發酵種泥的投加對新鮮剩余污泥發酵產酸的影響
2016-08-08 06:56:22彭永臻王淑瑩
哈爾濱工業大學學報 2016年8期
袁 悅,彭永臻,劉 曄,王淑瑩
(北京工業大學 北京市水質科學與水環境恢復工程重點實驗室,北京市污水脫氮除磷處理與過程控制工程技術研究中心,北京100124)
?
發酵種泥的投加對新鮮剩余污泥發酵產酸的影響
袁悅,彭永臻,劉曄,王淑瑩
(北京工業大學 北京市水質科學與水環境恢復工程重點實驗室,北京市污水脫氮除磷處理與過程控制工程技術研究中心,北京100124)
摘要:為進一步提高剩余污泥厭氧發酵產短鏈脂肪酸(SCFAs)的效率,將pH=10下長期(90 d)厭氧發酵的種泥接入至新鮮剩余污泥考察其對新鮮剩余污泥厭氧發酵產酸的影響.實驗在2種pH條件下進行:不控制pH (pH=7.4±0.2),一組反應器中投加新鮮剩余污泥,并接種厭氧發酵種泥,另一組反應器只投加新鮮剩余污泥;控制pH為堿性 (pH=10±0.2),一組反應器中投加新鮮剩余污泥,并接種厭氧發酵種泥,另一組反應器只投加新鮮剩余污泥.結果表明:相同pH條件下,接種厭氧發酵種泥反應器中的最大產酸量較不接種分別提高4.9(pH=7.4±0.2) 和16.4 mg /g VSS(以COD計) (pH=10±0.2);同時接種發酵污泥、剩余污泥在pH=10±0.2下的最大產酸量較pH=7.4±0.2下的最大產酸量高146.2 mg/g VSS;堿性條件、不接種發酵種泥的產酸量較不控制pH、接種發酵種泥的高129.8 mg/g VSS.投加發酵種泥可提高新鮮剩余污泥厭氧發酵過程中SCFAs的產生效率;堿性條件與厭氧發酵種泥對新鮮剩余污泥發酵產酸具有協同促進作用;與接種發酵種泥相比,堿性條件對新鮮剩余污泥厭氧發酵產酸促進能力高.
關鍵詞:發酵種泥; 剩余污泥; 厭氧發酵; 短鏈脂肪酸; pH值
目前,城市污水處理廠多采用活性污泥法去除生活污水中的有機物、氮和磷,然而,傳統活性污泥法會產生大量的污泥,而處理這些污泥的費用占水廠總運行費用的60%[1].因此,為降低污泥處理成本,需對污泥進行減量化.污泥減量化處理常用的方法有厭氧消化和好氧消化[2-4].其中厭氧消化又以能產生甲烷氣體、氫氣,活性污泥法脫氮除磷的重要有機碳源——短鏈脂肪酸(SCFAs)等能源質引起廣泛關注[5-9].城市生活污水的碳氮比、碳磷比較低,污水處理廠為了提高脫氮除磷效果,需要外加碳源,方法之一是直接加入化學制成的碳源,如乙酸鈉、乙醇等,該方法無疑增加了污水處理廠的處理成本.近年來,有文獻報道污泥厭氧發酵不僅可以達到污泥減量,還可以獲得大量的易生物降解碳源——SCFAs[10-11]. 污泥厭氧消化包括3個階段:水解、酸化和產甲烷.其中水解是整個消化過程的限制性過程[12-13].從產酸的角度出發,提高水解過程并且抑制產甲烷過程才能使得SCFAs積累.研究表明,污泥在堿性條件下發酵產酸量比酸性或中性時高得多[14-15],但是達到最大SCFAs產量的發酵時間仍需要8~10 d.
厭氧條件下污泥的水解、酸化和產甲烷各過程是通過大量的細菌或者古菌實現的.因此,污泥厭氧發酵過程中微生物起著很重要的作用[16-18].該過程引進水解菌、發酵菌或許會進一步提高剩余污泥產酸量,縮短發酵時間.接偉光等[19]通過分離剩余污泥中的耐堿性菌株HIT-01和HIT-02,并將2種細菌單獨或者同時投加到新鮮污泥中,觀察其在堿性條件下的產酸量情況.結果表明,接種耐堿性菌株可以增加剩余污泥發酵產酸,同時接種2種菌株時SCFAs積累量最多.然而該研究依然控制整個發酵過程為堿性條件,未考查不控制pH(即中性)的情況,并且投加的菌種是通過分離純培養得到的,操作復雜. 基于以上研究,將pH=10下長期發酵的污泥作為種泥接至新鮮剩余污泥中,在不同條件下(堿性條件、不控制pH條件)考察其對新鮮剩余污泥厭氧發酵過程中產生SCFAs的影響,并進一步從產酸所需的底物(蛋白質和多糖)方面分析不同條件下影響產SCFAs的原因.
1實驗
1.1剩余污泥
實驗所用新鮮剩余污泥均取自以實際生活污水為處理對象的中試SBR (有效體積6 m3).此SBR 的1個運行周期為12 h,運行方式為:進水(0.25 h)、好氧(6 h)、缺氧(1 h)、厭氧(2 h)、沉淀(2.25 h)、排水(0. 5 h).其中好氧段主要是短程硝化,缺氧段則是短程反硝化,剩余污泥在厭氧末端排出.排出的污泥在4 ℃下沉淀濃縮,濃縮后污泥的主要成分如表1所示.

表1 實驗用剩余污泥性質
注:TSS為污泥中懸浮物固體濃度, VSS為污泥中揮發性懸浮物固體濃度, TCOD為剩余污泥總COD, SCOD為可溶性COD, SCFAs為短鏈脂肪酸.
1.2實驗方法
1.2.1厭氧發酵種泥的馴化
采用5 L的半連續厭氧發酵反應器(SAFR).控制污泥發酵的pH在10±0.2,溫度為(30±2)℃,污泥停留時間SRT為6 d,即每天排833 mL發酵污泥,并加入等體積(833 mL)的新鮮剩余污泥至SAFR中.該反應器的污泥來源為上述新鮮剩余污泥,運行30 d后監測其產酸量,在90 d時測定發酵污泥的微生物菌群結構.
1.2.2厭氧發酵種泥對剩余污泥發酵產酸的影響
取SAFR中的厭氧發酵種泥1 200 mL,離心棄上清液,再用蒸餾水清洗離心2次.本實驗 共有4組反應器:1)不控制pH(pH=7.4±0.2),投加新鮮剩余污泥(1#);2)不控制pH(pH=7.4±0.2),投加新鮮剩余污泥,并接種厭氧發酵種泥(2#);3)控制pH(pH=10±0.2),投加新鮮剩余污泥(3#);4)控制pH(pH=10±0.2),投加新鮮剩余污泥,并接種厭氧發酵種泥(4#).每組反應器設有2個平行實驗 ,將4組反應器置于磁力攪拌器上,轉速為120 r/min,溫度θ在(30±2) ℃.每組反應器吹氮氣3 min,去除反應器中的溶解氧,保持厭氧環境.反應期間,為了保證各反應器的pH在10±0.2,用2 mol·L-1的NaOH和2 mol·L-1HCl調節.取樣周期為1 d,取出的水樣在轉速為4 000 r/min的離心機中離心10 min,然后用0.45 μm的濾膜抽濾,濾液用來分析各項指標,過濾后的水樣立即測定,最終數據為每組平行實驗的平均值.
1.3DNA提取、PCR擴增和Miseq測序
在SAFR運行到第90 天并且投加新鮮剩余污泥前取發酵污泥.首先,泥樣在轉速為4 000 r/min的離心機中離心5 min,棄上清液,然后將泥樣放置冷凍干燥機進行冷凍干燥.稱取0.034 6 g干污泥,用DNA試劑盒(FastDNA Spin Kit for Soil)提取DNA.設計的引物用于擴增16S rRNA基因的V3+V4區[20],引物系列見表2. PCR反應體系(20 μL)如下:5×FastPfu Buffer,4 μL;2.5 mmol/L dNTPs,2 μL;Forward Primer (5 μmol/L),0.4 μL;Reverse Primer (5 μmol/L),0.4 μL; FastPfu Polymerase,0.4 μL;Template DNA,10 ng;ddH2O,20 μL.
PCR擴增條件如下:95 ℃預變性2 min;然后,95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸45 s,共25個循環;72 ℃最終延伸10 min,最終4 ℃保存.Illumina Miseq測序平臺為PE300,測得的原始數據可在NCBI數據庫通過檢索號SRA230879獲得,序列的基本信息見表3.
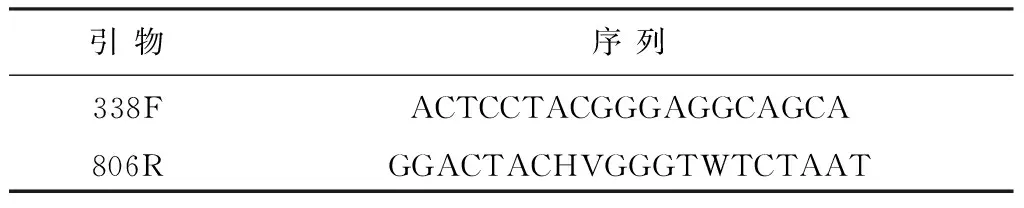
表2 PCR擴增所用引物

表3 Illumina Miseq測序序列信息
注:OUT為Operational taxonomic units, 相似水平為0.97.
1.4分析方法
TSS、VSS采用稱重法測定.TCOD、SCOD采用5B-3(B)型COD快速測儀,碳水化合物采用苯酚-硫酸比色法;蛋白質采用Lowry-Folin試劑.SCFAs采用Agilent7890A氣相色譜儀測定,FID檢測器,色譜柱型號及尺寸:AgilentdB-WAXetr, 30 m×0.53 mm×0.001 mm,N2為載氣,載氣流量為20 mL·min-1,進樣口和檢測器分別維持在220和250 ℃,烘箱起始溫度為80 ℃,最后升溫至240 ℃,進樣體積2 μL.實驗 中檢測到的SCFAs包括乙酸、丙酸、正丁酸、異丁酸、正戊酸和異戊酸,將其濃度分別乘以比例系數1.07、1. 51、1. 82、1.82、2.04、2.04轉化為COD 濃度[21]后的加合計為總SCFAs濃度.采用WTWpH/Oxi340i測定儀測定pH.
2結果與討論
2.1SAFR中的SCFAs產量及微生物菌群
如圖1所示,SAFR反應器在運行30 d后,經過60 d的穩定運行,其平均產酸量為(1 721.4±91.2) mg/L. Illumina Miseq測序表明:在phylum水平上分析可得到10種細菌(圖2),其中Chloroflexi, Proteobacteria, Bacteroidetes和Firmicute是主要的菌屬,Chloroflexi為SAFR中最豐富的菌屬,占30.1%[22].為了進一步確定檢測的細菌在污泥發酵過程所起的作用,又從genus水平上對Illumina Miseq測序結果進行分析,見表4.水解細菌——Alcaligenes(3.47%)和Anaerolinea(3.34%)、產酸細菌——Erysipelothrix(7.31%),Paludibacter(4.96%)和Tissierella(3.59%)分別得到富集[22].
因此,綜合SAFR反應器穩定的產酸性及富有水解菌和產酸菌,其發酵污泥可用作種泥.
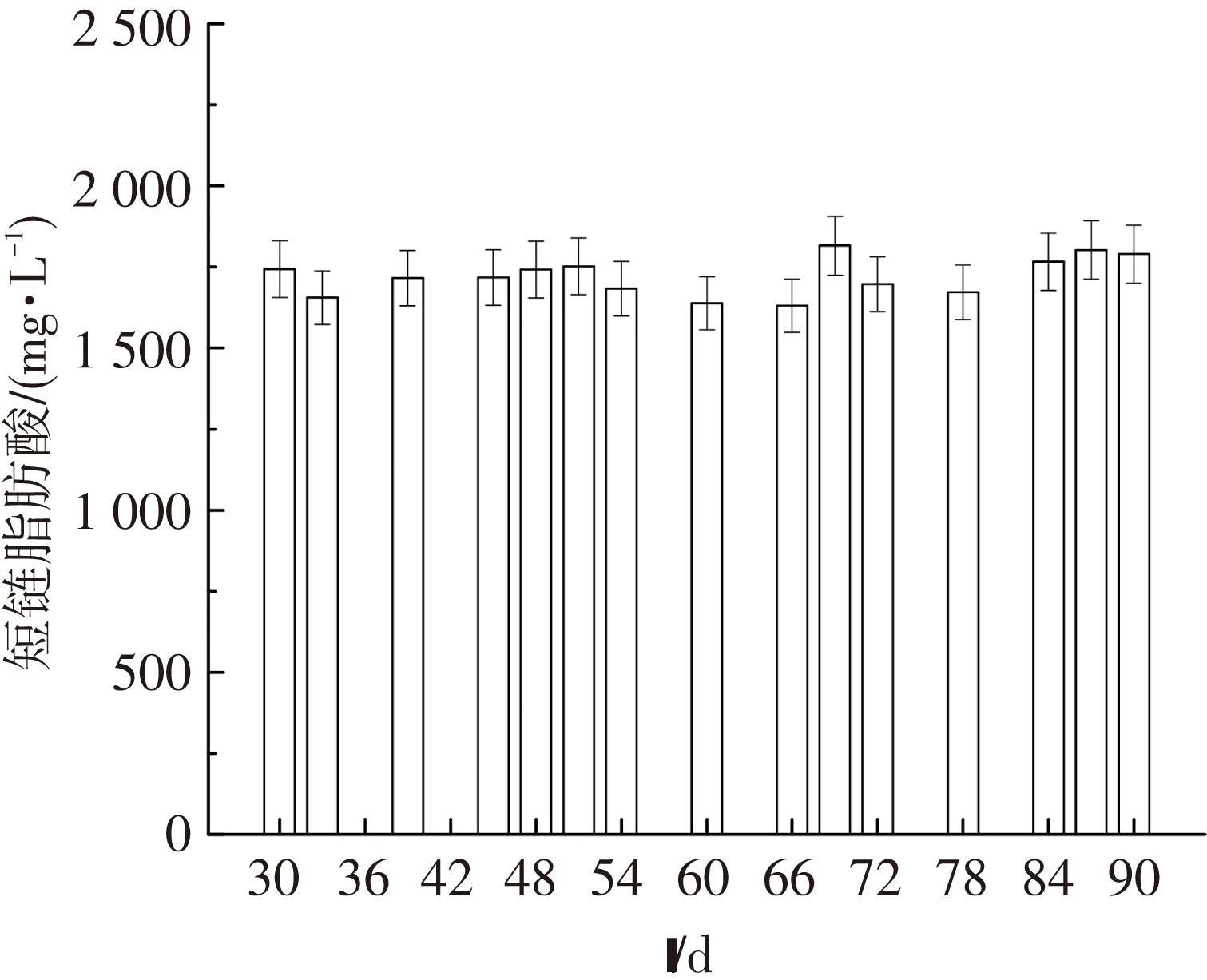
圖1 SAFR中隨時間變化的產酸量
Fig.1SCFAs production from excess sludge with the fermentation time
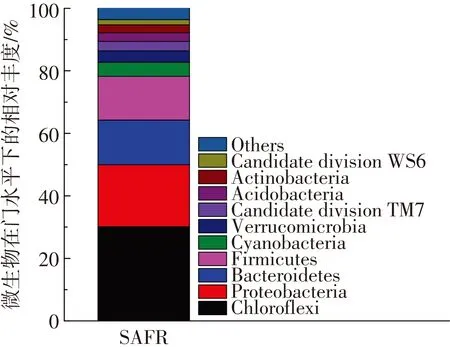
圖2 SAFR中微生物在門水平上的相對豐度
表4SAFR中16 s rRNA基因序列的系統分類(屬的水平, 相對豐度>1%)
Tab.4Phylogenetic classification of the 16 s rRNA gene sequences (relative abundance>1% at genus level) at SAFR
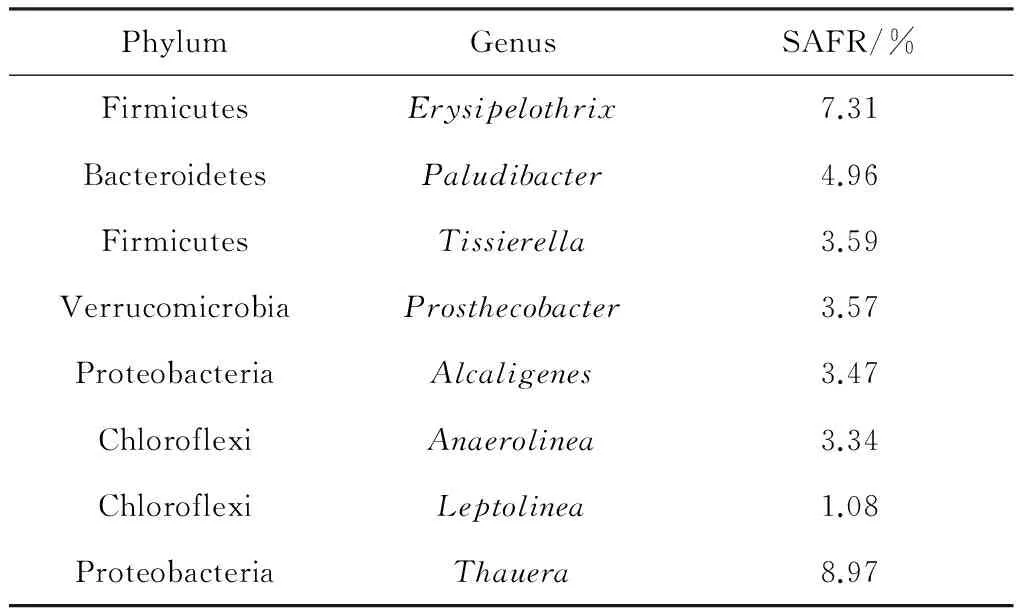
PhylumGenusSAFR/%FirmicutesErysipelothrix7.31BacteroidetesPaludibacter4.96FirmicutesTissierella3.59VerrucomicrobiaProsthecobacter3.57ProteobacteriaAlcaligenes3.47ChloroflexiAnaerolinea3.34ChloroflexiLeptolinea1.08ProteobacteriaThauera8.97
2.2剩余污泥厭氧發酵產生的SCFAs
圖3表明新鮮剩余污泥在不同條件下產短鏈脂肪酸(SCFAs)的情況.發酵前期,1#(1~9 d)、2#(1~9 d)、3#(1~10 d)、4#(1~6 d)反應器的產酸量均呈現增長的趨勢.而反應后期,1#、2#、4#呈下降趨勢,3#則趨于穩定.這是因為1#、2#反應器的pH均是中性,隨著發酵時間的進行,產甲烷菌出現,消耗了SCFAs.而3#反應器的堿性條件短時間內抑制了產甲烷菌的生長,4#反應器雖然是堿性條件,但是接種的厭氧發酵種泥中含有極少量的產甲烷菌[17、22],待適應了厭氧發酵環境后便消耗SCFAs.因此,為了獲取不同條件下的最大產酸量,需要控制各自的SRT,即1#~9 d、2#~9 d、3#~10 d、4#~6 d.
從圖3還可以看出,在發酵第6 天,4#反應器(控制pH=10±0.2,并接種厭氧發酵種泥)產酸量達187.2 mg /g VSS,而1#、2#、3#反應器的最大產酸量分別是36.1 (9 d)、41.0 (9 d)和170.8 mg /g VSS(10 d).這是因為堿性條件利于剩余污泥發酵產酸[14-15],其次接種的厭氧發酵種泥引入了產酸菌, 提高了產酸效率, 將最大產酸量的時間提前了3~4 d.另外,在pH不控的條件下,投加種泥的最大產酸量(2#)較不投加種泥的最大產酸量(1#)提高了4.9 mg/g VSS(顯著性P<0.05);在堿性條件下,投加種泥(4#)較不投加種泥的最大產酸量(3#)提高了16.4 mg/g VSS.這說明投加發酵種泥可提高新鮮剩余污泥厭氧發酵過程中SCFAs的產生效率,并且堿性條件及厭氧發酵種泥對剩余污泥厭氧發酵產酸具有協同促進作用.由于堿性條件利于剩余污泥發酵產酸[14-15],均接種發酵污泥的剩余污泥在pH=10±0.2下的最大產酸量較pH=7.4±0.2的高146.2 mg /g VSS.此外,相對于2#反應器,3#反應器的最大產酸量提高了129.8 mg/g VSS,這表明堿性條件較厭氧發酵種泥對新鮮剩余污泥厭氧發酵產酸更具促進作用.
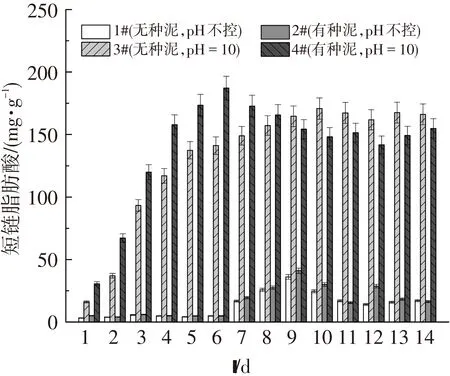
圖3 4組反應器中短鏈脂肪酸產量
2.3剩余污泥厭氧發酵產生的溶解性蛋白質和多糖
污泥胞外聚合物(extracellular polymeric substances, EPS)是附著于微生物細胞壁上的大分子有機多聚物,主要來源為進水基質、微生物新陳代謝和細胞自溶,是產生SCFAs所需要的主要基質.EPS具有復雜的化學組成,占總量70%~80%的蛋白質和多糖是最主要的2種成分,而核酸、腐殖質、糖醛酸、脂類和氨基酸等的含量相對較低[23].因此,本文主要研究了溶解性蛋白質和多糖,從產酸所需的基質進一步分析剩余污泥在不同條件下產酸量不同的原因.
圖4、5表明,溶解性的多糖濃度低于蛋白質的濃度,這與污泥中蛋白質含量高于多糖有關.另外,溶解性的多糖濃度變化較蛋白質的波動大.相對于中性條件,溶解性蛋白質在堿性條件下的溶出量較高,這是因為堿性導致EPS中的酸性基團分離,使得帶負電的EPS相互排斥,進而增加蛋白質和多糖的溶解性[24].然而接種了厭氧發酵種泥的4#反應器又比3#的溶解性蛋白質高,這是因為接種種泥引入了水解菌,這些細菌會釋放胞外酶[23],進而促進了蛋白質、多糖的釋放.
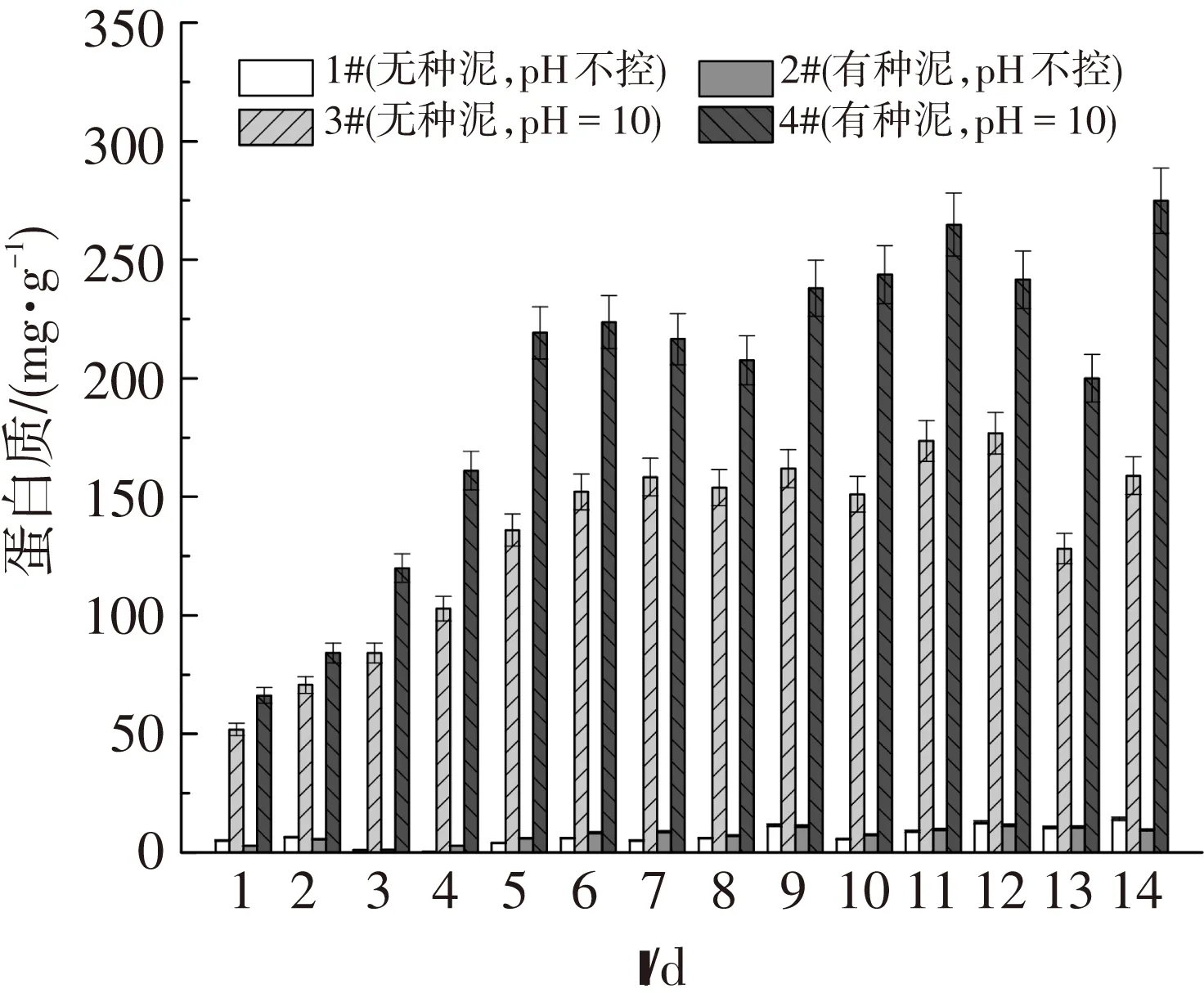
圖4 4組反應器中溶解性蛋白質的釋放量
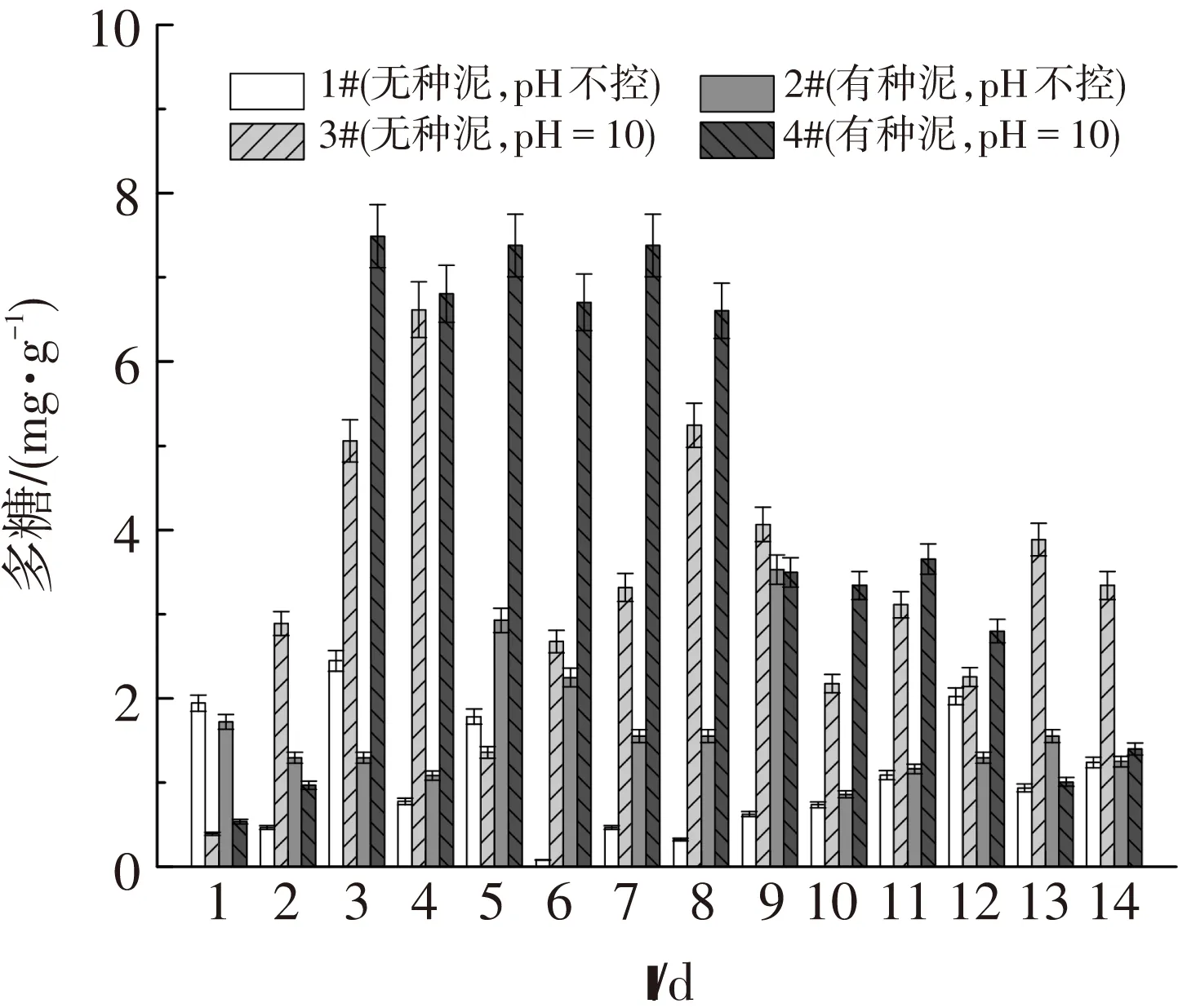
圖5 4組反應器中溶解性多糖的釋放量
短鏈脂肪酸是產酸菌利用溶解性蛋白質和多糖的水解產物(氨基酸、單糖)生成的,而蛋白質的水解伴隨有NH4+-N的生成.因此,發酵液中NH4+-N的釋放可以表征溶解性蛋白質的水解作用[25].由圖6可以看出,隨著發酵時間不斷增加,4組反應器發酵液中的NH4+-N含量逐步升高,這表明溶解性有機物不斷水解生成小分子有機物,為產酸菌發酵產酸提供了所需的基質.4組反應器的NH4+-N含量高低順序為4#>3#>2#>1#.顯然4#反應器的產酸基質最充足,這與產酸量最多相吻合.另外,堿性條件抑制產甲烷菌的活性,也造就了SCFAs的積累.因此,接入的產酸菌、足夠的發酵產酸所需基質以及產甲烷過程受抑制是4#反應器產酸效率高的原因.
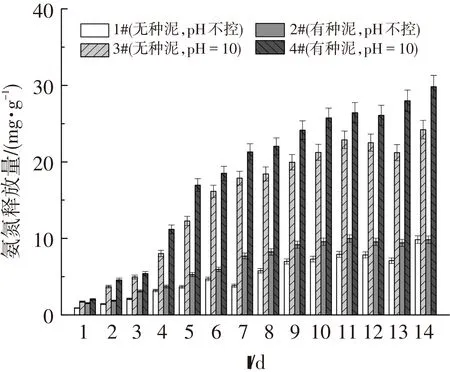
圖6 4組反應器中氨氮(NH4+-N)的釋放量
3結論
1)投加厭氧發酵種泥提高了剩余污泥厭氧發酵產短鏈脂肪酸的效率.在pH不控的條件下,投加種泥的產酸量較不投加提高了4.9 mg/g VSS;在堿性pH條件下,投加種泥較不投加種泥的產酸量提高了16.4 mg /g VSS. 接入的產酸菌以及足夠的發酵產酸所需基質是產酸效率高的原因.
2)種泥的投加與堿性條件對剩余污泥發酵產酸具有協同促進作用.
3)堿性條件比投加種泥對剩余污泥發酵產酸更具促進作用.
參考文獻
[1] WEI Yuansong, VAN-HOUTEN R T, BORGER A R, et al. Minimization of excess sludge production for biological wastewater treatment [J]. Water Research, 2003, 37(18):4453-4467.
[2] FOLADORI P, ANDREOTTOLA G, ZIGLIO G. Sludge reduction technologies in wastewater treatment plants [M]. London: IWA Publishing, 2010.
[3] ZHANG Dong, CHEN Yinguang, ZHAO Yuxiao, et al. New sludge pretreatment method to improve methane production in waste activated sludge digestion [J]. Environmental Science and Technology, 2010, 44 (12):4802-4808.
[5] YAN Yuanyuan, CHEN Hanlong, XU Wenying, et al. Enhancement of biochemical methane potential from excess sludge with low organic content by mild thermal pretreatment[J]. Biochemical Engineering Journal, 2013, 70:127-134.
[6] GUO Liang, LI Xiaoming, ZENG Guangming, et al. Effective hydrogen production using waste sludge and its filtrate [J]. Energy, 2010, 35(9): 3557-3562.
[7] LU Lu, XING Defeng, LIU Bingfeng, et al. Enhanced hydrogen production from waste activated sludge by cascade utilization of organic matter in microbial [J]. Water Research, 2012, 46(4):1015-1026.
[8] JIANG Su, CHEN Yinguang, ZHOU Qi, et al. Biological short-chain fatty acids (SCFAs) production from waste-activated sludge affected by surfactant [J]. Water Research, 2007, 41: 3112-3120.
[9] LIU He, WANG Jin, LIU Xiaoling, et al. Acidogenic fermentation of proteinaceous sewage sludge: effect of pH [J]. Water Research, 2012, 46(3):799-807.
[10]TONG Juan, CHEN Yinguang. Enhanced biological phosphorus removal driven by short-chain fatty acids produced from waste activated sludge alkaline fermentation [J]. Environmental Science and Technology, 2007, 41(20):7126-7130.
[11]GAO Yongqing, PENG Yongzhen, ZHANG Jingyu, et al. Biological sludge reduction and enhanced nutrient removal in a pilot-scale system with 2-step sludge alkaline fermentation and A(2)O process [J]. Bioresource Technology, 2011, 102(5):4091-4097.
[12]KIM M, GOMEC C Y, AHN Y, et al. Hydrolysis and acidogenesis of particulate organic material in mesophilic and thermophphilic anaerobic digestion [J]. Environmental Technology, 2003, 24(9):1183-1190.
[13]BOUSKOVA A, DOHANYOS M, SCHMIDT J E, et al. Strategies for changing temperature from mesophilic to thermophilic conditions in anaerobic CSTR reactors treating sewage sludge [J].Water Research, 2005,39:1481-1488.
[14]CHEN Yinguang, JIANG Su, YUAN Hongying, et al. Hydrolysis and acidification of waste activated sludge at different pHs [J]. Water Research, 2007, 41(3): 683-689.
[15]JIE Weiguang, PENG Yongzhen, REN Nanqi, et al. Volatile fatty acids (VFAs) accumulation and microbial community structure of excess sludge (ES) at different pHs [J]. Bioresource Technology,2014, 152:124-129.
[16]RIVIERE D, DESVIGNES V, PELLETIER E, et al. Towards the definition of a core of microorganisms involved in anaerobic digestion of sludge [J]. International Society for Microbial Ecology, 2009, 3(6):700-714.
[17]ZHENG Xiong, SU Yinglong, LI Xiang, et al. Pyrosequencing reveals the key microorganisms involved in sludge alkaline fermentation for efficient short-chain fatty acids production[J]. Environmental Science and Technology, 2013, 47(9):4262-4268.
[18]LI Xiaoling, PENG Yongzhen, REN Nanqi, et al. Effect of temperature on short chain fatty acids (SCFAs) accumulation and microbiological transformation in sludge alkaline fermentation with Ca(OH)2adjustment [J].Water Research, 2014, 61:34-45.
[19]JIE Weiguang, PENG Yongzhen, REN Nanqi, et al. Utilization of alkali-tolerant stains in fermentation of excess sludge [J]. Bioresource Technology, 2014, 157:52-59.
[20]DENNIS K L, WANG Y, BLATNER N R, et al. Adenomatous polyps are driven by microbe-instigated focal inflammation and are controlled by IL-10-producing T cells [J]. Cancer Research, 2013, 73(19):5905-5913.
[21]EASTMAN J A, FERGUSON J F. Solubilization of particulate organic carbon during the acid phase of anaerobic digestion [J]. Journal of the Water Pollution Control Federation, 1981, 53(3): 352-366.
[22]YUAN Yue, WANG Shuying, LIU Ye, et al. Long-term effect of pH on short-chain fatty acids accumulation and microbial community in sludge fermentation systems[J]. Bioresource Technology, 2015,197: 56-63.
[23]FROLUND B, PALMGREN R, KEIDING K, et al. Extraction of extracellular polymers from activated sludge using a cation exchange resin [J]. Water Research, 1996, 30(8):1749-1758.
[24]WINGENDER J, NEU T R, FLEMMING H C. Microbial extracellular polymeric substances: characterisation, structure and function [M]. Berlin:Springer, 1999.
[25]PIJUAN M, WANG Qilin, YE Liu, et al. Improving secondary sludge biodegradability using free nitrous acid treatment [J]. Bioresource Technology, 2012,116: 92-98.
(編輯劉彤)
doi:10.11918/j.issn.0367-6234.2016.08.006
收稿日期:2016-02-18
基金項目:國家自然科學基金(51578014); 北京市教委資助項目
作者簡介:袁悅(1988—), 女, 博士;
通信作者:彭永臻, pyz@bjut.edu.cn
中圖分類號:X703
文獻標志碼:A
文章編號:0367-6234(2016)08-0037-05
Effect of fermented sludge on short-chain fatty acids (SCFAs) production from excess sludge
YUAN Yue, PENG Yongzhen, LIU Ye, WANG Shuying
(Key Laboratory of Beijing for Water Quality Science and Water Environment Recovery Engineering,Engineering Research Center of Beijing, Beijing University of Technology, Beijing 100124, China)
Abstract:To enhance the production efficiency of short chain fatty acids (SCFAs) in anaerobic sludge fermentation reactor, sludge from another stabilized fermentation reactor (over 90 days) under pH 10 was seeded into the reactors to examine its effect on SCFAs production. Two sets of experiment were designed to investigate the SCFAs production with/without seeding sludge under pH=7.4±0.2 and pH=10±0.2, respectively. Experimental results showed that (1) under same pH condition, the maximum SCFAs accumulation with seeding sludge additive increased by 4.9 mg COD/g VSS (pH=7.4±0.2) and 16.4 mg COD/g VSS (pH=10±0.2) respectively, in comparison with the control reactors; (2) with two reactors with seeding sludge, the maximum SCFAs accumulation at pH 10±0.2 increased by 146.2 mg COD/g VSS as compared with the condition of pH=7.4±0.2; (3) the maximum SCFAs accumulation at pH 10±0.2 without seeding sludge was 129.8 mg COD/g VSS higher than that of pH 7.4±0.2 with seeding sludge. Overall, seeding sludge improved SCFAs production efficiency, meanwhile, seeding sludge under alkaline pH could play synergetic role in enhancing the SCFAs accumulation.
Keywords:fermented sludge; excess sludge; anaerobic fermentation; short-chain fatty acids; pH value
彭永臻(1949—), 男, 博士生導師,中國工程院院士
