天童常綠闊葉林砍伐后凋落物層跳蟲群落生態(tài)學(xué)研究
2016-08-06 02:33:01靳亞麗
安徽農(nóng)業(yè)科學(xué) 2016年16期
靳亞麗, 蔣 躍
(1.上海自然博物館自然史研究中心,上海科技館,上海 200127;2.華東師范大學(xué)生態(tài)與環(huán)境科學(xué)學(xué)院,上海 200062)
?
天童常綠闊葉林砍伐后凋落物層跳蟲群落生態(tài)學(xué)研究
靳亞麗1, 蔣 躍2
(1.上海自然博物館自然史研究中心,上海科技館,上海 200127;2.華東師范大學(xué)生態(tài)與環(huán)境科學(xué)學(xué)院,上海 200062)
摘要[目的]了解不同砍伐處理對常綠闊葉林凋落物層跳蟲群落結(jié)構(gòu)的影響以及各處理樣地跳蟲群落恢復(fù)狀況。[方法]于2009年12月至2010年9月在浙江天童地區(qū)常綠闊葉林5個砍伐7年后的恢復(fù)樣地進行四季采樣,對浙江森林公園不同砍伐處理下常綠闊葉林土壤動物優(yōu)勢類群跳蟲群落進行調(diào)查。樣地處理分別為:(Ⅰ)去除大樹保留萌枝、(Ⅱ)去除地表植物保留枯枝落葉層、(Ⅲ)去除地表植物和10 cm表土、(Ⅳ)清除下木層保留大樹、(Ⅴ)對照。[結(jié)果]各處理樣地各科跳蟲類群組成相差不大,密度從大到小依次為Ⅱ號樣地(28.79%)、Ⅳ號樣地(21.03%)、Ⅰ號樣地(20.75%)、Ⅴ號樣地(17.09%)、Ⅲ號樣地(12.33%),表現(xiàn)為適度地干擾處理能夠讓跳蟲保持較高密度。不同季節(jié)跳蟲群落組成不同,且跳蟲密度在不同樣地季節(jié)變化存在差異,總體上夏秋季明顯高于春冬季。不同砍伐程度下,各樣地凋落物有機碳、總氮和總磷均存在顯著差異,隨著砍伐程度的加劇,凋落物有機碳和總氮含量逐漸降低。結(jié)合不同砍伐程度處理中跳蟲群落密度,雖然各處理樣地經(jīng)過了7年的恢復(fù),但不同處理間跳蟲群落差異仍較為明顯。[結(jié)論]跳蟲群落對環(huán)境變化非常敏感,可作為評價土壤環(huán)境受干擾程度的指示種。
關(guān)鍵詞干擾;跳蟲;凋落物層;常綠闊葉林;天童
跳蟲是天童森林生態(tài)系統(tǒng)凋落物層土壤動物中的優(yōu)勢類群,它們在自然界物質(zhì)循環(huán)[1]、改善土壤理化特性、維護土壤生物群落穩(wěn)定[2]以及指示環(huán)境變化[3]等方面有重要作用,尤其在落葉分解過程中[4]跳蟲起著重要的輔助作用[5-6]。對于森林干擾,跳蟲能夠在密度和多樣性等方面迅速做出反應(yīng)[7-8]。森林砍伐是森林干擾的一種形式,對林地和周邊地區(qū)的生態(tài)環(huán)境產(chǎn)生較大影響,直接影響森林生態(tài)系統(tǒng)的結(jié)構(gòu)功能和演替過程[9],從而影響其中的生物成分。陳小鳥等[10]在不同程度砍伐的影響下常綠闊葉林中的土壤動物群落特征的研究中發(fā)現(xiàn)土壤動物群落受砍伐的影響明顯,且砍伐強度越大,其受影響越大。易蘭等[11-12]于2003年9月至2004年10月對砍伐樣地進行了砍伐前、砍伐后1月、砍伐后1年對這5個樣地中的土壤動物群落進行了跟蹤調(diào)查。陳小鳥[13]于2007年7月至2008年4月對以上砍伐樣地凋落物層和土層的土壤動物進行了進一步跟蹤調(diào)查。筆者對浙江國家森林公園不同砍伐處理下常綠闊葉林土壤動物優(yōu)勢類群跳蟲群落進行了調(diào)查,探討它們的群落特征及其對不同干擾強度的響應(yīng)和恢復(fù)狀況,旨在為不同干擾處理下跳蟲的生態(tài)學(xué)研究提供資料。
1材料與方法
1.1研究區(qū)概況研究區(qū)位于浙江省鄞縣東南部天童國家森林公園(121°47′E,28°48′N),海拔260 m,坡度20°~ 30°,屬于典型的亞熱帶季風(fēng)氣候,年平均溫度為16.2 ℃,年平均降雨量為1 374.7 mm,年平均相對濕度達83%[14]。此次調(diào)查參考宋永昌等[14]2003年設(shè)置的5個20 m×20 m樣地,其中樣地Ⅰ、Ⅱ、Ⅲ位于山路上方,樣地間距為5 m,樣地Ⅳ、Ⅴ號位于山路下方,二者之間相距約100 m(圖1)。2003年10月,模擬當(dāng)?shù)爻R姷娜藶楦蓴_,對其中的4塊樣地進行了不同程度的采伐處理。5個樣地采伐情況和經(jīng)過6年恢復(fù)各樣地優(yōu)勢植物如下:樣地Ⅰ優(yōu)勢植物有木荷(Schimasuperba)、米櫧(Castanopsiscarlesii)、石櫟(Lithocarpusglaber)、格藥柃(Euryamuricata)、毛柄連蕊茶(Camelliafraterna)、老鼠矢(Symplocosstellaris)、山礬(S.sumuntia)、四川山礬(S.setchuensis)、海桐山礬(S.heishanensis)和窄基紅褐柃(Euryarubiginosavar.attenuata);樣地Ⅱ優(yōu)勢植物有木荷、米櫧、茅栗 (Castaneaseguinii)、栲樹 (Castanopsisfargesii)、小葉青岡 (Cyclobalanopsismyrsinifolia)、石櫟、細葉青岡 (C.gracilis)、楊梅 (Myricarubra)、格藥柃和毛柄連蕊茶;樣地Ⅲ優(yōu)勢植物為木荷、米櫧、細葉青岡、石櫟、楓香 (Liquidambarformosana)、苦櫧(Castanopsissclerophylla)、楊梅、老鼠矢、格藥柃和栲樹;樣地Ⅳ優(yōu)勢植物有木荷、馬尾松 (Pinusmassoniana)、石櫟、米櫧、茅栗、細葉青岡、楓香、海桐山礬、苦櫧和東南石櫟;樣地Ⅴ優(yōu)勢植物有木荷、石櫟、米櫧、青岡、楓香、白櫟(Quercusfabric)、苦櫧、老鼠矢、花櫚木和格藥柃。
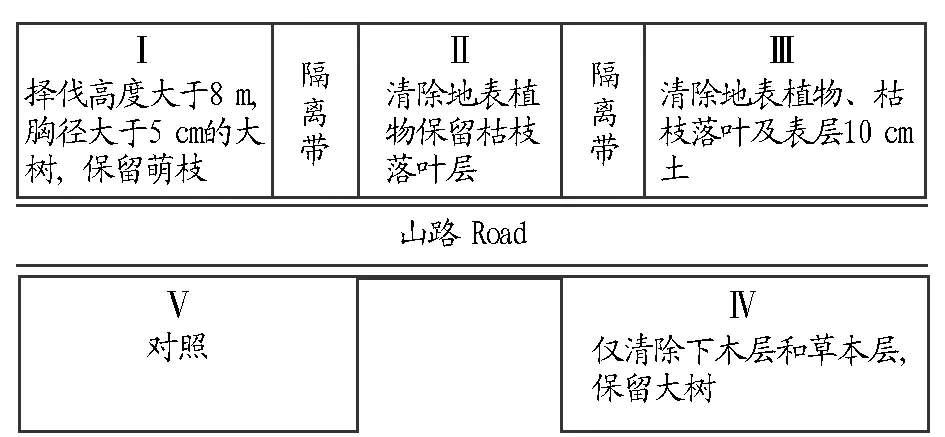
圖1 樣地設(shè)置示意Fig.1 The distribution of the experimental plots
1.2 研究方法對5個樣地進行四季采樣,每個樣地隨機取5個點,樣點之間相距5 m以上,每個點手撿35 cm× 35 cm方形樣方凋落物于布袋中。采集的凋落物樣品帶回室內(nèi),采用Tulltern法分離48 h[15]。
對分離得到的跳蟲進行分類鑒定和數(shù)量統(tǒng)計,采用Bellinger、Christiansen和Janssens等共同修改的最新跳蟲分類系統(tǒng)-“彈尾綱4目分類系統(tǒng)”進行分類[16]。
1.3數(shù)據(jù)處理參考宋永昌等[14]計算凋落物中跳蟲群落相似性指數(shù)、多樣性指數(shù)、均勻度、優(yōu)勢度和豐富度等。按照以下公式計算Shannon - Wiener多樣性指數(shù)(H’):
H’=-∑ni/Nln(ni/N)
(1)
Js=H′/lnS
(2)
按照以下公式計算Simpson優(yōu)勢度指數(shù)(C):
C=∑Pi2,Pi=ni/N
(3)
按照以下公式計算Margalef豐富度指數(shù)(D):
D=(S-1)/lnN
河北省是典型的草食畜牧業(yè)和優(yōu)質(zhì)大豆的主產(chǎn)省,2018年,我省青貯玉米、苜蓿等飼草飼料作物種植面積達250萬畝,大豆種植面積超過300萬畝。隨著國家玉米調(diào)減政策的出臺和中美貿(mào)易摩擦的影響,未來大豆的播種面積將進一步擴大,布局進一步優(yōu)化,豐富的飼草、大豆和秸稈為我省畜牧業(yè)發(fā)展提供了飼料來源。我省主要畜產(chǎn)品產(chǎn)量一直位居全國前列,近三年我省畜牧業(yè)平均產(chǎn)值達1902億元,2017年我省畜牧業(yè)總產(chǎn)值達1899.37億元,位居全國第四位,占全省農(nóng)林牧漁總產(chǎn)值超過30%。
(4)
式中,ni為第i個類群的個體數(shù),N為群落中所有類群的個體數(shù),S為類群數(shù)。
對跳蟲與總氮、總磷和有機碳的關(guān)系進行了Pearson線性相關(guān)分析,各樣地總氮、總磷和有機碳用One-way ANOVA進行差異顯著性分析,使用SPSS 19.0統(tǒng)計軟件完成。
2結(jié)果與分析
2.1不同砍伐程度對跳蟲群落的影響
2.1.1不同砍伐程度對跳蟲群落結(jié)構(gòu)的影響。由表1可知,5個樣地4個季節(jié)共獲得跳蟲11 191只,隸屬4目11科。優(yōu)勢類群(個體數(shù)占總數(shù)>10%)為等節(jié)蟲兆科(Isotomidae)、長角蟲兆科(Entomobryidae)和疣蟲兆科(Neanuridae),三者共占個體總數(shù)的81.66%,等節(jié)蟲兆科最多,分別為長角蟲兆科的2.39倍、疣蟲兆科的4.6倍。常見類群(個體數(shù)占總數(shù)的1%~10%)有5科,各科個體數(shù)從多到少依次為棘蟲兆科(Onychiuridae)、土蟲兆科(Tullbergiidae)、球角蟲兆科(Hypogastruridae)、卡天蟲兆科(Katiannidae)和鱗蟲兆科(Tomoceridae),共占總捕獲數(shù)量的17.74%,其余為稀有類群(個體數(shù)占總數(shù)<1%)僅占總個體數(shù)的0.60%。
隨著砍伐程度的不同,跳蟲群落的類群數(shù)和密度也發(fā)生相應(yīng)的變化,5個樣地跳蟲的個體數(shù)從多到少依次為:Ⅱ號樣地(28.79%)、Ⅳ號樣地(21.03%)、Ⅰ號樣地(20.75%)、Ⅴ號樣地(17.09%)、Ⅲ號樣地(12.33%)。類群數(shù)的變化與密度的變化趨勢相同,Ⅱ號樣地和IV號樣地類群數(shù)最多,III號樣地類群數(shù)最少。
此外,不同砍伐程度下各科優(yōu)勢類群和常見類群并不完全相同,等節(jié)蟲兆科(Isotomidae)和長角蟲兆科(Entomobryidae)在5個樣地中始終為優(yōu)勢類群,疣蟲兆科僅在II號樣地和V號樣地為優(yōu)勢類群,其余樣地均為常見類群,棘蟲兆科在III號樣地和IV號樣地為常見類群,在其余3個樣地均為優(yōu)勢類群。土蟲兆科、鱗蟲兆科、卡天蟲兆科和球角蟲兆科在5個樣地均為常見類群。
2.1.2不同砍伐程度對跳蟲群落多樣性的影響。運用Shannon-Wiener多樣性指數(shù)、Pielou均勻度、Simpson優(yōu)勢度指數(shù)和Margalef豐富度指數(shù),對跳蟲群落多樣性的季節(jié)變化進行測度。從圖2可以看出,清除地上植物、保留枯枝落葉的II號樣地雖然密度最高,但其Shannon-Wiener多樣性指數(shù)和Pielou均勻度指數(shù)卻最低;作為對照的V號樣地Shannon-Wiener多樣性指數(shù)和Pielou均勻度指數(shù)最高,密度最低的III號樣地(清除地上全部植物并清除枯枝落葉層和表層土)次之;II號樣地等節(jié)蟲兆科所占比例極大(61.61%),因此其Simpson優(yōu)勢度指數(shù)最高;類群數(shù)最少的III號樣地Margalef豐富度指數(shù)最低,各樣地類群數(shù)從少到多依次為Ⅲ、Ⅰ、Ⅴ、Ⅱ、Ⅳ。
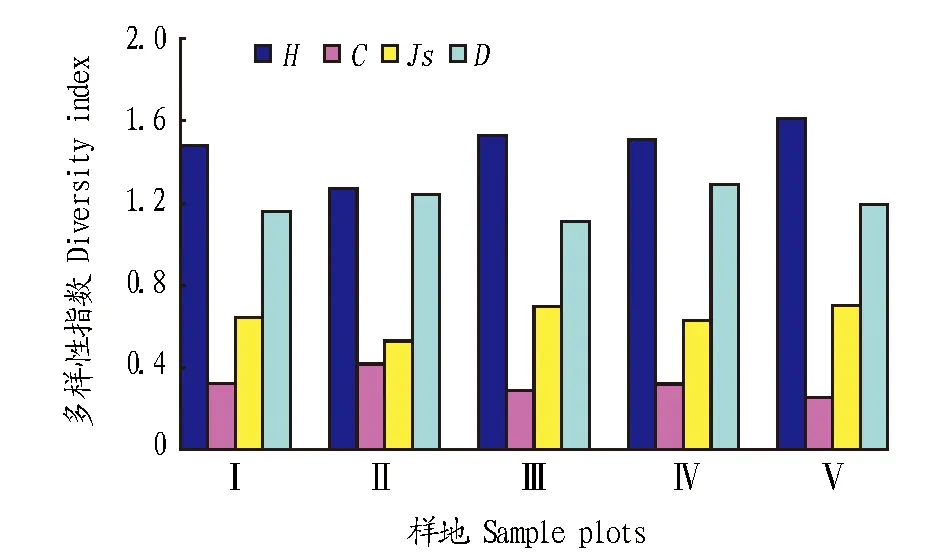
圖2 不同樣地跳蟲群落多樣性比較Fig.2 Comparison of diversity of collembolan communities in different slashed plots
2.2各樣地跳蟲群落的季節(jié)變化從圖3可以看出,跳蟲群落類群組成的季節(jié)變化規(guī)律不明顯,多數(shù)類群在四季均有出現(xiàn),但密度卻發(fā)生明顯變化,跳蟲群落總平均密度為8.80×104ind./m2。秋季大量發(fā)生,占40.45%;夏秋季節(jié)密度相近,冬季密度極少,僅占4季的7.79%。
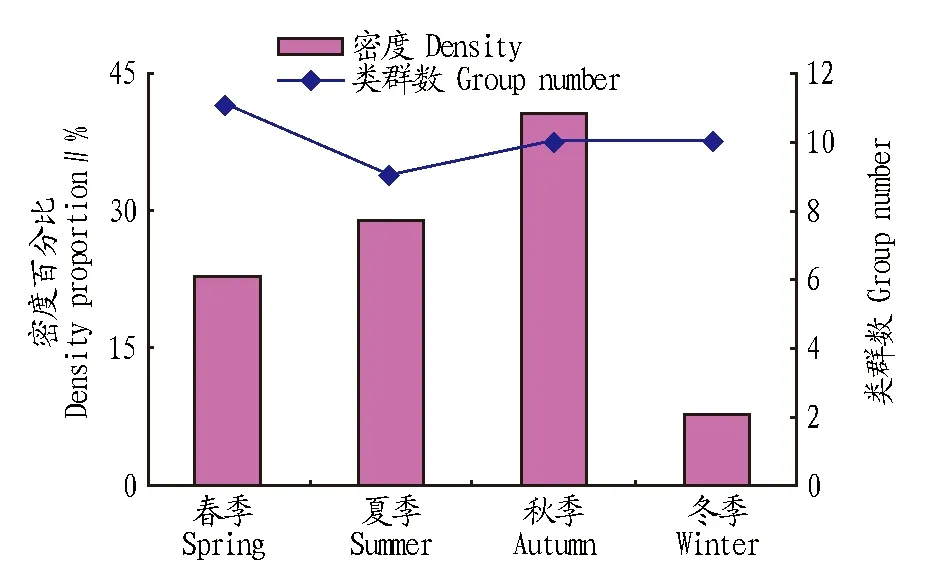
圖3 跳蟲類群數(shù)與密度的季節(jié)變化Fig.3 The seasonal changes of groups number and density of collembols
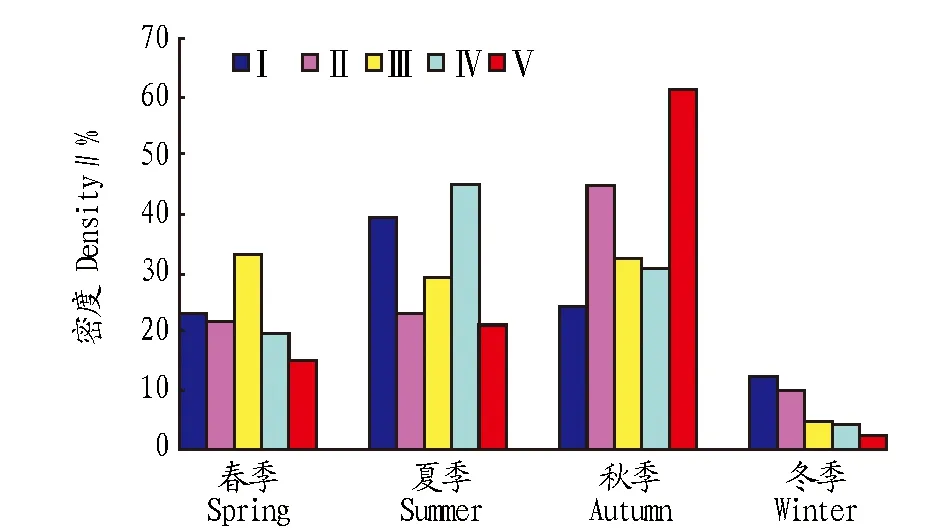
圖4 不同砍伐樣地跳蟲群落數(shù)量的季節(jié)變化Fig.4 Seasonal fluctuation of collembolan density percentage at different treatment plots
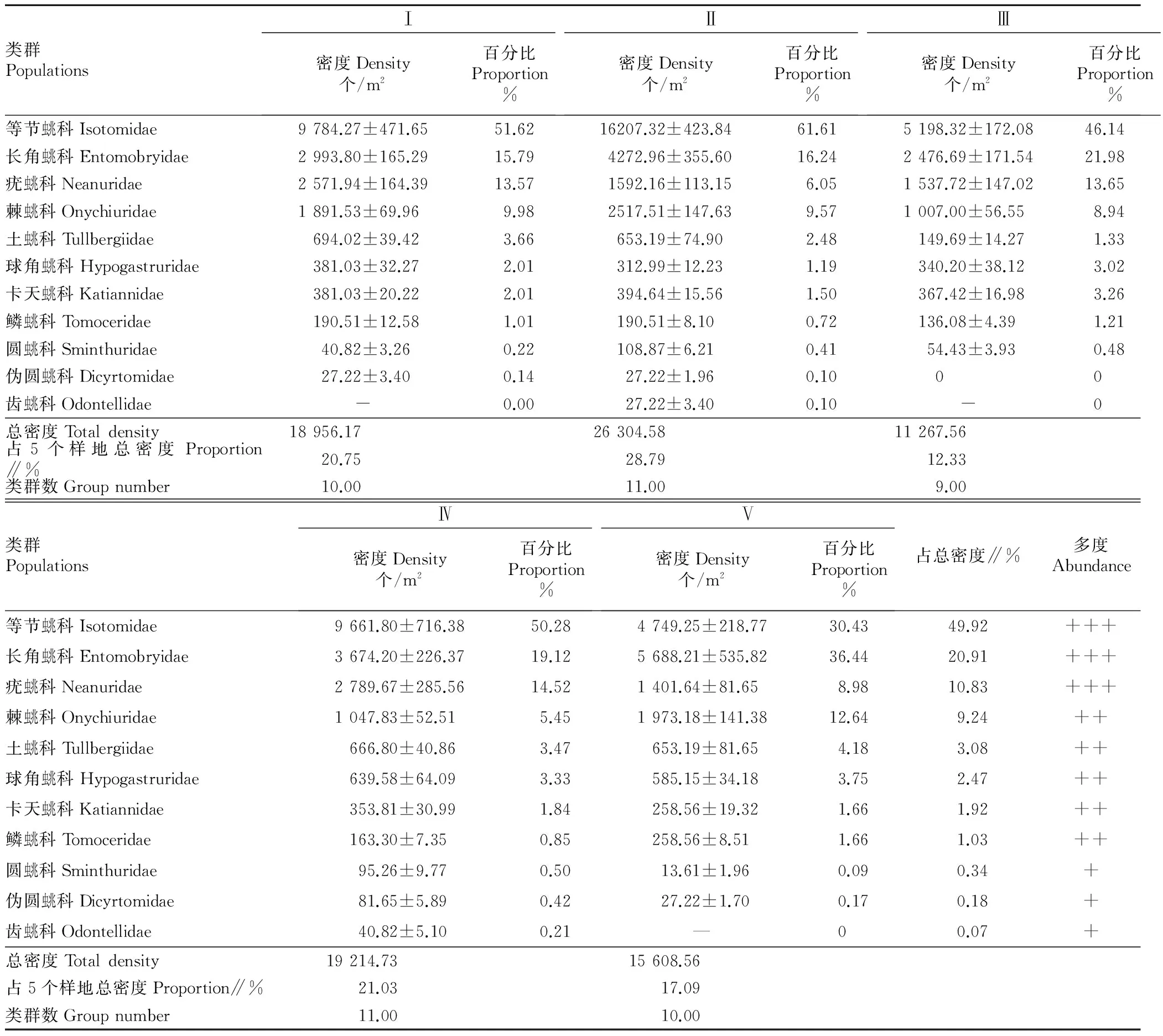
表1 不同砍伐程度跳蟲類群及其密度
由表2可知,不同砍伐程度下跳蟲優(yōu)勢類群也發(fā)生季節(jié)性變化,各樣地夏秋季節(jié)優(yōu)勢類群均為等節(jié)蟲兆科、長角蟲兆科和棘蟲兆科,春冬季節(jié)優(yōu)勢類群主要為等節(jié)蟲兆科和疣蟲兆科,但不同砍伐程度下各樣地又有季節(jié)性變化。等節(jié)蟲兆科在各個樣地4個季節(jié)中始終為優(yōu)勢類群,疣蟲兆科僅在春冬季節(jié)優(yōu)勢明顯,棘蟲兆科和長角蟲兆科僅在夏秋季節(jié)優(yōu)勢明顯,球角蟲兆科只在IV、V號樣地的春季為優(yōu)勢類群,卡天蟲兆科只在V號樣地的冬季為優(yōu)勢類群。
主要類群的季節(jié)消長導(dǎo)致了跳蟲群落組成百分比的季節(jié)性變化。從圖5可以看出,不同類群的數(shù)量季節(jié)消長不同。等節(jié)蟲兆科(Isotomidae)始終是四季的優(yōu)勢類群,等節(jié)蟲兆科的最高峰在夏季,占該類群全年總數(shù)的35.66%,冬季密度最低,僅占19.7%;而長角蟲兆科(Entomobryidae)在秋季大量發(fā)生,占該類群全年總數(shù)的52.44%,春冬季節(jié)密度最低,分別占7.42%和7.53%;疣蟲兆科(Neanuridae)和球角蟲兆科(Hypogastruridae)均是在春季密度最高,分別占該類群全年總數(shù)的53.89%和66.55%,其次為冬季,夏秋季節(jié)密度相近且占全年密度最低。卡天蟲兆科(Katiannidae)和鱗蟲兆科(Tomoceridae)棘蟲兆科(Onychiuridae)和土蟲兆科(Tullbergiidae)。
2.3凋落物養(yǎng)分含量的變化及其與跳蟲群落之間的關(guān)系由表3可知,各樣地有機碳、總氮和總磷均存在顯著差異,其中有機碳和總氮含量從高到低均依次為:Ⅲ號樣地、Ⅱ號樣地、Ⅰ號樣地、Ⅴ號樣地、Ⅳ號樣地;總磷含量在Ⅰ號樣地最高(0.27),Ⅲ號樣地最低(0.11),其余3個樣地較接近。
由表4可知,多數(shù)類群跳蟲與凋落物養(yǎng)分含量之間無顯著相關(guān)性,只有土蟲兆科密度與總氮含量呈顯著正相關(guān)(r=-0.907,P< 0.05),與總磷含量呈顯著負相關(guān)(r=-0.99,P<0.01),球角蟲兆科密度與有機碳含量呈顯著正相關(guān)(r=0.969,P<0.01)。
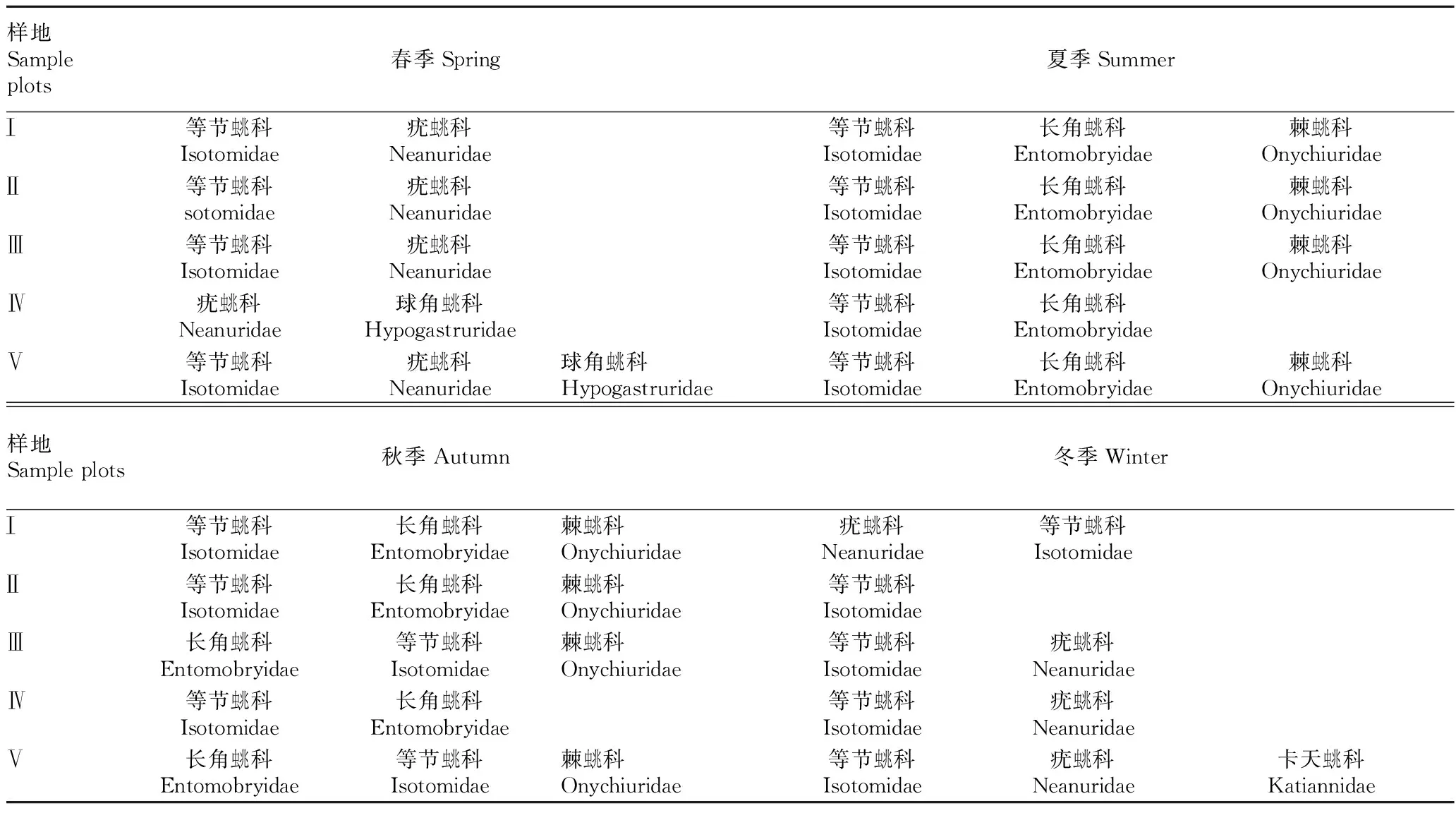
表2 各樣地優(yōu)勢類群的季節(jié)變化
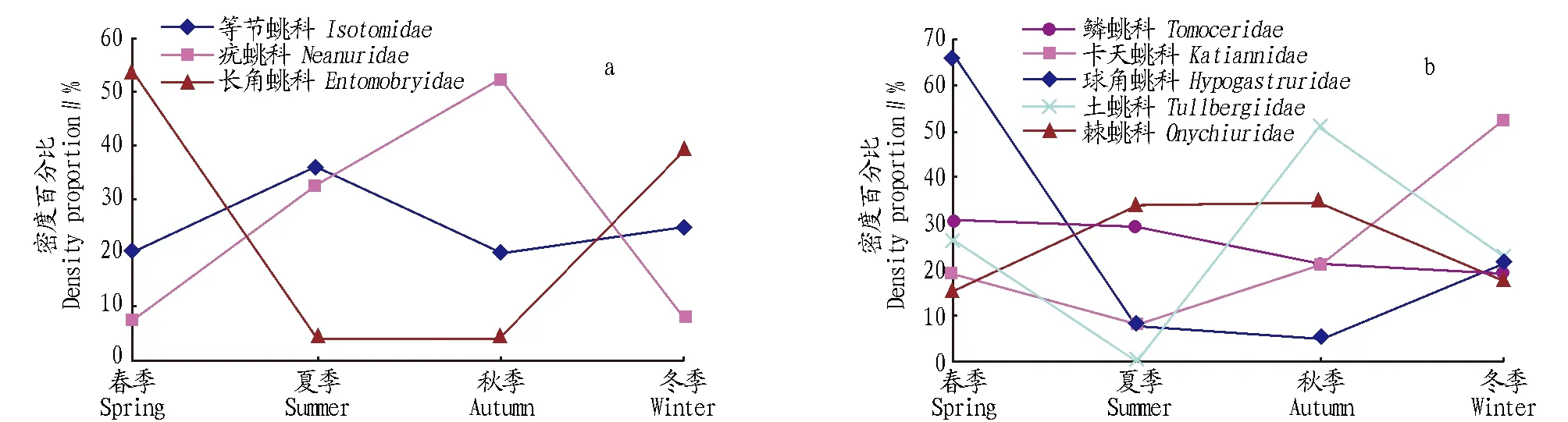
圖5 主要類群的季節(jié)變化Fig.5 Seasonal changes of main collembolan groups
3討論與結(jié)論
森林中跳蟲群落的組成和分布,一方面與森林立地和土壤條件有關(guān),另一方面受林內(nèi)微環(huán)境的影響。森林的干擾不僅表現(xiàn)在植物群落多樣性的變化,而且表現(xiàn)在森林生態(tài)環(huán)境的變化[17]。森林被砍伐后,原來復(fù)雜的植物群落結(jié)構(gòu)發(fā)生改變,地表裸露程度增大,導(dǎo)致林內(nèi)溫濕度和光照等氣候變化[18],凋落物層微環(huán)境發(fā)生相應(yīng)變化,這些變化對跳蟲的棲息環(huán)境和食物源產(chǎn)生很大的影響,它直接作用于跳蟲群落,從而導(dǎo)致凋落物層跳蟲群落結(jié)構(gòu)發(fā)生很大改變[19]。不同砍伐強度使各樣地所受干擾程度不同,引起樣地中植被恢復(fù)不同。該研究結(jié)果表明,雖然經(jīng)過6年的恢復(fù)后,跳蟲類群組成雖然相似,但優(yōu)勢類群、群落密度及多樣性也有所差異。由此可見,跳蟲群落對不同程度的林地砍伐處理較為敏感。
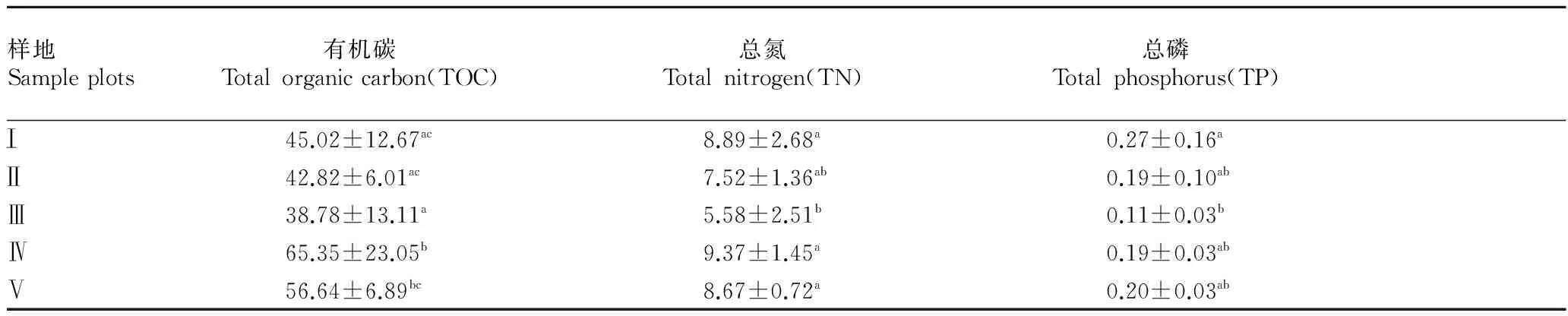
表3 不同砍伐程度下凋落物層養(yǎng)分含量的比較
注: 同列不同小寫字母表示差異顯著(P<0.05).
Note: Different lowercases in the same column stand for significant difference(P<0.05).
表4主要類群密度與養(yǎng)分含量的相關(guān)系數(shù)
Table 4Correlation coefficients of densities of main groups with nutrients contents
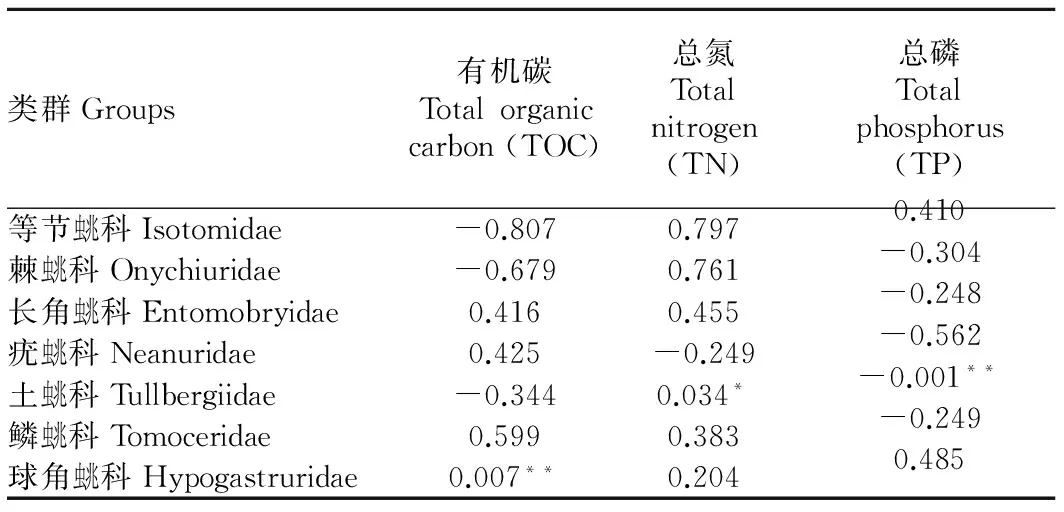
類群Groups有機碳Totalorganiccarbon(TOC)總氮Totalnitrogen(TN)總磷Totalphosphorus(TP)等節(jié)蟲兆科Isotomidae-0.8070.7970.410棘蟲兆科Onychiuridae-0.6790.761-0.304長角蟲兆科Entomobryidae0.4160.455-0.248疣蟲兆科Neanuridae0.425-0.249-0.562土蟲兆科Tullbergiidae-0.3440.034*-0.001**鱗蟲兆科Tomoceridae0.5990.383-0.249球角蟲兆科Hypogastruridae0.007**0.2040.485
注:*表示相關(guān)性達到顯著水平(P<0.05);**表示相關(guān)性達到極顯著水平(P<0.01)。
Note: * stands for significant correlation(P<0.05); ** stands for extremely significant correlation (P<0.01).
跳蟲群落密度和豐富度最高值并非出現(xiàn)在未砍伐的Ⅴ號樣地,而是砍伐程度較輕的Ⅰ、Ⅱ、Ⅳ號樣地較高,皆伐的Ⅲ號樣地密度和豐富度最低,這一現(xiàn)象出現(xiàn)的原因可能包括:適度的干擾會促進植物群落新舊更替,擁有較高的生物多樣性,而伴隨著樣地不斷的恢復(fù)更新,豐富的凋落物也為跳蟲的生存和發(fā)展提供了充分的條件[10]。Ⅲ號樣地受干擾程度較大,不僅清除地上植物部分,且去除最適宜跳蟲生存的10 cm表層土,這在很大程度上改變了跳蟲生存的微環(huán)境,即使經(jīng)過6年的恢復(fù),跳蟲群落密度和豐富度仍較其他樣地低。該研究結(jié)果表明,隨著砍伐程度的加劇,跳蟲群落類群組成減少,群落密度和豐富度也隨之降低。由此可見,跳蟲群落的結(jié)構(gòu)變化可以在一定程度上反映森林受干擾程度,對于進一步揭示反映森林演替和干擾的土壤動物指示類群具有重要意義。
跳蟲群落的季節(jié)變化主要表現(xiàn)在各樣地不同季節(jié)所占的百分比差異和不同樣地間優(yōu)勢類群的季節(jié)性差異。該研究結(jié)果表明,各樣地四季中夏秋季節(jié)跳蟲密度占比較高,冬春較低,這一結(jié)果與國內(nèi)同類研究結(jié)果相似[20-21]。跳蟲對溫濕度變化比較敏感[22],夏秋季豐富的凋落物和適宜的溫濕度可能是跳蟲密度偏高的主要原因。優(yōu)勢類群和常見類群的季節(jié)變化體現(xiàn)出不同類群跳蟲對環(huán)境的適應(yīng)性差異[23],如四季溫度變化、季節(jié)性降雨差異[24]等,此外,不同季節(jié)凋落物處在不同的分解階段為跳蟲提供了不同的食物源[5,25],直接影響了跳蟲群落結(jié)構(gòu)的季節(jié)性變化。
該研究結(jié)果表明隨著砍伐程度的加劇,凋落物有機碳和總氮含量呈減少趨勢,Ⅰ、Ⅱ、Ⅲ號樣地的地表植物被完全清除或部分清除,地表凋落物含量較少,所以有機碳含量較低,尤其是Ⅲ號樣地凋落物含量極少,因此有機碳、總氮和總磷含量均最少。只清除下木層的Ⅳ號樣地砍伐程度較弱,經(jīng)過6年恢復(fù)后其凋落物儲量已超過原有水平,因此有機碳和總氮含量最高。總磷含量雖然在不同砍伐程度下有顯著差異,但沒有明顯的變化規(guī)律。
跳蟲群落主要類群密度與養(yǎng)分含量的相關(guān)性分析表明,不同種類的跳蟲其偏好的養(yǎng)分情況并不一致。由此推測,不同的跳蟲類群偏好的主要食物可能存在差異,這也可能是眾多跳蟲類群能夠在同一環(huán)境中共存的主要機理[26-27]。養(yǎng)分狀況只能反映凋落物在樣地間的含量差異,關(guān)于凋落物對具體某一類群跳蟲的適口性和偏好性并不能在養(yǎng)分狀況上得以明確體現(xiàn)[28],這也是一部分跳蟲類群與養(yǎng)分含量的相關(guān)關(guān)系并不顯著的主要原因。
參考文獻
[1] FILSER J.The role of Collembola in carbon and nitrogen cycling in soil[J].Pedobiologia,2002, 46: 234-245.
[2] 陳建秀,麻智春,嚴(yán)海娟,等.跳蟲在土壤生態(tài)系統(tǒng)中的作用[J].生物多樣性,2007(2): 154-161.
[3] RUSEK J.Biodiversity of Collembola and their functional role in the ecosystem[J].Biodiversity and conservation,1998, 7: 1207-1219.
[4] SINGH J S,GUPTA S R.Plant decomposition and soil respiration in terrestrial ecosystems[J].The botanical review,1977, 43(4): 449-528.
[5] 柯欣,趙立軍,尹文英.青岡林土壤跳蟲群落結(jié)構(gòu)在落葉分解過程中的變化[J].生態(tài)學(xué)報,2001(6): 982-987.
[6] 柯欣,趙立軍,尹文英.青岡林土壤動物群落結(jié)構(gòu)在落葉分解過程中的演替變化[J].動物學(xué)研究,1999(3): 48-54.
[7] MALMSTR?M A,PERSSON T,AHLSTR?M K.Effects of fire intensity on survival and recovery of soil microarthropods after a clearcut burning[J].Canadian journal of forest research,2008, 38(9): 2465-2475.
[8]CUCHTAP,MIKLISOVD,KOVCL’.Athree-yearstudyofsoilCollembolacommunitiesinspruceforeststandsoftheHighTatraMts(Slovakia)afteracatastrophicwindthrowevent[J].Europeanjournalofsoilbiology,2012, 50: 151-158.
[9] 陳利頂,傅伯杰.干擾的類型、特征及其生態(tài)學(xué)意義[J].生態(tài)學(xué)報,2000(4): 581-586.
[10] 陳小鳥,由文輝,王向陽,等.常綠闊葉林不同砍伐處理下土壤動物的群落特征[J].生物多樣性.2009, 17(2): 160-167.
[11] 易蘭,由文輝,宋永昌.天童常綠闊葉林與演替階段凋落物中的土壤動物群落[J].生態(tài)學(xué)報,2005(3):466-473.
[12] 易蘭.浙江天童受損常綠闊葉林的次生演替對土壤動物群落的影響[D].上海:華東師范大學(xué),2005.
[13] 陳小鳥.天童常綠闊葉林不同砍伐程度對土壤動物群落的影響[D].華東師范大學(xué), 2009.
[14] 宋永昌,陳小勇.中國東部常綠闊葉林生態(tài)系統(tǒng)退化機制與生態(tài)恢復(fù)[M].北京: 科學(xué)出版社, 2007.
[15] 忻介六.土壤動物知識[M].北京: 科學(xué)出版社, 1986.
[16]BELLINGERP,CHRISTIANSENK,JANSSENSF.ChecklistoftheCollembolaoftheworld[Z].2009.
[17]BENGTSSONJ,NILSSONSG,F(xiàn)RANCA,etal.Biodiversity,disturbances,ecosystemfunctionandmanagementofEuropeanforests[J].ForestEcologyandManagement.2000, 132: 39-50.
[18] 胡小飛,陳伏生,葛剛.森林采伐對林地表層土壤主要特征及其生態(tài)過程的影響[J].土壤通報,2007, 38(6): 1213-1218.
[19] 柯欣,趙立軍,尹文英.青岡林土壤跳蟲群落結(jié)構(gòu)在落葉分解過程中的變化[J].生態(tài)學(xué)報,2001(6): 982-987.
[20] 吾瑪爾·阿布力孜,阿布都如蘇力·土遜,木卡熱木·阿吉木,等.烏魯木齊地區(qū)不同生境下土壤跳蟲群落結(jié)構(gòu)及多樣性研究[J].四川動物.2013, 32(1): 60-67.
[21] 靳亞麗,由文輝,易蘭,等.天童森林生態(tài)系統(tǒng)凋落物層跳蟲群落的生態(tài)學(xué)研究[J].生態(tài)環(huán)境學(xué)報,2011(2): 241-247.
[22] 劉晶,王云彪,吳東輝.土壤跳蟲適應(yīng)低溫環(huán)境的策略與機制研究進展[J].應(yīng)用生態(tài)學(xué)報,2012(12): 3526-3532.
[23]NAKAMORIT,SUZUKIA.Spore-breakingcapabilitiesofcollembolansandtheirfeedinghabitatwithinsporocarps[J].Pedobiologia.2005, 49(3): 261-267.
[24]TURNBULLMS,LINDOZ.CombinedeffectsofabioticfactorsonCollembolacommunitiesrevealprecipitationmayactasadisturbance[J].Soilbiologyandbiochemistry,2015, 82: 36-43.
[25] 董煒華, 殷秀琴.小興安嶺白樺林土壤跳蟲的動態(tài)特征[J].生態(tài)學(xué)雜志, 2005(3):278-282.
[26]ITOHR.GrowthandlifecycleofanArborealCollembola,XenyllabrevispinaKinoshita,withspecialreferencetoitsseasonalmigrationbetweentreeandforestfloor[J].Edaphologia,1991,45:33-48.
[27]NAKAMORIT,SUZUKIA.Spore-breakingcapabilitiesofcollembolansandtheirfeedinghabitatwithinsporocarps[J].Pedobiologia, 2005, 49(3):261-267.
[28] 易蘭, 由文輝.天童植被演替過程中環(huán)境因子對土壤動物群落的影響[J].華東師范大學(xué)學(xué)報(自然科學(xué)版), 2006(6):109-116.
基金項目高等學(xué)校博士學(xué)科點專項基金科研項目(20070269036);上海科技館種子資金項目。
作者簡介靳亞麗(1985-),女,河北武安人,研究實習(xí)員,碩士,從事土壤動物生態(tài)學(xué)研究。
收稿日期2016-04-23
中圖分類號Q 958.1
文獻標(biāo)識碼A
文章編號0517-6611(2016)16-023-05
Ecological Distribution ofCollembolain the Litter under Different Ground Cover Treatment of Evergreen Broad-leaved Forest
JIN Ya-li1, JIANG Yue2
(1.Natural History Research Center of Shanghai Natural History Museum, Shanghai 200127; 2.School of Ecological and Environment Sciences, East China Normal University, Shanghai 200062)
Abstract[Objective] To understand the effects of ground cover removal on collembola communities in the litter and the community restoration situations.[Method] The collembola community investigation of 4 seasons in an evergreen broad-leaved forest after 7 years since the ground cover removal in Tiantong Region, Zhejiang Province was carried out during Dec.2019-Sep.2010.The five treatments in five sites were as follows: (Ⅰ)big trees were removed but sprouts retained; (Ⅱ) ground surface vegetation was removed but litter retained; (Ⅲ) all vegetation and 0-10 cm topsoil were removed; (Ⅳ) undergrowth was removed but big trees retained; and (Ⅴ) a control plot without any disturbance.[Result] The number of collembolan groups did not vary widely.The number of collembola community was as follows: Ⅱ (28.79%) > Ⅳ(21.03%) > Ⅰ (20.75%) > Ⅴ (17.09%) >Ⅲ (12.33%) .The community composition vary in different seasons.The number of collembola community was as follows: autumn > summer > spring > winter.The organic carbon, total nitrogen and total phosphorus had obvious differences among the plots.Even after 7 years of restoration, the collembola community traits were obviously different among the treatments.[Conclusion] Collembola community was very sensitive to the environmental disturbing, which can be used as the indicator species to evaluate the soil environmental disturbance.
Key wordsDisturbance; Collembola; Litter layer; Evergreen broad-leaved forest; Tiantong
