冬小麥葉片顯微結構和光合特性與產量的關系
2016-07-29 07:47:35黨紅凱李瑞奇李雁鳴李曉爽
麥類作物學報 2016年6期
黨紅凱,李瑞奇,李雁鳴,李曉爽,孟 建
(1.河北農業大學農學院/河北省作物生長調控重點實驗室,河北保定 071000;2.河北省農林科學院旱作農業研究所,河北衡水 053000)
?
冬小麥葉片顯微結構和光合特性與產量的關系
黨紅凱1,2,李瑞奇1,李雁鳴1,李曉爽1,孟 建1
(1.河北農業大學農學院/河北省作物生長調控重點實驗室,河北保定 071000;2.河北省農林科學院旱作農業研究所,河北衡水 053000)
摘要:為了解冬小麥葉片顯微結構和光合特性與產量的關系,采用大田跟蹤對比調查方法,對8個高產冬小麥品種的葉片顯微結構、光合特征及產量性狀進行分析。結果表明,小麥不同葉位葉片主要由規則環狀細胞組成,各葉位葉片規則細胞的平均環數為1.7~4.5環。隨葉位升高,葉肉細胞平均環數及上表皮和下表皮氣孔密度均有增加的趨勢。各品種旗葉葉綠素含量最大峰值出現在5月10日前后;旗葉可溶性蛋白含量變化與葉綠素含量變化相似,最大值陸續出現在5月7-20日。各品種葉片葉肉細胞平均環數和平均光合速率均以旗葉最高。但品種間光合勢、葉源量的差異與葉肉細胞和葉綠素含量并不完全一致。品種間產量性狀的差異主要表現在生物產量與經濟系數上。生物產量與倒2葉光合勢呈顯著正相關,與倒3葉光合勢呈極顯著正相關。綜上可見,光合性能好且光合產物能夠有效轉運進入籽粒,是小麥品種獲取高產的必要特征。
關鍵詞:冬小麥;葉片;解剖特征;光合特性;產量性狀
葉片是小麥重要的光合器官。葉片顯微結構是行使其光合功能的形態學和解剖學基礎[1]。小麥葉片顯微結構受遺傳因素影響,但可塑性比較強[2]。不同葉位葉片生長過程中,解剖特征與光合特性對環境變化的響應不同[3]。系統比較不同品種小麥葉片顯微結構、光合特性和產量形成,對認識葉片結構與光合性能的內在聯系,提高小麥綜合生產能力具有重要意義。關于冬小麥葉片解剖特征與光合功能的關系,前人已經進行了系統研究。劉 瑩等[2]和李雁鳴等[4]利用華北地區不同年代的小麥品種,研究了葉肉細胞形態和光合性能演替規律。劉永康等[5]通過研究小麥旗葉的解剖特征、群體與個體光合特性,明確了不同株型小麥的高光效機理。李金霞等[6]通過噴施不同濃度不同種類的植物生長調節劑進行研究,結果發現,由于調節劑的影響,在不增加葉面積的條件下,減小旗葉3環葉肉細胞長與寬,增加旗葉厚度,可以改善葉片質量。小麥是河北省的主要糧食作物,但對河北省主推冬小麥品種葉片顯微結構、光合功能及與產量形成的關系尚缺乏系統研究。本研究以河北省8個高產優質冬小麥品種為材料,通過觀察和分析葉片顯微結構、光合特性及產量構成,探討了其內在的聯系,以期為進一步挖掘與冬小麥產量密切相關的光合生產潛力提供參考依據。
1材料與方法
1.1試驗基本情況
田間試驗于2014-2015年在河北省藁城市農業科學研究所進行。供試小麥品種為石新733、石家莊8號、石4185、河農822、藁8901、金麥 54、師欒02-1和藁優2018,其中前4個品種為高產品種,分別用SX733、SJZ8、S4185、HN822表示;后4個品種為優質強筋小麥品種,分別用G8901、JM54、SL02-1、GY2018表示。供試土壤有機質含量18.41 g·kg-1,全氮含量0.94 g·kg-1,堿解氮含量107.2 mg·kg-1,速效磷含量22.4 mg·kg-1,速效鉀126.7 mg·kg-1,速效鋅含量3.6 mg·kg-1,速效錳含量18.3 mg·kg-1,速效銅含量1.3 mg·kg-1。前茬夏玉米收獲后按常規整地,2014年10月11日播種,行距15 cm,采用各品種審定時推薦的適宜種植密度,SX733、SJZ8、S4185、HN822、G8901、JM54、SL02-1、GY2018的基本苗分別為每公頃322.5萬、285.1萬、280.4萬、300.2萬、273.1萬、264.0萬、260.0萬和285.1萬株。每個品種設3次重復,小區面積30 m2。春季生長階段(3月1日-6月11日)>0℃積溫1 666.1 ℃,日照時數844.2 h。田間管理采用常規高產栽培技術。
1.2觀測內容與方法
1.2.1頂三葉光合速率測定
從旗葉展開開始測定頂三葉的光合速率。在晴天上午9:00-11:00,每個品種選取3~5株受光方向和生長狀況一致的葉片,用美國產CI-340光合作用測定系統進行閉路測定。每5~7 d測定一次。
1.2.2葉片解剖學特征觀察
于春季每個葉位新生葉片定型時,各品種分別選擇有代表性的健壯植株,摘取3~5片新定型葉位的葉片,室內洗凈擦干,進行以下性狀的觀察。
(1)葉肉細胞形態觀察:取葉片的中部,用段續川[7]細胞離析法固定、分離、染色后制片。每個葉位制作3張制片,作為3次重復。每個制片在日本產奧林巴斯CHS顯微鏡下觀察400~500個細胞,統計每個細胞的形態和環數。
(2)規則葉肉細胞大小測定:在顯微鏡下,每個制片中用測微尺連續測量25個2環細胞的高度和寬度。
(3) 葉片厚度測定:去掉中脈,葉片中部徒手縱切。從邊緣到中脈,用測微尺等距離測量5點厚度,重復10次。計算平均數,作為葉片厚度。
(4)氣孔密度測定:同上取樣,每次取10片葉片,用帶尖的小鑷子分別撕取葉片中部靠近中脈部位的上表皮和下表皮,置顯微鏡下觀察,數5個視野的氣孔數,用測微尺測算視野面積,計算氣孔密度。
1.2.3旗葉葉綠素含量與可溶性蛋白含量測定
于旗葉定型后,分7個時期取各品種旗葉,用去離子水洗凈其表面灰塵,采用趙世杰等[8]的方法測定葉綠素含量。分5個時期取各品種旗葉,用去離子水洗凈其表面灰塵,采用Bradford的方法[9]測定可溶性蛋白含量。
1.2.4產量及其構成因素測定
成熟期按常規方法測定每公頃生物產量、產量構成因素(每公頃穗數、穗粒數、千粒重)和籽粒產量。
2結果與分析
2.1不同冬小麥品種葉片解剖特征的比較
2.1.1不同冬小麥品種各葉位葉片葉肉細胞的形態差異
由表1可見,小麥不同葉位春生葉片主要由規則環狀細胞組成,占總細胞數的93.1%~99.9%,而不規則細胞(分支細胞)所占的比例較小。在規則細胞中,1~5環細胞的比例較大,達到60%以上,8環以上細胞所占比例較小。各葉位葉片規則細胞的平均環數1.7~4.7環,隨葉位升高,葉片規則細胞平均環數有增高的趨勢。
各品種不同葉位葉片細胞組成不同。倒6葉主要由1環和2環細胞組成,多數品種以1環細胞所占比例最高。不同品種比較,倒6葉1環細胞比例以SL02-1最高,為51.3%;2環細胞比例以SX733最高,為43.8%。倒5葉主要由1~3環細胞組成,以2環細胞所占比例最高。不同品種比較,倒5葉1環細胞比例以S4185最高,為31.2%;2環細胞比例以G8901最高,為48.4%;3環細胞比例以HN822最高,為24.1%。與倒5葉相似,倒4葉也主要由1環、2環和3環細胞組成,多數品種以2環細胞所占比例最高。倒3葉主要由1~5環細胞組成,以2環或3環細胞比例最高或較高。其中2環細胞比例以G8901最高,為40.3%;3環細胞比例以HN822最高,達30.6%。倒2葉主要由2~5環細胞組成,以2環或3環細胞比例最高。不同品種比較,倒2葉2環細胞比例以G8901最高,為39.4%;3環細胞比例以GY2018最高。旗葉主要由2~6環細胞組成,除HN822以4環細胞比例最高外,其他品種均以2環或3環細胞比例最高。不同品種比較,旗葉2環細胞比例以JM54最高,達26.5%;3環細胞比例以SX733最高,達23.7%;4環細胞比例以HN822最高,達25.6%;5環細胞比例以G8901最高,達18.9%。
同一品種不同葉位葉片比較,倒5葉的葉肉細胞平均環數不同程度高于倒6葉,但均低于頂三葉。頂三葉隨葉位變化在品種間表現不同,SX733、SJZ8、S4185、G8901等4個品種隨葉位的升高,平均細胞環數逐漸增加,均以旗葉最高;而SL02-1、GY2018、JM54等3個品種,雖然以旗葉葉肉細胞平均環數最高,但倒3葉次之,倒2葉最低;只有HN822以倒3葉最高,旗葉次之,倒2葉最低。
2.1.2不同冬小麥品種頂三葉面積和2環葉肉細胞的大小
不同品種間同葉位比較,頂三葉面積均以GY2018最大,SL02-1最小。同一品種頂三葉之間比較,除HN822、SX733以倒2葉面積最大外,其他6個品種均以旗葉最大,所有品種都以倒3葉面積最小(表2)。
不同品種冬小麥頂三葉葉肉細胞寬度變化范圍為25.0~50.2 μm,高度為28.8~62.8 μm。不同品種間同葉位葉片葉肉細胞寬度和高度的差異較大。其中,S4185倒3葉的細胞寬度和寬高積最大,細胞高度則與2個最大的品種間差異不顯著;G8901倒2葉的細胞高度、寬度及其乘積都最大,但均與GY2018的差異不顯著; GY2018旗葉的細胞寬度、高度及其乘積最大。同一品種不同葉位葉片比較,S4185、SJZ8、SX733等3個品種葉片葉肉細胞寬高積隨葉位升高而減小,以旗葉最小;SL02-1、G8901、GY2018等3個品種也以旗葉最小,但以倒2葉最大;HN822、JM54以倒2葉最小,倒3葉最大。
2.1.3不同冬小麥品種的葉片厚度
各品種春生葉片厚度變化范圍為154.0~293.3 μm(表3)。不同品種間同葉位比較,葉片厚度沒有規律性差異。頂三葉以SX733最厚或較厚,以JM54最薄或較薄;倒4葉、倒5葉以HN822較厚,以JM54最薄或較薄;倒6葉以S4185、SJZ8較厚,GY2018和SX733較薄。同一品種不同葉位間比較,隨葉位變化,葉片厚度的變化規律不同,G8901、SL02-1、JM54、S4185和SJZ8等5個品種以倒6葉最厚,基本上隨葉位升高逐漸變薄,旗葉最薄,但SL02-1、JM54、S4185表現為倒2葉厚于倒3葉。GY2018以倒5葉最厚,倒3葉最薄,倒3葉以上隨葉位升高逐漸加厚。HN822先隨葉位升高逐漸加厚,以倒4葉最厚,倒4葉以上又逐漸變薄。
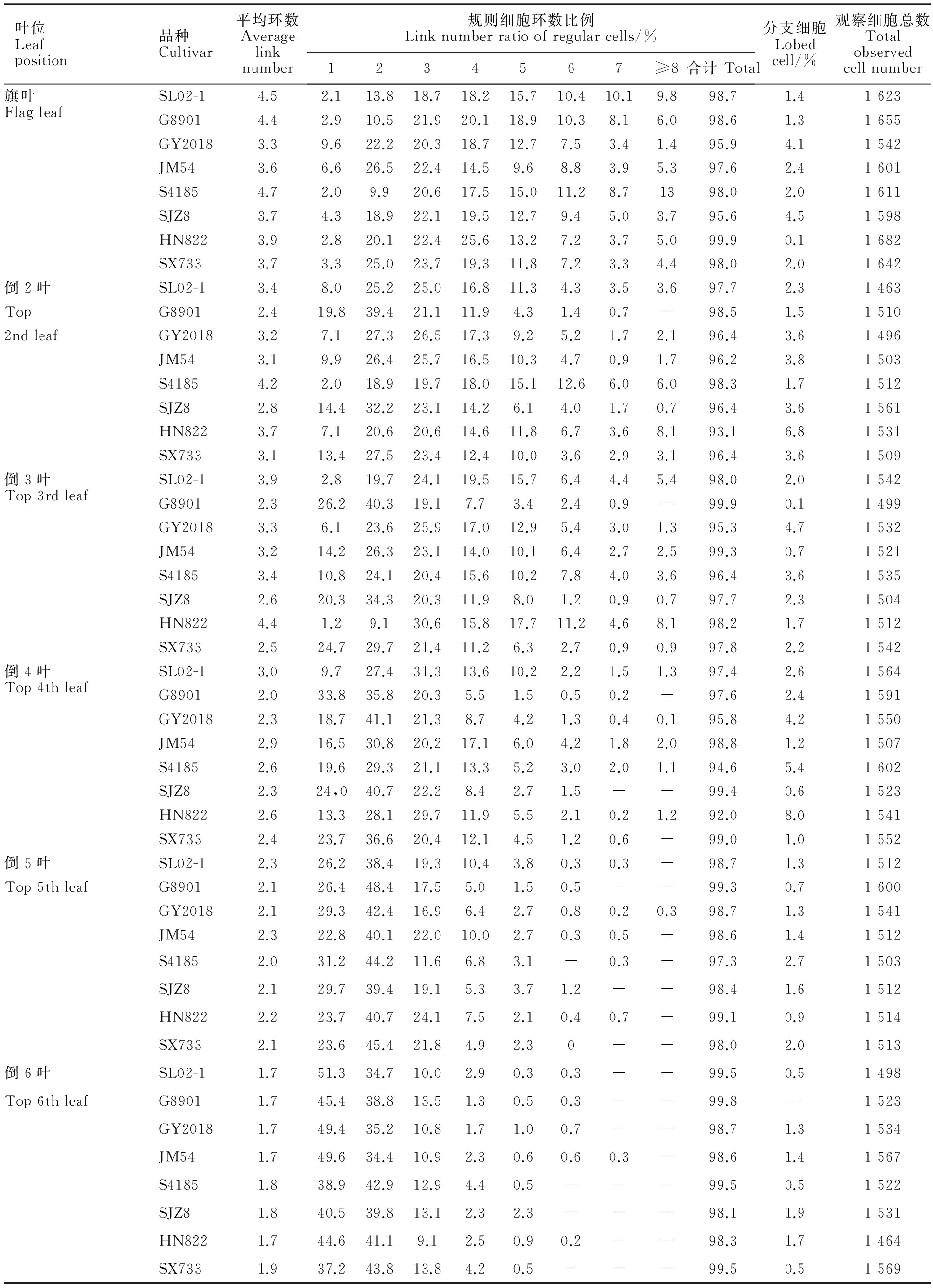
表1 冬小麥春生葉片中不同形態葉肉細胞的分布
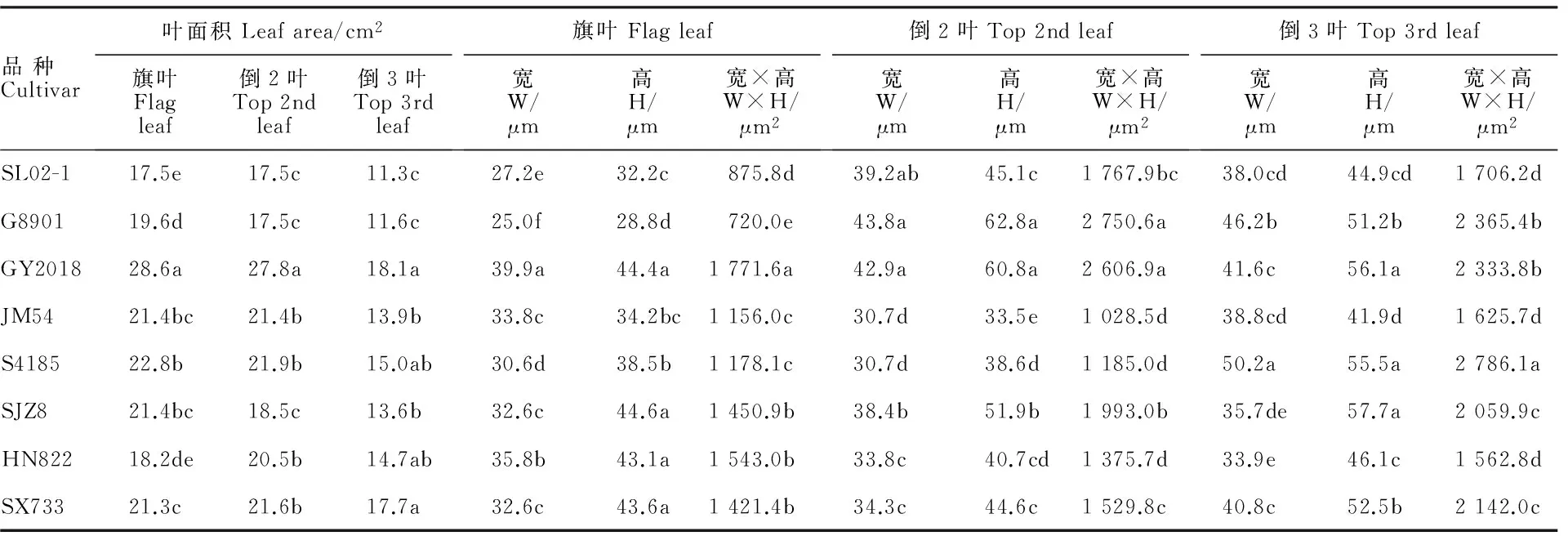
表2 不同冬小麥品種頂三葉面積與2環葉肉細胞的大小
數字后面的小寫字母不同表示不同品種間差異顯著(P<0.05)。下同
W:Width;H:Height.The different small letters following the data show the significant different among different cultivars at 0.05 level.The same as bellow
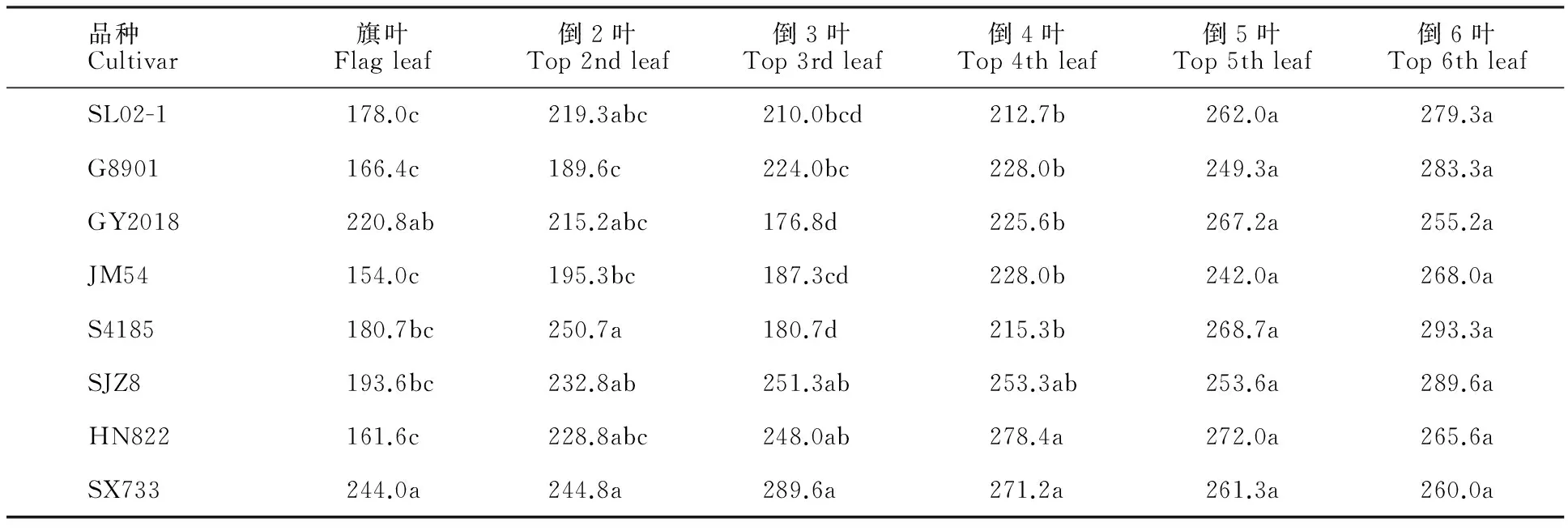
表3 不同冬小麥品種主莖春生葉片的厚度 μm
2.1.4不同冬小麥品種各葉位葉片的氣孔密度
春生葉片上表皮氣孔密度變化范圍為52.3~233.0個·mm-2,下表皮的氣孔密度變化范圍為47.0~203.4個·mm-2(表4)。不同品種間同葉位比較,S4185的葉片氣孔密度較大,其次為SX733,二者顯著高于其他品種。同一品種不同葉位間比較,隨葉位升高,葉片上、下表皮氣孔密度有逐漸增加的趨勢,基本以旗葉最高。上下表皮間比較,同一品種同葉位葉片上表皮的氣孔密度均高于下表皮。
2.2冬小麥旗葉葉綠素和可溶性蛋白質含量的動態變化
冬小麥品種旗葉葉綠素含量最大值為3.49~4.94 mg·g-1,陸續出現在挑旗到開花期(5月2日-5月11日)(圖1)。旗葉葉綠素含量最大值出現后,大部分品種呈逐漸下降趨勢。但SL02-1、GY2018表現有所不同,在葉綠素含量最大峰值出現后,灌漿中期又有所升高。品種間比較,SL02-1葉綠素含量最大值最小,但高峰持續期最長。灌漿末期(6月2日)開始,各品種葉片葉綠素含量隨葉片衰亡而迅速下降。
各品種旗葉可溶性蛋白與葉綠素含量的變化趨勢相似。旗葉可溶性蛋白含量最大值為32.96~52.32 mg·g-1。品種間可溶性蛋白含量最大值比較,以GY2018最大,SL02-1最小。可溶性蛋白含量最大值以G8901、SJZ8、HN822和SX733等品種出現較早。品種間可溶性蛋白含量的變化趨勢,HN822、SX733為雙峰曲線,HN822峰值分別出現在5月7日和5月23日,SX733的峰值則分別出現在5月7日與5月30日;其他品種為單峰曲線,峰值陸續出現在5月7日或5月12日。灌漿末期(5月30日)開始,各品種葉片可溶性蛋白含量隨葉片的衰亡而逐漸下降。
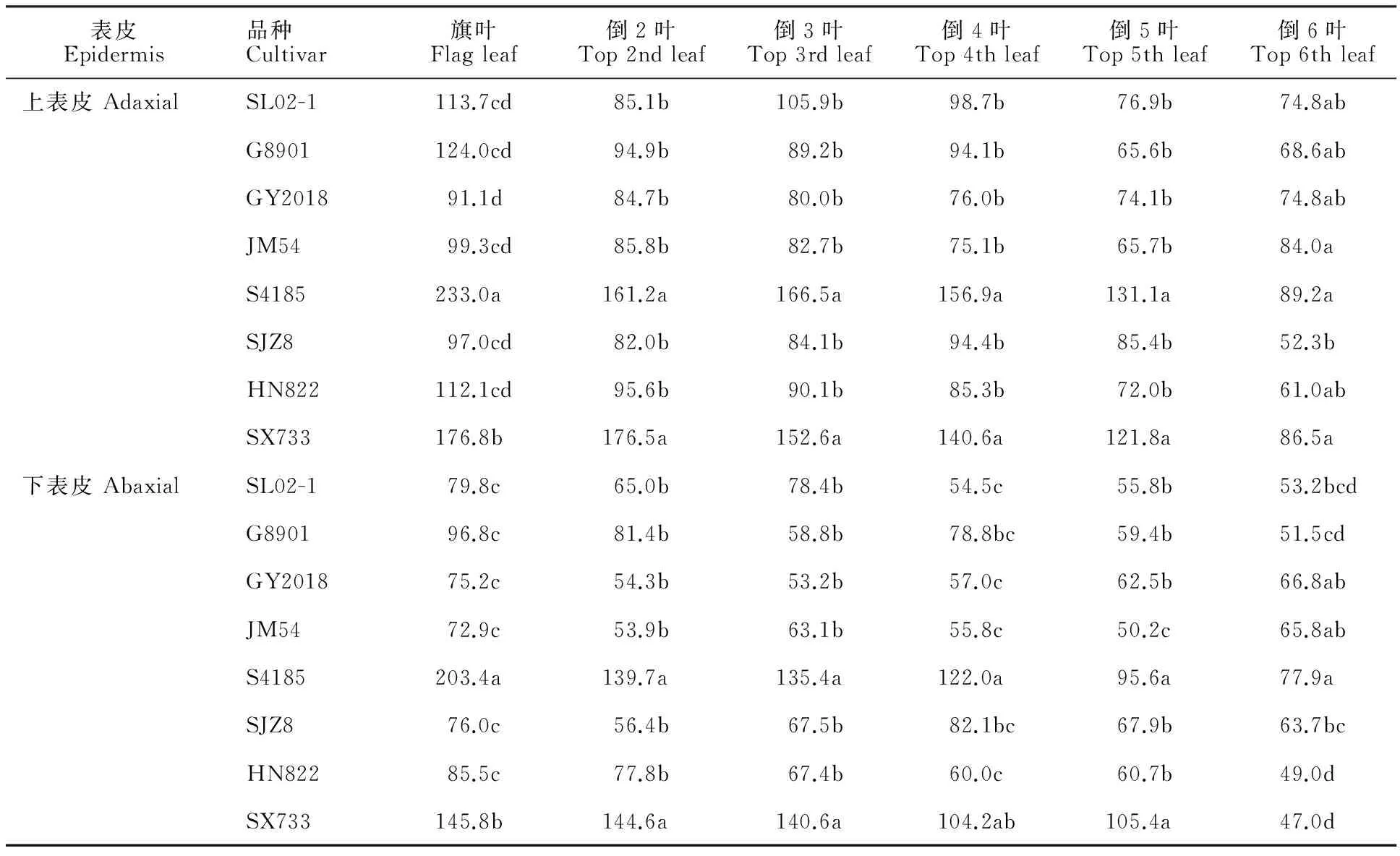
表4 不同冬小麥品種春生葉片的表皮氣孔密度
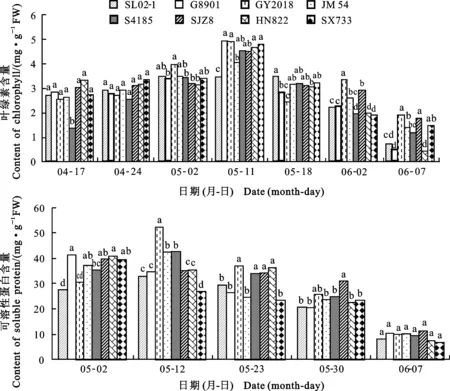
圖1 不同小麥品種旗葉的葉綠素和可溶性蛋白質含量
2.3不同冬小麥品種的光合特性與產量性狀
2.3.1不同冬小麥品種頂三葉的光合特性
由表5可見,不同品種的同葉位葉片比較,S4185、G8901旗葉的平均氣孔導度最高,GY2018最低;倒2葉氣孔導度以SJZ8、GY2018、HN822較高,JM5號和S4185較低;倒3葉氣孔導度以GY8901最高,SX733最低。同一品種不同葉位比較,SL02-1、G8901、JM54、S4185均以旗葉平均氣孔導度最高,其他4個品種以倒2葉氣孔導度最高,各品種均以倒3葉氣孔導度最低。品種間同葉位葉片的平均光合速率差異基本不顯著,其中,旗葉以GY2018的最高,倒2葉以HN822最高,倒3葉以SL02-1最高。各品種不同葉位葉片的平均光合速率都表現為旗葉>倒2葉>倒3葉。
不同品種間比較,頂三葉的光合勢以GY2018最大。不同葉位間比較,GY2018、G8901、SJZ8均以旗葉光合勢最大,其他5個品種以倒2葉光合勢最大,各品種頂三葉的光合勢都以倒3葉最小。不同品種間同葉位比較,旗葉和倒2葉葉源量均以GY2018最大,倒3葉以SX733最大。同一品種不同葉位間比較,除HN822以倒2葉葉源量最大外,其他7個品種均以旗葉葉源量最大;除SX733以倒2葉葉源量最小外,其他品種均以倒3葉葉源量最小。

表5 不同冬小麥品種頂三葉的光合特性
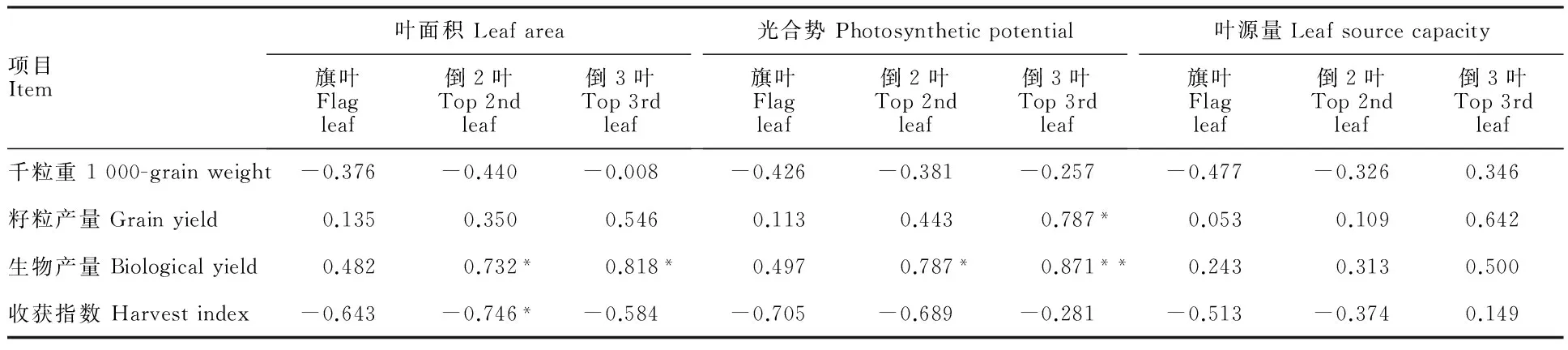
表6 冬小麥品種頂三葉的光合特性與產量構成的相關系數
*:P<0.05;**:P<0.01;n=6
2.3.2不同冬小麥品種頂三葉光合特性與產量構成的相關性
由表6可見,小麥生物產量與頂三葉面積均呈正相關,但在旗葉上相關不顯著,說明旗葉葉面積對生物產量的影響程度不及倒2葉、倒3葉的大。收獲指數與頂三葉面積均呈負相關,但只在倒2葉上顯著。頂三葉面積與千粒重和籽粒產量分別呈負相關和正相關,但均不顯著,說明頂三葉面積與千粒重、籽粒產量的依存性很小。籽粒產量與頂三葉光合勢均呈正相關,但只有與倒3葉的相關性達到顯著水平;生物產量與倒2葉、倒3葉光合勢分別呈顯著和極顯著正相關,與旗葉呈正相關,但不顯著。頂三葉葉源量與產量各性狀的相關性均沒有達到顯著水平。
另外,葉面積、光合勢和葉源量與生物產量和籽粒產量的相關系數均表現為旗葉<倒2葉<倒3葉。這表明,除了重視旗葉光合性能的改善以外,適當提高倒2葉、倒3葉的光合性能,對產量的提高可能起更為重要的作用。
3討 論
3.1小麥葉片的解剖學特征及其生態生理意義
小麥葉片結構組成與其光合性能密切相關。氣孔是小麥葉片與外界進行物質能量交換的主要通道。本研究結果表明,春生葉片氣孔密度小于在后期出生的葉片,這與早春溫度低、葉片代謝弱、稀少的氣孔可增強御寒能力有關[10]。相比之下,后期葉片生長在溫度高、光照強的環境中,稠密的氣孔可提高葉片的蒸騰速率[11],減少強光、高溫對葉片的損傷。這說明氣孔的發育、密度及運動規律與小麥的適應性、抗逆性密切相關[12]。另外,本研究中GY2018的頂三葉面積最大,而其氣孔密度卻較小,表明葉面積的增大一定程度上抵消了氣孔數目的增加[2]。但不同品種表現也有不同,如S4185葉面積并不小,而其氣孔密度最大,這可能有利于植株在干旱條件下通過較強蒸騰作用來吸收水分和保證呼吸[3],從而提高對干旱的適應性。
葉片厚度是葉片解剖特征的另一個重要指標。葉片厚度增加,單位葉面積的葉肉細胞增多[13]。本研究對8個品種的比較表明,小麥旗葉以GY2018較厚,JM54較薄,而品種間光合速率也以GY2018較高,JM54較低。說明較厚的葉片含較多的光合細胞,有助于葉片對光的吸收,從而提高光合速率[13]。可見,在高產條件下,通過水肥調控使葉片展開到適宜面積后,增加葉片厚度對生產的意義可能更大[6]。
本研究中各品種葉片的葉肉細胞,都由具有峰、谷、腰、環的規則環狀細胞及少量不規則細胞組成,而且隨葉位升高,葉片中多環葉肉細胞的比例增高,這些結果都與前人的研究結果一致[5-6]。多環細胞體積較小,單位體積內葉肉細胞總面積較大。葉片進行光合作用,主要依賴于葉片內的葉綠體。葉綠體沿細胞質膜內側排列[14],較大的葉肉細胞總面積有利于在膜內側容納更多的葉綠體,進而提高光合速率。另外,從本研究不同葉位葉片2環葉肉細胞大小看,旗葉的2環細胞高寬積最小或較小,而倒3葉的最大。說明隨葉位升高,相同環數的葉肉細胞變小,在葉片體積一定條件下,增大了葉肉細胞表面積。頂三葉光合能力強,對籽粒貢獻大,與其葉肉細胞光合膜面積大有一定關系。綜上可見,小麥各葉位葉片細胞形態的差異,無論對適應其所擔負的功能或是生長過程中的環境,都具有特定的生理生態意義[15]。本研究灌漿期間干旱少雨(5月11日-6月5日降雨量為3 mm),從相關分析結果看,旗葉對產量的貢獻甚至不如倒2葉、倒3葉。張永平等[16]研究認為,干旱脅迫條件下,適當控制葉面積,相對增加非葉器官面積,有利于小麥產量的提高。該結論與本研究結果一致。
3.2小麥旗葉葉綠素含量和可溶性蛋白質含量動態變化及其相關性
本研究中小麥旗葉生長期中葉綠素含量的變化經歷了低-高-低的過程,這與楊 晴等[17]的研究結果一致。旗葉葉綠素含量高峰出現的早晚、峰值維持時間的長短及單峰與多峰的變化形式,在不同品種中的表現有所不同。這與楊再杰等[18]的結果一致。結合本研究的調查,從旗葉出生到4月24日(挑旗期前后),旗葉葉綠素含量是逐漸提高的過程,從4月24日到6月2日(灌漿末期)是葉綠素高值持續期;灌漿末期后,旗葉葉綠素含量隨葉片的衰亡而迅速下降。小麥旗葉可溶性蛋白含量變化與葉綠素含量變化相似,但也有所不同,旗葉葉綠素含量最大值出現在5月10日前后,而不同品種旗葉可溶性蛋白含量的最大值陸續在5月7日-12日出現。另外,小麥生育末期葉綠素含量會迅速下降,而可溶性蛋白含量下降相對緩慢,葉片在失綠后仍含有一定量的可溶性蛋白。可見,小麥生育末期葉片停止光合活動后,包括可溶性蛋白在內的光合物質仍有活性。說明小麥產量形成不僅與葉片葉綠素的含量總體水平有關,還與光合產物再分配密切相關[19]。
3.3冬小麥群體光合特性對產量形成的影響
前人研究表明,小麥不同葉位葉片的光合產物對籽粒的貢獻不同,頂三葉光合能力強,對籽粒產量貢獻較大[11]。本研究中不同品種各葉位葉片葉肉細胞的平均環數均以旗葉最高,平均光合速率也以旗葉最高。綜合分析本研究與他人研究結果,環數較多的葉肉細胞個體小,葉片單位體積內葉肉細胞總表面積高,進而擴大了光合膜面積[6,20]。說明旗葉具有良好的光合性能,與其葉肉細胞組成密不可分[20]。另外,在品種間,S4185、SL02-1旗葉細胞環數較高,但其葉綠素含量、可溶性蛋白含量和平均光合速率均較低。而GY2018 旗葉葉肉細胞環數雖低,但其葉面積大,葉綠素高值持續期長,可溶性蛋白含量高,光合速率與葉源量均較高,最終生物產量也最高。但由于GY2018收獲指數較低,與其他品種比較,其產量并不是最高。相關分析發現,頂三葉的葉面積、光合勢、葉源量等參數與生物產量、籽粒產量呈不同程度的正相關,這與王成雨等[21]的研究結果一致。具體來看,相關系數表現為旗葉<倒2葉<倒3葉。綜合來看,具有理想的葉片細胞顯微結構并不意味著產量性狀一定優越,光合器官細胞形態與產量構成屬于不同層次的性狀,不一定具有一一對應的關系。結合品種特性,采用合理的栽培技術,處理好內部與外部、群體與個體的矛盾,使光合產物最大限度產出并有效轉移到籽粒中去,才是獲取高產的有效保證[22-23]。
本研究還發現,在一定光合生產能力條件下,葉片顯微結構是一個相互協調、相互制約的平衡系統。這與小麥產量構成因素的相互關系相似[24]。如SL02-1、G8901、S4185等小麥品種旗葉的葉肉細胞平均環數較多,2環細胞的高、寬、積也較小,而且旗葉較薄。相比之下,SX733、GY2018葉肉細胞平均環數較少,2環細胞的高、寬、積也較大,而且旗葉較厚。葉片不同葉位間比較也有此變化趨勢。品種間葉片光合勢、葉源量的差異與葉肉細胞和葉綠素含量表現并不完全一致。說明造成品種之間產量差異的主要原因不能單從細胞形態組成等方面闡釋。品種間產量的主要差異表現在生物產量和收獲指數上,光合物質生產和轉運的差異是造成各品種產量差的主要原因[19,21]。
參考文獻:
[1]Li H W,Jiang D,Wollenweber B,etal.Effects of shading on morphology,physiology and grain yield of winter wheat [J].EuropeanJournalofAgronomy,2010,33:267-275.
[2]劉 瑩,李雁鳴,張立言.華北地區小麥品種更替過程中葉片細胞形態和光合性能演替規律的研究Ⅰ.葉片細胞形態的演替[J].河北農業大學學報,1998,21(3):7-11.
Liu Y,Li Y M,Zhang L Y.The studies on the changes of leaf cell morphology and photosynthetic performances during evolutional succession of winter wheat cultivars in North China Ⅰ.Changes of leaf cell morphology [J].JournalofAgricultureUniversityofHebei,1998,21(3):7-11.
[3]李芳蘭,包維楷.植物葉片形態解剖結構對環境變化的響應與適應[J].植物學通報,2005,22(增刊):118-127.
Li F L,Bao W K.Responses of the morphological and anatomical structure of the plant leaf to environmental change [J].ChineseBulletinofBotany,2005,22(S):118-127.
[4]李雁鳴,劉 瑩,張立言.華北地區小麥品種更替過程中葉片細胞形態和光合性能演替規律的研究Ⅱ.旗葉光合性能的演替[J].河北農業大學學報,1998,21(4):6-11.
Li Y M,Liu Y,Zhang L Y.Study on the changes of leaf cell morphology and photosynthetic performances during evolutional succession of winter wheat cultivars in North China Ⅱ.Changes in photosynthetic performances of flag leaves [J].JournalofAgricultureUniversityofHebei,1998,21(4):6-11.
[5]劉永康,李明軍,李景原,等.小麥旗葉直立轉披動態過程對其高光效的影響[J].科學通報,2009,54(15):2205-2211.
Liu Y K,Li M J,Li J Y,etal.Effect of vertical turning with dynamic process on high light efficiency of wheat flag leaf [J].ChineseScienceBulletin,2009,54(15):2205-2211.
[6]李金霞,李 云,李瑞奇,等.4種植物生長調節劑對冬小麥旗葉細胞形態的影響[J].河北農業大學學報,2010,33(1):1-5,11.
Li J X,Li Y,Li R Q,etal.Effect of four plant growth regulators on the cell morphology of flag leaves of winter wheat [J].JournalofAgricultureUniversityofHebei,2010,33(1):1-5,11.
[7]段續川.植物細胞和細胞器的固定、水解、分離和染色的革新[J].植物學報,1959,8(1):1-14.
Tuan H C.Renovation in technique for the study of plant cells and their organoids [J].ActaBotanicaSinica,1959,8(1):1-14.
[8]趙世杰,劉華山,董新純.植物生理學實驗指導[M].北京:中國農業科技出版社,1998:57-157.
Zhao S J,Liu H S,Dong X C.Experimental Guide of Plant Physiology [M].Beijing:China Agricultural Science and Technology Press,1998:57-157.
[9]Bradford M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding [J].AnalyticalBiochemistry,1976,72:248-254.
[10]鄭丕堯,李雁鳴.不同播期生態條件下燕麥葉片細胞形態的觀察[J].作物學報,1992,18(3):183-190.
Zheng P Y,Li Y M.Observation on the morphology of leaf blade cells in oats under different sowing-date conditions [J].ActaAgronomicaSinica,1992,18(3):183-190.
[11]苗 芳,張嵩午.小麥植株發育過程中頂三葉結構的變化特征[J].西北農林科技大學學報(自然科學版),2004,32(10):15-19.
Miao F,Zhang S W.Change characteristics of the top-three leaves' structure in wheat development process [J].JournalofNorthwestA&FUniversity(NaturalScienceEdition),2004,32(10):15-19.
[12]Wang H C,Ngwenyama N,Liu Y D,etal.Stomatal development and patterning are regulated by environmentally responsive mitogen-activated protein kinases inArabidopsis[J].PlantCell,2007,19:63-73.
[13]Wilson D.Assimilation of lolium in relation to leaf mesophyll [J].Nature,1967,214(3):989-993.
[14]苗 芳,張嵩午,王長發,等.低溫小麥種質葉片結構及某些生理特性[J].應用生態學報,2006,17(3):408-412.
Miao F,Zhang S W,Wang C F,etal.Leaf structure and some physiologic characteristics of low temperature wheat germplasm [J].ChineseJournalofAppliedEcology,2006,17(3):408-412.
[15]李雁鳴,鄭丕堯,王瑞舫.高粱葉片及其它營養器官細胞形態的初步觀察[J].作物學報,1994,20(1):106-113.
Li Y M,Zheng P Y,Wang R F.Preliminary observation on the cell morphology of leaf blades and other vegetative organs in sorghum [J].ActaAgronomicaSinica,1994,20(1):106-113.
[16]張永平,張英華,王志敏.不同供水條件下冬小麥葉與非葉綠色器官光合日變化特征[J].生態學報,2011,31(5):1312-1322.
Zhang Y P,Zhang Y H,Wang Z M.Photosynthetic diurnal variation characteristics of leaf and non-leaf organs in winter wheat under different irrigation regimes [J].ActaEcologicaSinica,2011,31(5):1312-1322.
[17]楊 晴,劉奇勇,白 巖,等.冬小麥不同葉層葉綠素和可溶性蛋白對氮磷肥的響應[J].麥類作物學報,2009,29(1):128-133.
Yang Q,Liu Q Y,Bai Y,etal.Responses of chlorophyll and soluble protein in different leaf layers of winter wheat to N and P nutrients [J].JournalofTriticeaeCrops,2009,29(1):128-133.
[18]楊再潔,陳 阜,史磊剛,等.華北平原不同年代小麥品種旗葉光合特性對水分虧缺的影響[J].作物學報,2013,39(4):693-703.
Yang Z J,Chen F,Shi L G,etal.Responses of photosynthetic characteristics of winter wheat cultivars released in different decades to water deficit in North China Plain [J].ActaAgronomicaSinica,2013,39(4):693-703.
[19]黃 玲,高 陽,邱新強,等.水分對不同年代主栽冬小麥品種光合特性的影響[J].麥類作物學報,2013,33(3):495-502.
Huang L,Gao Y,Qiu X Q,etal.Effects of water on photosynthetic characteristics of major winter wheat cultivars at different decades [J].JournalofTriticeaeCrops,2013,33(3):495-502.
[20]劉月蘭,于振文,張永麗,等.拔節期和開花期不同土層深度測墑補灌對北方小麥旗葉葉綠體超微結構和熒光特性的影響[J].中國農業科學,2014,47(14):2751-2761.
Liu Y L,Yu Z W,Zhang Y L,etal.Effects of supplemental irrigation based on the measurement of moisture content in soil layers at jointing and anthesis stages on the chloroplast ultramicrostructure and chlorophyll fluorescence parameters of flag leaves of winter wheat [J].ScientiaAgriculturaSinica,2014,47(14):2751-2761.
[21]王成雨,代興龍,石玉華,等.花后小麥葉面積指數與光合和產量關系研究[J].植物營養與肥料學報,2012,18(1):27-34.
Wang C Y,Dai X L,Shi Y H,etal.Effects of leaf area index on photosynthesis and yield of winter wheat after anthesis [J].PlantnutritionandFertilizerScience,2012,18(1):27-34.
[22]郝啟飛,陳 煒,鄧西平.不同栽培模式對長武塬區冬小麥干物質積累轉運的影響[J].水土保持研究,2011,18(3):121-125.
Hao Q F,Chen W,Deng X P.Dry matter accumulation and transportation of winter wheat under different cultivation models on Changwu Tableland [J].ResearchofSoilandWaterConservation,2011,18(3):121-125.
[23]王紅光,李東曉,李雁鳴,等.河北省10 000 kg·hm-2以上冬小麥產量構成及群個體生育特性[J].中國農業科學,2015,48(14):2718-2729.
Wang H G,Li D X Li Y M,etal.Yield components and population and individual characteristics of growth and development of winter wheat over 10 000 kg·hm-2in Hebei province [J].ScientiaAgriculturaSinica,2015,48(14):2718-2729.
[24]黨紅凱,李瑞奇,李雁鳴,等.超高產冬小麥四種微量元素的積累及其與產量性狀的關系[J].麥類作物報,2012,32(2):326-332.
Dang H K,Li R Q,Li Y M,etal.Accumulation of four microelements and their relationship with yield traits in super high-yield winter wheat [J].JournalofTriticeaeCrops,2012,32(2):326-332.
收稿日期:2015-10-05修回日期:2015-11-19
基金項目:國家科技支撐計劃項目(2011BAD16B08,2012BAD04B06,2013BAD07B05);國家小麥產業技術體系建設專項(CARS-3-2-3)
通訊作者:李雁鳴(E-mail:nxzwst@hebau.edu.cn或liym315@126.com)
中圖分類號:S512.1;S311
文獻標識碼:A
文章編號:1009-1041(2016)06-0742-10
Relationship among Leaf Micro-structure,Photosynthetic Characteristics and Yield Traits of Winter Wheat
DANG Hongkai1,2,LI Ruiqi1,LI Yanming1,LI Xiaoshuang1,MENG Jian1
(1.College of Agronomy,Hebei Agricultural University/Key Laboratory of Crop Growth Regulation of Hebei Province,Baoding,Hebei 071000,China; 2.Dryland Farming Institute,Hebei Academy of Agriculture and Forestry Sciences,Hengshui,Hebei 053000,China)
Abstract:In order to verify the relationship among photosynthetic characteristics,leaf micro-structure and yield traits of winter wheat,a field experiment was conducted in Gaocheng,Hebei pronvince,using eight wheat cultivars as experiment materials.The main results showed that leaves at different positions on the main stem of wheat mainly composed of regular mesophyll cells.The amounts of links of regular mesophyll cells ranged from 1.7 to 4.5.The average amounts of links of mesophyll cells and the stomatal density on adaxial and abaxial epidermis all had tendencies to increase with rising leaf positions for all cultivars.The maximum values of chlorophyll content in flag leaves appeared at around May 10 for all wheat cultivars.The dynamics of soluble protein content of flag leaves were similar to those of chlorophyll content with the maximum contents appeared from May 7 to May 20 in different cultivars.The average amounts of links of mesophyll cells and photosynthetic rates of flag leaves were highest among all leaves of wheat plants.The values of photosynthetic potential and leaf source capacity,however,were not entirely consistent with those of the link amounts of mesophyll cells and chlorophyll content among wheat cultivars.Yield differences among cultivars were mainly due to the different biological yield and harvest index.Biological yield was significantly positive correlated(at 0.05 level) to photosynthetic potential of top 2nd leaf,and also significantly positive correlated(at 0.01 level) to photosynthetic potential of top 3rd leaf.According to these results,good photosynthetic performance and high efficiency of photosynthetic products translocation into grains are essential features for a wheat cultivar to achieve high yield.
Key words:Winter wheat; Leaves; Anatomical characters; Photosynthetic characteristics; Yield traits
網絡出版時間:2016-05-30
網絡出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160530.1536.020.html
第一作者E-mail:wheatcrop@126.com.
