表達H1N1亞型豬流感病毒三聚體HA重組慢病毒的免疫原性研究①
2016-08-09 07:37:58鄧穎琦李俊峰唐雨博孫藝學王偉利叢彥龍
中國免疫學雜志 2016年7期
鄧穎琦 李俊峰 曲 輝 唐雨博 孫藝學 李 鑫 王偉利 丁 壯 叢彥龍
(吉林大學動物醫學學院,長春130062)
?
表達H1N1亞型豬流感病毒三聚體HA重組慢病毒的免疫原性研究①
鄧穎琦李俊峰②③曲輝②④唐雨博孫藝學⑤李鑫王偉利⑥丁壯叢彥龍
(吉林大學動物醫學學院,長春130062)
[摘要]目的:探討表達豬H1N1亞型流感病毒三聚體HA的重組慢病毒(rLV-HA-GCN4)對BALB/c小鼠的免疫保護效果。方法:將雌性BALB/c小鼠隨機分為重組慢病毒rLV-HA-GCN4組、重組慢病毒rLV-HA組、慢病毒LV空載體對照組及PBS對照組,先后進行質粒和慢病毒2次免疫,間隔為2周,注射部位均為小鼠大腿內側肌肉;于初次免疫后的第28天,各組小鼠鼻腔滴注50 μl 100TCID50的H1N1病毒,采用淋巴細胞轉化實驗、流式細胞術、間接ELISA和脾肺指數檢測各免疫指標。結果:加強免疫后第14天,rLV-HA-GCN4組脾淋巴細胞轉化率為0.3±0.11,與PBS組相比,差異有統計學意義(P<0.01),產生以Th1型CD4+ T細胞為主的細胞免疫反應;rLV-HA-GCN4組小鼠IgG抗體效價可達1∶8 000,攻毒后第14天約為1∶7 000;攻毒后,rLV-HA-GCN4組脾肺指數小于PBS組,小鼠體重在前3 d略有下降,隨后逐漸上升,兩個指標與PBS組相比,差異均有統計學意義(P<0.05)。結論:rLV-HA-GCN4能夠誘導小鼠良好的細胞與體液免疫應答,從而有效地保護小鼠抵御豬H1N1流感病毒的感染。
[關鍵詞]豬流感;H1N1亞型;三聚體HA;慢病毒載體;免疫原性
流行性感冒(Influenza)簡稱流感(flu),是由流感病毒(Influenza virus)引起的一種潛伏期短、傳染性強、傳播迅速的急性呼吸道傳染病。根據核蛋白(NP)和基質蛋白(M)抗原性的不同,將流感病毒分為3個型,即甲(A)、乙(B)、丙(C)型,其中A型流感病毒宿主感染譜最為廣泛,包括多種禽類和哺乳動物。A型流感病毒根據血凝素(HA)和神經氨酸酶(NA)抗原性的不同可分為多種不同的亞型。其中H1N1亞型流感病毒自1918年發現至今,不僅導致了養豬業嚴重的經濟損失,而且作為引發人群季節性流感的主要致病亞型,也給人類健康帶來了巨大威脅。此外,H1N1亞型病毒在豬群與人群中的跨種傳播也受到了密切關注,因此防控該亞型流感具有重要的公共衛生學意義[1,2]。本研究對前期利用慢病毒載體系統包裝的表達H1N1亞型豬流感病毒三聚體HA的重組慢病毒進行了免疫原性評價,為今后防控H1N1亞型豬流感病毒提供理論和實驗依據。
1材料與方法
1.1材料
1.1.1病毒H1N1亞型豬流感病毒由吉林大學動物醫學學院傳染病實驗室分離并保存;重組慢病毒rLV-HA-GCN4、重組慢病毒rLV-HA、慢病毒LV均由吉林大學動物醫學學院傳染病實驗室構建包裝[3]。
1.1.2實驗動物60只6~8周齡SPF雌性BALB/c小鼠,體重15~18 g,購自北京華阜康生物科技股份有限公司,合格證號:SCXK(京)2012-0004。
1.1.3主要試劑紅細胞裂解液、MTT購自碧云天生物有限公司;ConA、PMA、離子酶素購自Sigma公司;阻斷劑BFA、穿膜劑購自eBioscience公司;APC anti-mouse CD3、FITC anti-mouse CD4、PE anti-mouse CD8、FITC anti-mouse TNF-α、PE anti-mouse IFN-γ、PE anti-mouse IL-12、PE anti-mouse IL-4、FITC anti-mouse IL-10均購自BD Biosciences公司;1640細胞培養液購自Gibco公司;兔抗鼠酶標二抗購自Southern Biotech公司。
1.2方法
1.2.1動物分組、免疫及攻毒將飼養于溫度22℃、相對濕度50%~60%環境中的小鼠隨機分為4組:重組慢病毒rLV-HA-GCN4組、重組慢病毒rLV-HA組、慢病毒LV空載體對照組及PBS對照組。初次免疫時,免疫組小鼠分別肌注各慢病毒質粒15 μg/100 μl,PBS組肌注PBS 100 μl;于初次免疫后的第14天,免疫組再分別以各106轉導單位(TU)的慢病毒進行加強免疫。質粒與慢病毒免疫部位均為小鼠大腿內側肌肉;于初次免疫后的第28天,小鼠鼻腔滴注50 μl 100TCID50的H1N1病毒,每日觀察小鼠狀態并稱重。
1.2.2脾細胞的制備加強免疫后第14天每組取3只小鼠,脫頸處死后無菌剪取部分脾臟置于200目細胞篩網上,充分研磨后將細胞懸液轉入5 ml離心管中,2 000 r/min×5 min,棄上清,加入紅細胞裂解液,混勻后立即加入1 ml含有10%血清的1640培養液中,2 000 r/min×5 min;用1640培養液洗滌2次后,再以1 ml培養液重懸細胞并計數。
1.2.3淋巴細胞轉化率的測定采用MTT法。將1×106ml-1的脾細胞懸液鋪至96孔細胞培養板中,每孔200 μl,置37℃、5%CO2培養箱中培養。待細胞長至單層后,用終濃度為5 μg/ml ConA刺激,未加刺激劑的自然增殖孔以細胞培養液代替,培養72 h后,加入10 μl MTT(5 mg/ml),繼續培養4 h后,離心棄上清,加入150 μl DMSO,室溫避光孵育20 min后,在酶標儀490nm波長處測定吸光度值(A)。
1.2.4T細胞亞群的檢測采用流式細胞術。將1×106ml-1的脾細胞懸液鋪至96孔細胞培養板中,每孔200 μl,置37℃、5%CO2培養箱中培養。待細胞長至單層后,以100 μl FACS buffer重懸細胞,每100萬個細胞分別加入1 μg的APC anti-mouse CD3、FITC anti-mouse CD4、PE anti-mouse CD8,同時設置同型對照。4℃避光孵育1 h后,2 000 r/min×5 min,用1640培養液洗滌2次,以100 μl FACS buffer重懸細胞,經尼龍膜濾至流式細胞管,上流式細胞儀進行檢測。
1.2.5脾細胞胞內細胞因子的檢測采用流式細胞術。將1×106ml-1的脾細胞懸液鋪至96孔細胞培養板中,每孔200 μl,置37℃、5%CO2培養箱中培養。待細胞長至單層后,每孔加入PMA與離子酶素(終濃度為10 mg/ml),12 h后加入BFA(3 μg/ml),培養5 h后收集4個復孔細胞。2 000 r/min×5 min棄上清,用PBS重懸細胞;2 000 r/min×5 min,棄上清,每管加入100 μl 4%多聚甲醛,室溫避光孵育20 min;每管加入1 ml穿膜劑,混勻,2 000 r/min×5 min,棄上清。重復上述操作步驟1次。每管加入400 μl PBS,混勻后分裝至100 μl/管。分別加入各10 μl的FITC anti-mouse TNF-α、PE anti-mouse IFN-γ、PE anti-mouse IL-12、PE anti-mouse IL-4、FITC anti-mouse IL-10,4℃孵育1 h后,用FACS buffer洗滌2次,2 000 r/min×5 min,剩余200 μl上清,經尼龍膜濾至流式細胞管,上流式細胞儀進行檢檢測。
1.2.6抗體效價的檢測采用間接ELISA方法。分別于初次免疫、加強免疫和攻毒后的第14天,經小鼠眼眶無菌采血。37℃水浴30 min,4℃ 1 500 r/min×10 min,分離血清。采用間接ELISA方法測定血清中IgG抗體效價:將100 μl病毒液加入96孔ELISA板,4℃包被過夜;每孔加入150 μl BSA,37℃封閉2 h;每孔加入100 μl不同稀釋度的待檢血清,37℃作用1 h;每孔加入100 μl HRP標記的兔抗鼠酶標二抗(1∶5 000稀釋),37℃作用1 h。上述各步驟之間均用PBST洗滌3次,每次5 min。二抗孵育后每孔加入50 μl底物,避光顯色;當陽性對照孔顯色時,每孔加入50 μl終止液,在酶標儀450nm波長處測定A值,A≥0.2判為陽性。
1.2.7脾肺指數的測定于攻毒后第14天,將小鼠斷頸,無菌取出小鼠脾臟和肺臟,用濾紙吸干表面水分后稱重,并計算:脾指數=(脾重/體重)×100%;肺指數=(肺重/體重)×100%。
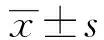
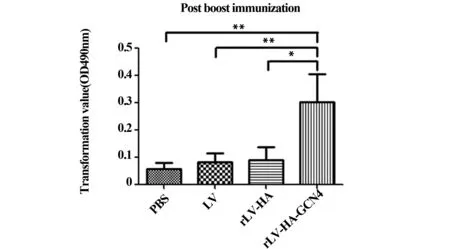
圖1 淋巴細胞轉化結果Fig.1 Result of lymphocyte transformationNote: *.P<0.05;**.P<0.01.
2結果
2.1細胞免疫水平
2.1.1淋巴細胞轉化水平加強免疫后第14天,rLV-HA-GCN4組淋巴細胞轉化率(TV)遠大于其他組,TV=0.3±0.11,與其他3組比較,差異有統計學意義(P<0.05)。見圖1。
2.1.2胞內細胞因子水平與對照組比較,初次免疫后第14天,rLV-HA-GCN4組各細胞因子含量均增加,其中IL-10、TNF-α分泌較多,分別達到26.13±2.51、18.67±1.17;加強免疫后第14天,各組細胞因子水平均有所下降,但重組慢病毒免疫組各細胞因子表達量均高于PBS組,以IFN-γ、TNF-α分泌較多。見圖2。
2.1.3CD4+/CD8+T細胞亞群檢測結果顯示,初次免疫后第14天,與LV組和PBS組相比,rLV-HA-GCN4組與rLV-HA組兩個亞群T細胞的百分含量均升高,且各組CD4+T細胞含量高于CD8+T細胞;與初次免疫相比,加強免疫后第14天,兩個亞群T細胞的百分含量均有所增加,且以rLV-HA-GCN4組CD4+T細胞含量增加最為明顯,為38.27±3.32,與PBS組相比,差異有統計學意義(P<0.01)。攻毒后第14天, rLV- HA- GCN4組
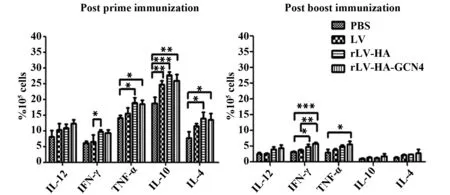
圖2 胞內細胞因子水平Fig.2 Expression level of intracellular cytokinesNote: *.P<0.05;**.P<0.01;***.P<0.001.

圖3 CD4+/CD8+ T細胞亞群分析Fig.3 Analysis of CD4+/CD8+ T cellsNote: *.P<0.05;**.P<0.01.
圖4 IgG抗體滴度Fig.4 IgG titers
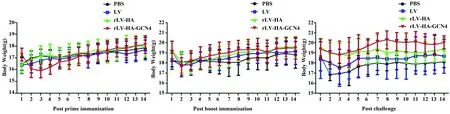
圖5 免疫攻毒后小鼠體重變化Fig.5 Body weight change of mice after immunization and challenge
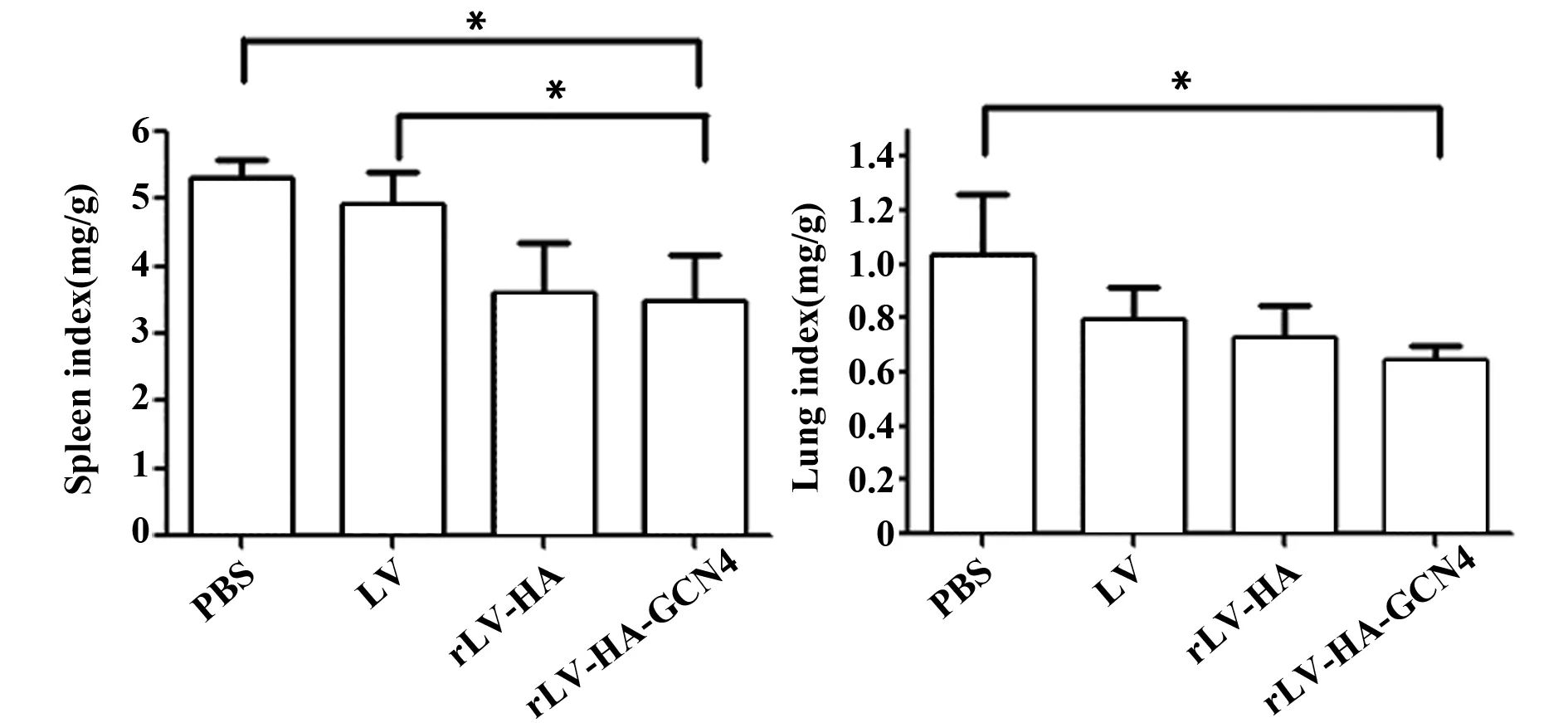
圖6 攻毒后小鼠的脾肺指數Fig.6 Spleen and lung indexes in mice after challengeNote: *.P<0.05.
CD4+T細胞的百分含量下降,為33.93±4.8,而CD8+T細胞的百分含量增加至20.9±0.98,與PBS組相比,差異有統計學意義(P<0.05)。見圖3。
2.2體液免疫水平ELISA檢測結果顯示,初次免疫后第14天,rLV-HA-GCN4組與rLV-HA組HA抗體效價約為1∶1 000;加強免疫后第14天,HA抗體效價明顯升高,rLV-HA-GCN4組A=0.208時的稀釋倍數為1∶8 000,rLV-HA組A=0.203時的稀釋倍數為1∶2 000;攻毒后第14天,rLV-HA-GCN4組HA抗體效價略有降低,A=0.191時的稀釋倍數為1∶8 000,1∶4 000 時A=0.235;rLV-HA組A=0.24時的稀釋倍數為1∶2 000。見圖4。
2.3免疫攻毒后小鼠的體重變化初次免疫后,rLV-HA-GCN4組小鼠體重變化較大,在免疫后第8天逐漸上升;加強免疫后,各組小鼠體重差異無統計學意義(P>0.05);攻毒后,rLV-HA-GCN4組小鼠體重在前3 d略有下降,隨后顯著上升,與LV組和PBS組相比,差異有統計學意義(P<0.05),表明rLV-HA-GCN4免疫后能有效保護小鼠抵御病毒的攻擊。見圖5。
2.4脾肺指數攻毒后第14天,LV-HA-GCN4組、rLV-HA組、LV組、PBS組小鼠脾指數分別為3.5±0.66、3.61±0.74、4.92±0.39、5.3±0.18。統計學分析顯示,rLV-HA-GCN4組脾指數與PBS和LV組相比有統計學意義(P<0.05)。rLV-HA-GCN4組、rLV-HA組、LV組、PBS組肺指數分別為0.64±0.05、0.73±0.14、0.8±0.12、1.04±0.2,rLV-HA-GCN4組與PBS組相比,差異有統計學意義(P<0.05)。由此表明,免疫rLV-HA-GCN4對攻毒小鼠脾肺都具有保護作用。見圖6。
3討論
作為流感病毒表面的重要糖蛋白,HA不僅能結合細胞表面的唾液酸受體,而且是病毒最主要的保護性抗原。蛋白的功能取決于結構,所表達蛋白的結構越接近于天然構象,免疫原性越好。為了保持HA的天然三聚體結構,本研究組在前期工作中利用GS-linker將HA與GCN4偶聯,形成HA-GCN4柔性鏈接,以此利于HA的自然折疊,并以慢病毒包裝系統表達了HA-GCN4偶聯蛋白,獲得了主要以三聚體形式存在的HA[3]。本研究以細胞與體液免疫、脾肺指數、體重變化等指標評價了rLV-HA-GCN4的免疫原性及其免疫保護效果。
CD4+和CD8+T細胞是成熟T淋巴細胞的兩個重要亞群。其中,CD4+T細胞通過產生細胞因子或細胞毒活性作用來清除病原,調節炎癥反應,輔助CD8+T細胞活化。CD8+T細胞為細胞毒性細胞或抑制性細胞,能夠發揮特異性的殺傷作用或抑制免疫反應。正常生理情況下,各亞群T淋巴細胞在數量和功能上保持穩定并相互協調,以維持機體的免疫穩態[4]。在本研究中,CD4+和CD8+分型結果顯示,免疫或攻毒后,CD4+T細胞數量均明顯多于CD8+T細胞。但是從免疫到攻毒,CD4+T細胞數量出現由低-高-低的走勢,而CD8+T細胞數量表現為持續升高,表明免疫已經有效激活了T淋巴細胞,而當CD4+T細胞發揮抗病毒作用后,免疫系統可通過促進CD8+T細胞的活化、增殖來調節CD4+和CD8+T細胞的功能平衡,從而有利于免疫系統趨于穩態。
CD4+T細胞是不均一的細胞亞群,主要亞群是輔助性T細胞(helper T lymphocyte,Th)。根據CD4+Th細胞所分泌細胞因子的不同,可進一步將其分為若干亞群,其中Th1與Th2細胞是最早被認識的兩個Th細胞亞型。Th1與Th2之間相互制約,通過遏制彼此的數量及功能而處于動態平衡,以保持機體正常的免疫功能。一旦發生平衡失調,即Th1/Th2漂移,就會導致免疫調節功能的紊亂。免疫功能的平衡主要由不同的T淋巴細胞亞群分泌的各種細胞因子進行調控。其中,Th1細胞以表達IFN-γ、TNF-α、IL-2、IL-12為主,Th2細胞主要分泌IL-4、IL-5、IL-6、IL-10、IL-13[5]。IFN-γ是主要的抗病毒因子,能激活CD8+T細胞、NK細胞及巨噬細胞,增強中性粒細胞的吞噬功能,促進炎性細胞因子(IL-1、IL-12等)的合成和分泌。IL-12是一種多功能中間物質,是介導Th0向Th1方向極化的最核心因子,可與IL-2共同誘導Th1細胞分泌更多的IFN-γ,是強化細胞免疫因子之一[6,7]。TNF-α是免疫與炎癥反應中所分泌的具有抗病毒作用的細胞因子[8]。IL-4是Th2細胞分泌的特征性抗炎細胞因子,對B細胞、T細胞、巨噬細胞等都有免疫調節作用[9]。IL-10也是一種抗炎細胞因子,對恢復及維持體內的免疫平衡有重要作用。它能明顯抑制Th1型細胞因子產生,并間接促進Th2細胞分化。此外,IL-10能夠降低抗原的提呈,抑制巨噬細胞的活性,進而抑制促炎癥因子和趨化因子的產生[10]。在本研究中,IFN-γ、TNF-α、IL-4、IL-10、IL-12五種細胞因子的檢測結果顯示,初次免疫后各組細胞因子含量高于加強免疫,這可能是由于初次免疫時機體首次受到抗原的刺激導致細胞因子在短期內大量釋放,而處于免疫狀態的機體受到同種抗原的再刺激時,不再表現出強烈的免疫反應。根據免疫后細胞因子的表達水平,本研究中收集的各免疫時間點的T淋巴細胞樣本主要表達Th1型細胞因子。
體液免疫檢測結果表明,rLV-HA-GCN4組小鼠在加強免疫后第14天,IgG抗體效價可達1∶8 000,攻毒后第14天約為1∶7 000;攻毒后,rLV-HA-GCN4組脾肺指數顯著低于PBS組,小鼠體重在前3 d略有下降,隨后逐漸上升。
4結論
由于酵母轉錄激活因子——GCN4能夠促進與其偶聯表達的蛋白分子保持天然的多聚體結構,從而有利于蛋白保留其生物學活性,因此利用慢病毒包裝系統構建的rLV-HA-GCN4免疫小鼠后表現出了良好的免疫原性,能夠有效誘導細胞與體液免疫應答,這比單純表達HA的rLV-HA組效果要好。本研究對流感病毒免疫制劑的研制具有一定的借鑒意義。
參考文獻:
[1]Watanabe T,Kawaoka Y.Pathogenesis of the 1918 Pandemic Influenza Virus[J].PLoS Pathogens,2011,7(1):e1001218.
[2]Leung GM,Nicoll A.Reflections on pandemic (H1N1) 2009 and the international response[J].PLoS Med,2010,7(10):e1000346.
[3]李鑫,宋戰昀,朱利塞,等.表達豬H1N1亞型流感病毒三聚體HA的重組慢病毒包裝[J].中國獸醫學報,2012,32(12):1828-1845.
[4]Swain SL,McKinstry KK,Strutt TM.Expanding roles for CD4+T cells in immunity to viruses[J].Nat Rev Immunol,2012,12(2):136-148.
[5]Motomura Y,Kitamura H,Hijikata A,etal.The transcription factor E4BP4 regulates the production of IL-10 and IL-13 in CD4+T cells[J].Nat Immunol,2011,12(5):450-459.
[6]Nazari B,Amirzargar A,Nikbin B,etal.Comparison of the Th1,IFN-γ secreting cells and FoxP3 expression between patients with stable graft function and acute rejection post kidney transplantation[J].Iran J Allergy Asthma Immunol,2013,12(3):262-268.
[7]Méndez-Samperio P.Role of interleukin-12 family cytokines in the cellular response to mycobacterial disease[J].Int J Infect Dis,2010,14(5):e366-e371.
[8]Walsh KB,Teijaro JR,Rosen H,etal.Quelling the storm:utilization of sphingosine-1-phosphate receptor signaling to ameliorate influenza virus-induced cytokine storm[J].Immunol Res,2011,51(1):15-25.
[9]Luzina IG,Keegan AD,Heller NM,etal.Regulation of inflammation by interleukin-4:a review of "alternatives"[J].J Leukoc Biol,2012,92(4):753-764.
[10]Fujio K,Okamura T,Yamamoto K.The family of IL-10-secreting CD4+T cells[J].Adv Immunol,2010,105:99-130.
[收稿2016-01-13修回2016-01-27]
(編輯倪鵬)
doi:10.3969/j.issn.1000-484X.2016.07.011
作者簡介:鄧穎琦(1992年-),女,在讀碩士,主要從事預防獸醫方面研究,E-mail:dengyingqi92@126.com。
通訊作者及指導教師:丁壯(1960年-),男,教授,博士生導師,主要從事人獸共患病毒病診斷與防控研究。
中圖分類號S855.3
文獻標志碼A
文章編號1000-484X(2016)07-0983-05
Study on immunogenicity of recombinant lentivirus expressing trimeric HA of swine H1N1 influenza virus
DENG Ying-Qi,LI Jun-Feng,QU Hui,TANG Yu-Bo,SUN Yi-Xue,LI Xin,WANG Wei-Li,DING Zhuang,CONG Yan-Long.
College of Veterinary Medicine,Jilin University,Changchun 130062,China
[Abstract]Objective:To evaluate the protective immunity by vaccination of BALB/c mice with rLV-HA-GCN4,a recombinant lentivirus expressing the trimeric HA of swine H1N1 influenza virus.Methods: The female mice were randomly divided into rLV-HA-GCN4,rLV-HA,LV and PBS groups.Mice were primed with plasmid and boosted with lentivirus by the administration of intramuscular thigh injections at an interval of two weeks.At day 28 post-prime immunization,mice were inoculated intranasally with 100TCID50of swine H1N1 influenza virus in a 50 μl volume.The immune levels were assessed by the T lymphocyte transformation test,flow cytometry,indirect ELISA and the indexes of spleen and lung.Results: The spleen lymphocyte transformation rate was 0.3±0.11 in the rLV-HA-GCN4 group at day 14 post-boost immunization,showing a statistical significance (P<0.01) compared to the PBS group.Meanwhile,rLV-HA-GCN4 could cause T lymphocyte response mainly based on the Th1-type CD4+ T cells.The IgG antibody titer reached to 1∶8 000 at day 14 post-boost immunization and approximately 1∶7 000 at day 14 post challenge.After challenge,the spleen and lung indexes of rLV-HA-GCN4 group were significantly lower than those of PBS group (P<0.05).The body weight of rLV-HA-GCN4 group demonstrated a slight decrease before 3 days post challenge and then a gradual increase compared to the LV and PBS groups (P<0.05).Conclusion: rLV-HA-GCN4 can effectively induce cellular and humoral immune response in BALB/c mice against swine H1N1 influenza virus.
[Key words]Swine influenza; H1N1 subtype; Trimeric HA; Lentiviral vector; Immunogenicity
①本文為國家質檢總局科技計劃項目(2014IK237)和吉林省科技廳重點科技攻關項目(20150204029NY)。
②共同第一作者。
③吉林農業大學校醫院,長春130118。
④吉林省動植物檢驗檢疫局,長春130062。
⑤吉林農業大學動物科學技術學院,長春130118。
⑥吉林省出入境檢驗檢疫局,長春130000。
叢彥龍(1977年-),男,教授,碩士生導師,主要從事人獸共患病毒病診斷與防控研究。
