℃保存, 用于氮穩定同位素測定。
實驗結束前1天停止投喂, 每桶魚測定總重并記錄總數。預先測定飼料的溶失率, 根據投餌量和矯正的殘餌量計算生長性能指標, 公式如下:
攝食率(Feed intake, FR, %BW/d)=100×攝食量/[(初始體重+末重)/2]/天數;
增重率(weight gain rate, WGR, %)=(末重-初始重)/初始重×100;
肥滿度(Condition factor, CF, g/cm3)=體重/體長3×100;
飼料系數(Feed coefficient, FC)=攝食飼料量/(末重-初始重)×100;
蛋白質效率(PER)=(末重-初始重)/攝入蛋白量(干物質)。
1.4 天然穩定同位素測定
飼料原料及大菱鲆體組織樣品的氮穩定同位素值均采用元素分析儀偶聯同位素比率質譜儀(EA-IRMS, ThermoFinnigan MAT Delta-plus)測定。穩定同位素結果以國際符號δ表示, 如下:
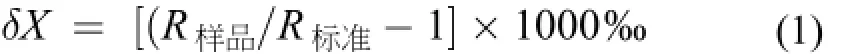
其中, X=15N, R=15N∶14N的比值。氮穩定同位素測定使用的標準品為國際統一標準物, 空氣(Air) N2。
1.5 基于時間的氮同位素周轉模型
本研究采用的時間周轉模型基于Hesslein指數模型[13], 該模型將進食飼料前后機體穩定同位素隨時間的變化歸因于機體組織的生長作用和代謝作用, 如下:
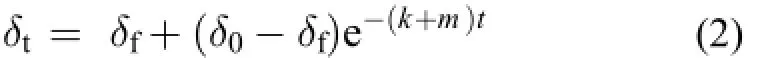
δt為實驗第t天時體組織的穩定同位素值, δf為體組織穩定同位素達到平衡狀態時的δ15N值, δ0為實驗開始時體組織δ15N值。k為實驗期間的特定生長率,m為新陳代謝率, t為實驗持續時間。特定生長率(k)通過如下公式計算[14]:

式中,W0為初始體重, Wf為取樣時間點的體重。由生長作用單獨驅動體組織氮穩定同位素周轉時, 用方程(2)計算, 將m值設為0[15]。機體代謝作用(m)或生長作用(k)對體組織氮穩定同位素周轉的貢獻率(Pm)或(Pg), 用如下方程計算[14]:
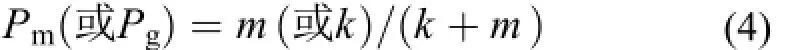
當體組織穩定同位素與實驗飼料達到平衡狀態時,其分餾系數Δ計算如下[16]:

δd是實驗飼料的δ15N值。體組織氮穩定同位素周轉α%所需的時間(T)用如下公式計算[15]:組織中氮穩定同位素周轉50%時, 對應的時間T為周轉半衰期。
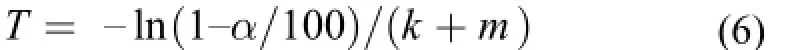
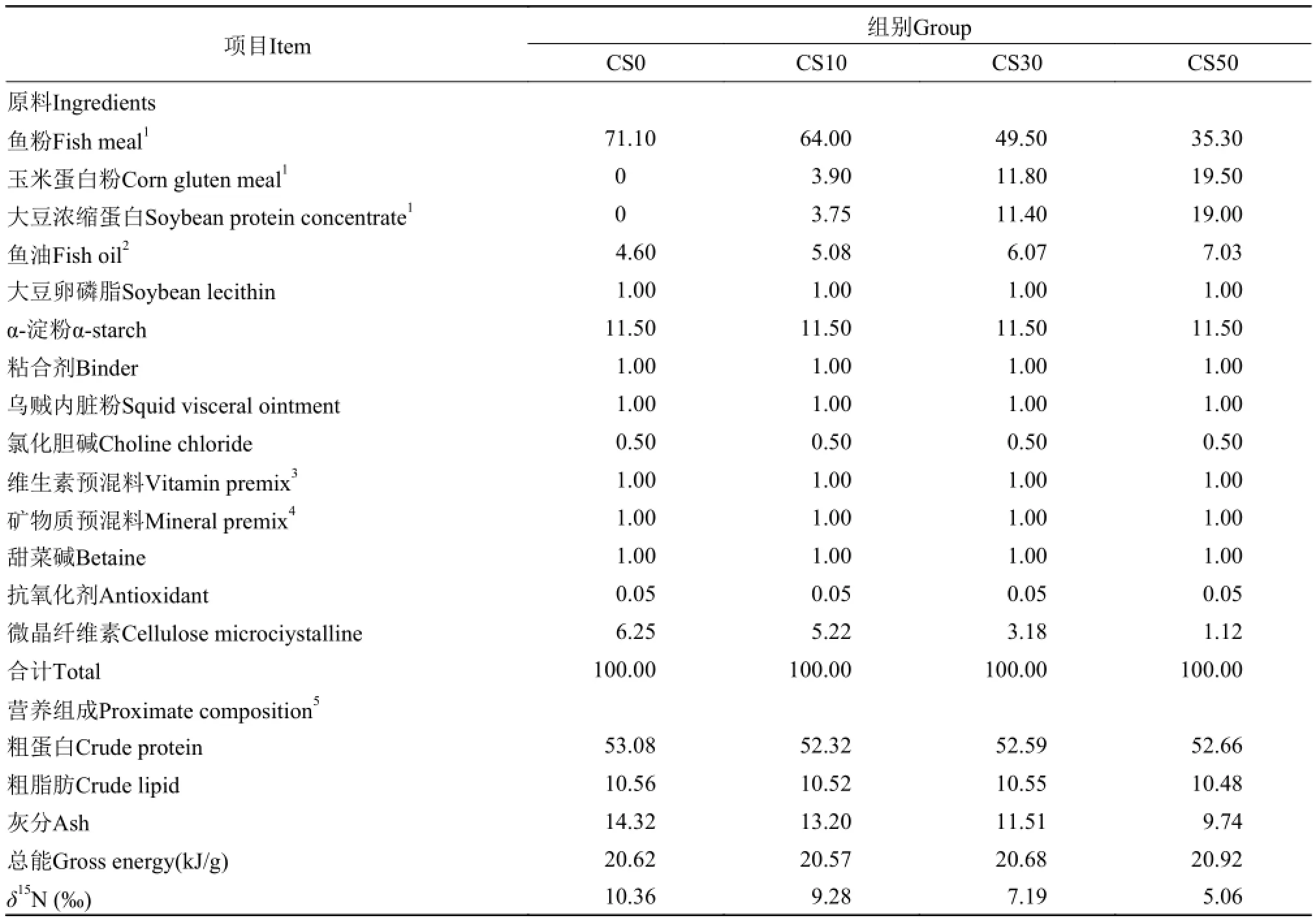
表 1 實驗飼料配方和營養成分(%, 干重)Tab. 1 Formulation and proximate composition of the experimental diets (%, dry weight)
1.6 數據統計分析
運用穩定同位素混合模型[13]和Isosource[4]計算3種蛋白源對大菱鲆幼魚生長的貢獻比例。用軟件SPSS 19.0中ANOVA和Tukey's對數據進行多重比較分析, P<0.05為差異顯著。統計數據用平均值±標準差(Mean±SD)的形式表示。
2 結果
2.1 生長和飼料利用情況
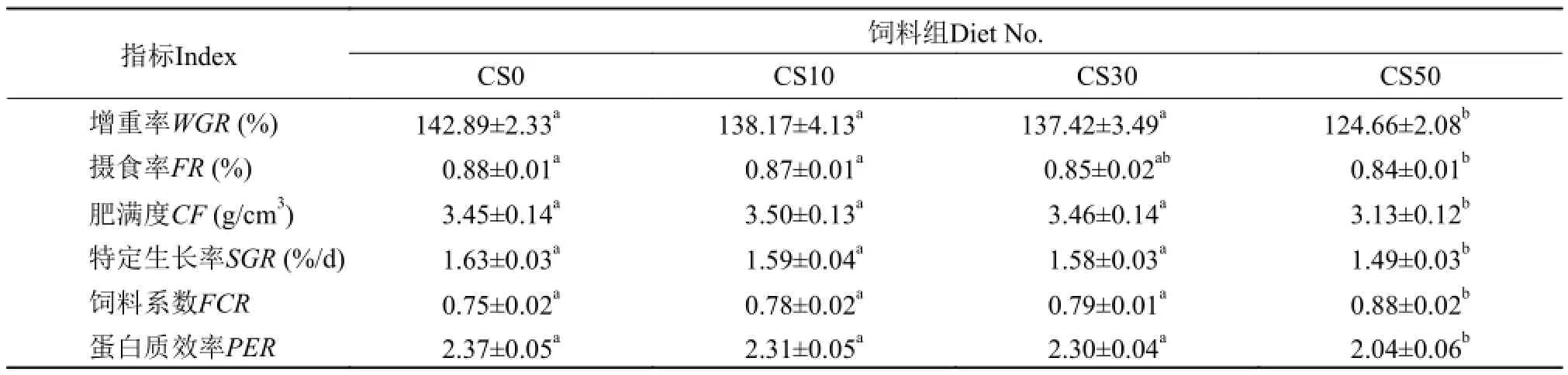
表 2 大菱鲆的生長和飼料利用情況Tab. 2 The effect of four experiment diets on growth performance of turbot fed and feed utilization
由表 2知, 經56d的投喂實驗后CS50組的增重率、肥滿度、特定生長率和蛋白質效率均顯著低于其他組(P<0.05), 而飼料系數則顯著高于其他組(P<0.05)。
2.2 體組織δ15N值隨時間變動情況
各實驗組間肌肉和肝臟初始δ15N值均無差異(P>0.05, 圖 1和圖 2)。隨養殖實驗進行, 肌肉和肝臟的δ15N值均逐漸升高。與初始值相比, CS50組肌肉的δ15N值從14d起出現顯著性差異(P<0.05), 而其他實驗組均從7d起就觀測到顯著性差異(P<0.05); 肝臟均在7d觀測到顯著性差異, 且各實驗組不同取樣點間也均存在顯著性差異(P<0.05)。
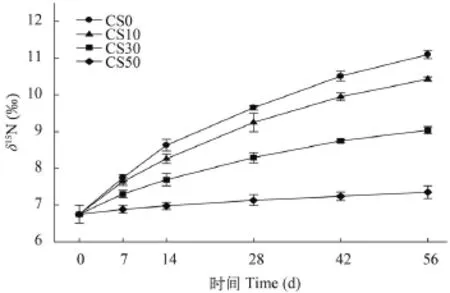
圖 1 投喂不同實驗飼料后大菱鲆肌肉δ15N值隨時間的變動情況Fig. 1 Effects of 4 diets on δ15N values of turbot muscle
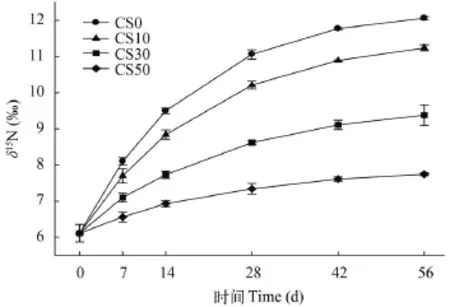
圖 2 投喂不同實驗飼料后大菱鲆肝臟δ15N指隨時間的變動情況Fig. 2 Effects of 4 diets on δ15N values of turbot liver
2.3 生長作用和代謝作用對大菱鲆體組織中氮同位素周轉的驅動
各實驗組由生長作用和代謝作用共同驅動大菱鲆體組織氮同位素的周轉速度(實線)明顯快于由生長作用單獨驅動的氮同位素周轉(虛線)(圖 3)。此外, 肝臟的周轉速度明顯快于肌肉。由表 3知,CS50組肌肉中的m值顯著低于其他組(P<0.05); 各實驗組肝臟中m值隨替代魚粉比例增加而逐漸減小, 但均顯著高于肌肉的m值(P<0.05)。肝臟中代謝作用(m)對氮同位素周轉的貢獻比例(Pm)為63.90%—70.83%, 顯著高于肌肉的26.67%—42.67% (P<0.05)。
2.4 氮同位素的周轉半衰期和分餾系數
基于時間的周轉模型計算可知, CS0、CS10和CS30組間大菱鲆肌肉的T0.5均無顯著性差異(P>0.05), 而CS50組則顯著高于其他組(P<0.05)。各實驗組肝臟的T0.5均存在顯著性差異(P<0.05), 且同組肝臟的T0.5均顯著低于肌肉(P<0.05)(表 3)。圖 4表明, T0.5值和飼料中魚粉替代比例存在明顯的相關性(肌肉的相關方程: y= 2.62x+23.15, r2=0.84; 肝臟的相關方程: y=1.43x+ 12.59, r2=0.96)。肌肉和肝臟中氮同位素周轉達到95%分別需要103.54—140.01d和53.53—72.74d。
由表 3知, δf值隨魚粉替代比例的升高而逐漸降低。CS30和CS50組肌肉的Δ值與CS0和CS10組均有顯著性差異(P<0.05)。各實驗組間肝臟的Δ值也存在顯著性差異(P<0.05)。由圖 5知, Δ與δ15N值呈負相關(y=3.43-0.14x, r2=0.80)。
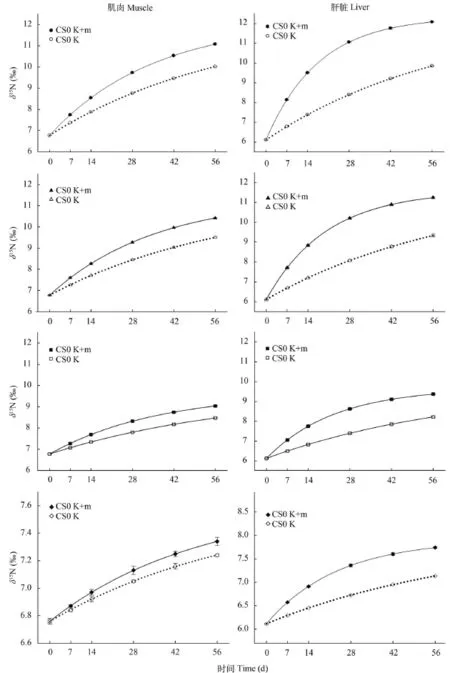
圖 3 基于時間模型估計的大菱鲆肌肉和肝臟中δ15N值的變動情況Fig. 3 The curve of δ15N values of liver and muscle of turbot fed the four experimental diets for 56 days
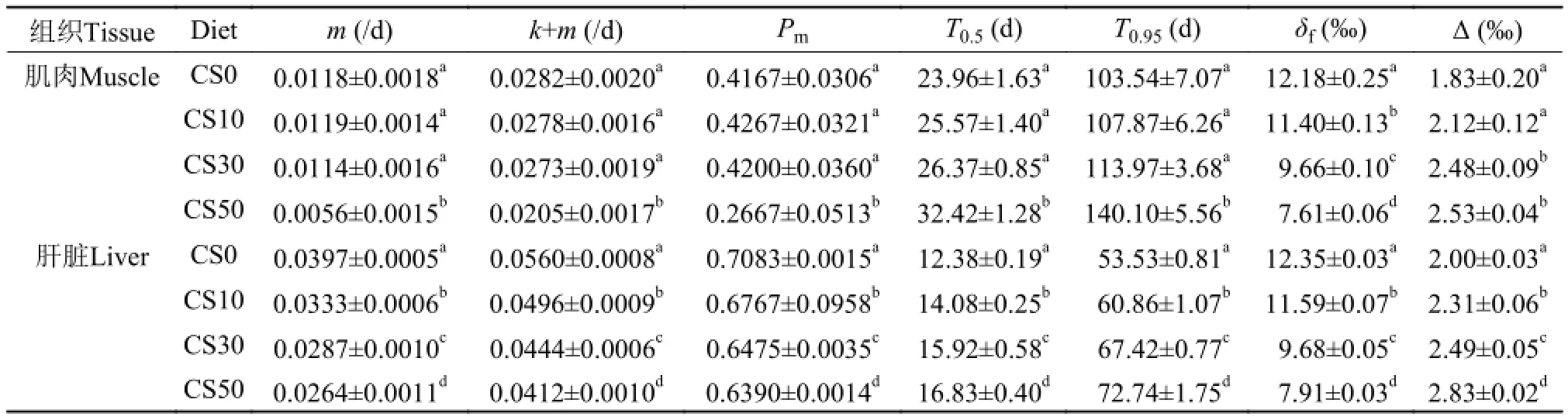
表 3 大菱鲆肝臟和肌肉的周轉半衰期(T0.5)、分餾系數(Δ)、生長作用(k)、代謝作用(m)、代謝作用貢獻(Pm)和最終漸進穩定同位素值(δf)Tab. 3 Time-based half-life (T0.5), fractionation between diet and tissue (Δ), metabolic turnover constant (m), growth constant (k),proportion of metabolic contribution to nitrogen turnover (Pm), final asymptotic nitrogen isotopic value (δf) in the liver and muscle of turbot
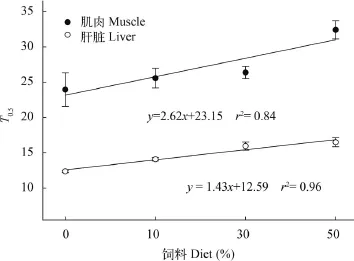
圖 4 大菱鲆肌肉和肝臟δ15N值的周轉半衰期與實驗飼料的魚粉、SPC和玉米蛋白中N替代比例的關系Fig. 4 The relationship of the half-life (T0.5) of δ15N turnover for liver and muscle of turbot to the proportions of substituting N from fish meal with soy protein concentrate and corn meal

圖 5 大菱鲆肌肉和肝臟的Δ值與飼料中δ15N值的關系Fig. 5 The relationship of Δ for liver and muscle of turbot to the δ15N of the experimental diets
2.5 不同蛋白源對大菱鲆生長的貢獻比例
采用Isosource[4]計算CS10、CS30和CS50組中魚粉、玉米蛋白和SPC對大菱鲆生長的貢獻比例分別為80%、7%和13%, 62%、11%和27%, 46%、13%和41% (圖 6)。玉米蛋白和SPC對大菱鲆的貢獻比例隨飼料中替代魚粉比例的增加而顯著升高(P<0.05)。玉米蛋白對大菱鲆的貢獻比例顯著低于SPC和魚粉(P<0.05)。
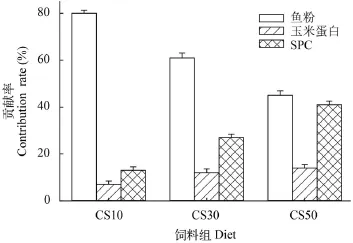
圖 6 各實驗組中三種蛋白源對大菱鲆生長的貢獻比例Fig. 6 Effects of three dietary protein sources to the growth of turbot
3 討論
3.1 對大菱鲆生長性能和飼料利用的影響
Kotaro[17]研究表明, 玉米蛋白可替代日本比目魚(Paralichthys olivaceus)飼料中40%的魚粉而不影響其生長。劉興旺等[18, 19]研究發現, 當飼料中玉米蛋白含量超過25%時, 大菱鲆特定生長率和飼料利用率將顯著降低, 而SPC替代80%魚粉時大菱鲆特定生長率才顯著低于其他組。本研究用玉米蛋白和SPC同時替代大菱鲆飼料中的魚粉, 結果表明隨替代魚粉比例的增加, 大菱鲆的增重率、特定生長率、攝食率以及蛋白質效率均呈下降趨勢, 而飼料系數有升高趨勢, 當替代比例達到50%時, 大菱鲆的生長和飼料利用率均顯著低于其他組。這主要是由于隨植物蛋白源替代魚粉比例的升高, 飼料的適口性越來越差, 導致攝食率逐漸下降[20]。此外,植物中的抗營養因子和氨基酸不平衡也會降低大菱鲆的表觀消化率[21—23]。
3.2 氮穩定同位素在體組織中的周轉情況及周轉半衰期
本研究表明, 不同實驗組間大菱鲆肌肉和肝臟的氮同位素周轉速度均存在差異。當替代50%魚粉時, 大菱鲆肌肉的周轉半衰期顯著高于其他組,且肝臟的周轉半衰期隨替代魚粉比例的升高而延長, 這可能是由于不同實驗組的攝食蛋白來源和品質存在差異[24]。氮穩定同位素周轉半衰期與飼料中替代魚粉比例的線性相關關系(肌肉: r2=0.84; 肝:r2=0.96)也說明了攝食蛋白來源和品質對大菱鲆組織中氮穩定同位素周轉代謝的影響。
當生物個體攝食與之前食物具有不同穩定同位素信號的新食物時, 其體組織中穩定同位素的比率取決于同位素在其體組織中的周轉速度[25, 26]。本研究結果顯示, 大菱鲆肝臟的周轉半衰期(12.38—16.83d)顯著低于肌肉(23.96—32.42d), 這表明氮穩定同位素在大菱鲆肝臟中的周轉速度明顯快于肌肉的。Suzuki等[27]研究發現, 日本鱸肌肉中氮同位素的周轉半衰期為25.7d, 而肝臟氮同位素周轉半衰期為14.4d。Logan等[28]在底鳉上的研究也表明肝臟中氮同位素的周轉速度顯著快于肌肉。這主要由于肝臟的組織替換及新陳代謝速度要比肌肉的快[29]。MacAvoy等[30]研究指出, 由于不同體組織代謝速度有所不同, 導致機體不同組織間同位素的周轉存在差異, 代謝越旺盛的組織其同位素的周轉率相應越快, 周轉半衰期也越短。
3.3 生長作用和代謝作用對大菱鲆體組織中氮同位素周轉的驅動
本研究發現大菱鲆體組織氮同位素的周轉速度要顯著快于由生長作用單獨驅動的氮同位素周轉, 這說明基于組織增加的生長作用和基于組織替代的代謝作用共同驅動了實驗期間大菱鲆肌肉和肝臟的氮同位素周轉[23]。肝臟中代謝作用對氮同位素周轉的貢獻率(63.90%—70.83%)顯著高于肌肉(26.67%—42.67%), 這說明組織替代在肝臟的氮同位素周轉中起主要作用, 而生長在肌肉氮同位素周轉中起主導作用。在白鮭(Coregonus lavaratus)[25]、底鳉幼魚(Fundulus heteroclitus)[28]和筍殼魚(Pomatoschistus minutus)[31]的研究中也得到了類似的研究結果。
3.4 分餾系數
大菱鲆氮同位素的分餾系數范圍為1.83‰—2.83‰, 且各實驗組中肌肉和肝臟的分餾系數均隨飼料中替代魚粉比例的增加而顯著升高。Le Vay等[32]研究表明分餾系數的大小與攝食蛋白的水平、質量和氨基酸水平等緊密相關。本研究中, 分餾系數與飼料中δ15N值呈明顯的負相關性(r2=0.80),進一步證明了這一假設。此外, 各實驗組間肌肉和肝臟的Δ值均存在差異。Xia等[14]研究指出氮同位素的分餾系數在不同物種或同一物種不同組織間均有差異。Sweeting等[33]研究發現歐洲鱸的肌肉、心臟和肝臟中氮同位素的分餾系數逐漸降低。不同體組織間分餾系數的差異可能與不同體組織的生化組成差異有關[24]。
3.5 魚粉蛋白、玉米蛋白和SPC對大菱鲆的貢獻比率
在本研究中替代10%魚粉蛋白時, 玉米蛋白和SPC對大菱鲆生長和代謝的貢獻比例(7%和13%)均高于其在飼料中的替代比例。黃云等[34]曾報道在飼料中補充適量植物蛋白時, 可以增強魚腸胃的蠕動, 提高營養物質的利用率。Li等[35]研究也發現微量的抗營養因子能促進星斑川鰈消化酶的分泌, 促進其對飼料的消化利用。替代30%和50%魚粉蛋白時, 玉米蛋白貢獻比例則顯著低于其在飼料中的替代比例, 同時顯著低于SPC的貢獻比例, 而SPC的貢獻比率均顯著高于其在飼料中的替代比例。周暉等[36]研究表明, 用10%玉米蛋白替代軍曹魚飼料中等量魚粉蛋白后, 玉米蛋白對軍曹魚生長的貢獻比率顯著低于飼料中的替代比例, 這主要與玉米蛋白的氨基酸不平衡有關。劉興旺等[18, 19]研究表明, 玉米蛋白可以替代大菱鲆幼魚飼料中25%的魚粉而不影響其生長, 而SPC替代比例大于80%時, 大菱鲆的特定生長率才顯著下降。這均表明大菱鲆對SPC的利用效率顯著優于玉米蛋白。此外, 研究還發現玉米蛋白和SPC對大菱鲆生長貢獻比例的增加量, 隨著飼料中替代魚粉比例的升高而減少, 這說明隨著飼料中玉米蛋白和SPC替代比例的升高,大菱鲆對飼料中植物蛋白的利用效率逐漸下降。
4 結論
本研究表明, 用玉米蛋白和SPC可同時替代大菱鲆飼料中30%的魚粉蛋白而不影響其生長。生長作用和代謝作用共同驅動了大菱鲆體組織中氮穩定同位素的周轉代謝, 肝臟氮穩定同位素的周轉速度明顯快于肌肉的, 相應的周轉半衰期更短。各實驗組中SPC對大菱鲆生長的貢獻比例要大于玉米蛋白, 說明大菱鲆對SPC的利用效率優于玉米蛋白。
[1]Luo J Q, Cao Z M, Chen D W, et al. The challenge of studies on protein nutrition [J]. China Journal of Animal Science, 2012, 48(13): 73—76 [羅鈞秋, 曹中明, 陳代文,等. 蛋白質營養研究新階段. 中國畜牧雜志, 2012,48(13): 73—76]
[2]Zhang Y, Zhou A G. Protein turnover and measurement [J]. Chinese Journal of Animal Nutrition, 2001, 13(4):7—13 [張勇, 周安國. 蛋白質周轉代謝及其測定. 動物營養學報, 2001, 13(4): 7—13]
[3]Gamboa-Delgado J, Le Vay L. Natural stable isotopes as indicators of the relative contribution of soy protein and fish meal to tissue growth in Pacific white shrimp (Litopenaeus vannamei) fed compound diets [J]. Aquaculture, 2009, 291(1—2): 115—123
[4]Gamboa-Delgado J, Rojas-Casas M G, Nieto-López M G,et al. Simultaneous estimation of the nutritional contribution of fish meal, soy protein isolate and corn gluten to the growth of Pacific white shrimp (Litopenaeus vannamei) using dual stable isotope analysis [J]. Aquaculture,2013, 380: 33—40
[5]Abimorad E G, Ducatti C, Castellani D, et al. The use of stable isotopes to investigate the effects of supplemental lysine and methionine on protein turnover and amino acid utilization in pacu, Piaractus mesopotamicus, juveniles [J]. Aquaculture, 2014, 433: 119—124
[6]Pan Y J. Aquatic Dictionary [M]. Shanghai: Shanghai Dictionary Press. 2007, 284—353 [潘迎捷. 水產辭典. 上海: 上海辭書出版社. 2007, 284—353]
[7]Wang J Y, Li P Y, Song Z D, et al. Effect of dietary Yucca Schidigera extract on the growth performance,blood physiological and biochemicals of turbot (Scophthalmus maximus) and water environment [J]. Acta Hydrobiologica Sinica, 2014, 38(6): 1117—1126 [王際英,李培玉, 宋志東, 等. 飼料中添加絲蘭提取物對大菱鲆幼魚生長和生理及水環境的影響. 水生生物學報, 2014,38(6): 1117—1126]
[8]Liu X Q, Zhu X M, Han D, et al. Effects of dietary ratio of fish meal and rapeseed meal on the growth and feed utilization Gibel garp (Carassius auratus gibelio) [J]. Acta Hydrobiologica Sinica, 2014, 38(4): 657—663 [劉曉慶, 朱曉鳴, 韓冬, 等. 飼料魚粉, 菜粕比例對異育銀鯽生長和飼料利用的影響. 水生生物學報, 2014, 38(4):657—663]
[9]Shi X, Luo Z, Huang C, et al. Effect of substituting Chlorella sp. for regular fishmeal on growth, body composition, hepatic lipid metabolism and histology in crucian carp Carassius auratus [J]. Acta Hydrobiologica Sinica, 2015, 39(3): 498—506 [石西, 羅智, 黃超, 等. 小球藻替代魚粉對鯽生長, 體組成, 肝臟脂肪代謝及其組織學的影響. 水生生物學報, 2015, 39(3): 498—506]
[10]Naylor R L, Hardy R W, Bureau D P, et al. Feeding aquaculture in an era of finite resources [J]. Proceedings of the National Academy of Sciences, 2009, 106(36):15103—15110
[11]Tusche K, Arning S, Wuertz S, et al. Wheat gluten and potato protein concentrate Promising protein sources for organic farming of rainbow trout (Oncorhynchus mykiss)[J]. Aquaculture, 2012, 344: 120—125
[12]Yan Q G, Zhu X M, Yang Y X, et al. Effect of replacement of fish meal with cottonseed meal on growth, hematological physiology, and body composition of grass garp [J]. Acta Hydrobiologica Sinica, 2014, 38(2):362—369 [嚴全根, 朱曉鳴, 楊云霞, 等. 飼料中棉粕替代魚粉蛋白對草魚的生長, 血液生理指標和魚體組成的影響. 水生生物學報, 2014, 38(2): 362—369]
[13]Phillips D L, Gregg J W. Source partitioning using stable isotopes: coping with too many sources [J]. Oecologia,2003, 136(2): 261—269
[14]Xia B, Gao Q F, Li H, et al. Turnover and fractionation of nitrogen stable isotope in tissues of grass carp Ctenopharyngodon idellus [J]. Aquaculture Environment Interactions, 2013, 3: 177—186
[15]Hesslein R H, Hallard K A, Ramlal P. Replacement of sulfur, carbon, and nitrogen in tissue of growing broad white fish (Coregonus nasus) in response to a change in diet traced by34S,13C and15N [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1993, 50(10): 2071—2076
[16]Buchheister A, Latour R J. Turnover and fractionation of carbon and nitrogen stable isotopes in tissues of a migratory coastal predator, summer flounder (Paralichthys dentatus) [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2010, 67(3): 445—461
[17]Kikuchi K. Partial replacement of fish meal with corn gluten meal in diets for Japanese flounder Paralichthys olivaceus [J]. Journal of the World Aquaculture Society,1999, 30(3): 357—363
[18]Liu X W, Ai Q H, Mai K S, et al. Effects of replacing fish meal with soy protein concentrate on feed intake and growth of turbot (Scophthalmus maximus) [J]. Journal of Fisheries of China, 2014, 38(1): 91—98 [劉興旺, 艾慶輝, 麥康森, 等. 大豆濃縮蛋白替代魚粉對大菱鲆攝食、生長及體組成的影響. 水產學報, 2014, 38(1):91—98]
[19]Liu X W, Mai K S, Ai Q H, et al. Replacement of fish meal by corn gluten meal in diets of Scophthatmus maximus [J]. Journal of Fisheries of China, 2012, 36(3):466—472 [劉興旺, 麥康森, 艾慶輝, 等. 玉米蛋白粉替代魚粉對大菱鲆攝食、生長及體組成的影響. 水產學報, 2012, 36(3): 466—472]
[20]Twibell R G, Wilson R P. Preliminary evidence that cho-lesterol improves growth and feed intake of soybean meal-based diets in aquaria studies with juvenile channel catfish, Ictalurus punctatus [J]. Aquaculture, 2004,236(1): 539—546
[21]Ai Q H, Xie X J. Advance in utilization of plant proteins by aquatic animals [J]. Periodical of Ocean University of China (Natural Science Edition), 2005, 36(6): 929—935[艾慶輝, 謝小軍. 水生動物對植物蛋白源利用的研究進展. 中國海洋大學學報(自然科學版), 2005, 36(6):929—935]
[22]Hardy R W. Utilization of plant proteins in fish diets: effects of global demand and supplies of fish meal [J]. Aquaculture Research, 2010, 41(5): 770—776
[23]Deng J, Mai K, Ai Q, et al. Alternative protein sources in diets for Japanese flounder Paralichthys olivaceus (Temminck and Schlegel): Ⅱ. Effects on nutrient digestibility and digestive enzyme activity [J]. Aquaculture Research,2010, 41(6): 86—870
[24]Xia B, Wang J, Gao Q F, et al. The nutritional contributions of dietary protein sources to tissue growth and metabolism of sea cucumber Apostichopus japonicus (Selenka): Evidence from nitrogen stable isotope analysis [J]. Aquaculture, 2015, 435: 237—244
[25]Guelinckx J, Maes J, Van Den Driessche P, et al. Changes in δ13C and δ15N in different tissues of juvenile sand goby Pomatoschistus minutus: a laboratory dietswitch experiment [J]. Maine Ecology Progress Series,2007, 341: 205—215
[26]Harvey C J, Hanson P C, Essinton T E, et al. Using bioenergetic models to predict stable isotope ratios in fishes [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2002, 59(1): 115—124
[27]Suzuki K W, Kasai A, Nakayama K, et al. Diferential isotopic enrichment and half-life among tissues in Japanese temperate bass (Lateolabrax japonicus) juveniles: implications for analyzing migration [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2005, 62(3): 671—678
[28]Logan J, Haas H, Deegan L, et al. Turnover rates of nitrogen stable isotopes in the salt marsh mummichog, Fundulus heteroclitus, following a laboratory diet switch [J]. Oecologia, 2006, 147(3): 391—395
[29]Buchheister A, Latour R J. Turnover and fractionation of carbon and nitrogen stable isotopes in tissues of a migratory coastal predator, summer flounder (Paralichthys dentatus) [J]. Canadian Journal of Fisheries and Aquatic Sciences, 2010, 67(3): 445—461
[30]MacAvoy S E, Ameson L S, Bassett E. Correlation of metabolism with tissue carbon and nitrogen turnover rate in small mammals [J]. Oecologia, 2006, 150(2): 190—201
[31]Perga M E, Gerdeaux D. ‘Are fish what they eat'all year round [J]? Oecologia, 2005, 144(4): 598—606
[32]Le Vay L, Gamboa-Delgado J. Naturally-occurring stable isotopes as direct meausres of larval feeding efficiency,nutrient incorporation and turnover [J]. Aquaculture,2011, 315(1): 95—103
[33]Sweeting C J, Barry J, Barnes C, et al. Effects of body size and environment on diet-tissue fractionation in fishes [J]. Journal of Experimental Marine Biology and Ecology, 2007, 340(1): 1—10
[34]Huang Y, Xiao D Y, Hu Y. Research status of alternative dietary protein sources for fish feed [J]. Feed Review,2010, (2): 11—15 [黃云, 肖調義, 胡毅. 魚類飼料中替代蛋白營養的研究進展. 飼料博覽, 2010, (2): 11—15]
[35]Li P Y, Wang J Y, Song Z D, et al. Evaluation of soy protein concentrate as a substitute for fish meal in diets for juvenile starry flounder (Platichthys stellatus) [J]. Aquaculture, 2015, 448: 578—585
[36]Zhou H, Chen G, Gu B H, et al. Contribution rate of dietary corn protein to protein growth of juvenile cobia (Rachycentron canadum L.) [J]. Chinese Journal of Animal Nutrition, 2013, 25(11): 2633—2642 [周暉, 陳剛, 古濱河, 等. 飼料中的玉米蛋白質對軍曹魚幼魚蛋白質生長的貢獻率. 動物營養學報, 2013, 25(11): 2633—2642]
THE NUTRIONAL CONTRIBUTIONS OF DIETARY PROTEIN SOURCES TO GROWTH OF TURBOT (SCOPHTHATMUS MAXIMUS L.): EVIDENCE FROM STABLE ISOTOPE ANALYSIS (SIA)
LI Zhong-Qing1, 2, XIA Bin2, ZHANG Li-Min2, WANG Ji-Ying2, LI Bao-Shan2, SUN Yong-Zhi2, 3and WANG Ming-Hui1, 2
(1. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China; 2. Shandong Marine Resource and Environment Research Institute, Yantai 264006, China; 3. Shengsuo Fishery Feed Research Centre of Shandong Province, Yantai 265500, China)
To investigate effects of dietary protein sources to turbot growth (Scophthatmus maximus L.), we substituted fish meal by plant protein blend (corn gluten and soy protein concentrate 1∶1 isonitrogenous mixed) in diets and determined nitrogen stable isotope ratios of diet and tissue. Four isonitrogenous and isoenergetic diets with different replacement 0, 10%, 30% and 50% fish meal protein by plant protein blend were formulated and correspondingly named as CS0, CS10, CS30 and CS50, respectively, for a 56 days experiment with 3 replicates in each group. Our results showed that food intake (FI) and specific growth rate (SGR) in CS50 were significantly lower but the feed coefficiency was higher compared with other groups. The turnover rates of nitrogen stable isotope were significantly different between each diet (P<0.05) with the highest turnover rate in the control group. Liver responded more quickly to the diet compared with muscle. Nitrogen half-lives (T0.5) of muscle were significantly higher than those of liver (P<0.05). The proportional contributions of growth to nitrogen turnover were 57.33%—73.33% for muscle and 29.17%—36.10% for liver, respectively. Based on turnover model, the fractionation (Δ) were ranged from 1.83‰ to 2.83‰. Calculated by Isosource software, the contributions of fish meal, corn gluten and SPC to tissue growth and metabolism of turbot were 80%, 7% and 13% for CS10, 62%, 11% and 27% for CS30, 46%, 13% and 41% for CS50, respectively. The result showed that stable isotopic turnover was driven by both tissue replacement in the form of metabolism and tissue increase in the form of growth, the proportional contritetiens of which between tissues exhibited significant discrepancy;nitrogen stable isotope, turnover rate of liver was faster than that of muscle, without, the half-time of liver was also faster; the contribution rate of dietary corn protein to the protein growth of the turbot was lower than the SPC.
Scophthatmus maximus L.; Nitrogen stable isotope; Growth; Metabolism; Turnover model
S917.4
A
1000-3207(2016)04-0843-10
10.7541/2016.109
2015-09-07;
2016-01-11
水生動物營養與飼料創新示范平臺項目(201501006); 國家海洋公益性行業科研專項(201205025); 山東省科技發展計劃(2014GHY115006)資助 [Supported by the Aquatic Animal Nutrition and Feed Research and Innovation Demonstration Platform (201501006); Public Science and Technology Research Funds of Ocean from the State Oceanic Administration, People's Republic of China (201205025); Shandong Science and Technology Plan Project (2014GHY115006)]
李忠清(1990—), 男; 山東臨沂人; 碩士; 研究方向為水產動物營養與飼料。 E-mail: longxingzhongqing@163.com
張利民, 研究員; 碩士生導師; E-mail: ytzlm@139.com; 王際英, 研究員; 碩士生導師; E-mail: ytwjy@126.com
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
