利用90K基因芯片進行小麥旗葉相關性狀的QTL定位
2016-07-29 07:47:30連俊方張德強武炳瑾宋曉朋馬文潔周麗敏孫道杰
麥類作物學報 2016年6期
連俊方,張德強,武炳瑾,宋曉朋,馬文潔,周麗敏,馮 毅,孫道杰
(西北農林科技大學農學院,陜西楊凌 712100)
?
利用90K基因芯片進行小麥旗葉相關性狀的QTL定位
連俊方,張德強,武炳瑾,宋曉朋,馬文潔,周麗敏,馮 毅,孫道杰
(西北農林科技大學農學院,陜西楊凌 712100)
摘要:為了發掘影響小麥旗葉相關性狀的QTL,以小麥骨干親本周8425B與優良品種小偃81構建的包含102個家系的重組自交系(Recombinant inbred line,RIL)為材料,采用小麥90K SNP基因芯片技術和SSR標記對其進行分子標記檢測,構建含有全基因組SNP和SSR 標記的高密度遺傳圖譜,并在4個環境下對小麥旗葉相關性狀QTL進行檢測。結果表明,所構建圖譜含有6 949對多態性標記,其中,SNP標記6 910對,SSR標記39對,覆蓋染色體總長度4 839.9 cM,標記間平均距離0.7 cM;A、B和D染色體組分別有2 085、4 677和187對標記,分別占總標記數的30.0%、67.5%和2.7%,標記間平均距離分別為1.0、0.6和0.8 cM。采用完備復合區間作圖法共檢測到22個旗葉性狀加性效應 QTL,10個旗葉長 QTL 分布于 2A、3B、4B、5A、6B和7B染色體上,解釋表型變異 7.900%~24.098%,除 Qfll2A-1能在2個環境中檢測到外,其余均為單環境QTL;4個旗葉寬 QTL分布于2A、3A和5B染色體上,解釋表型變異9.080%~16.540%,其中, Qflw2A-1在3個環境中均能檢測到,解釋表型變異12.483%~16.540%,為1個穩定的主效QTL;8個旗葉面積 QTL 分布于2A、3B、4B、5A、6B和7A染色體上,解釋表型變異9.310%~30.498%,其中,3個QTL位于5A染色體上。此外,鑒定出3個分布于2A、5A和6B染色體上的 QTL富集區段。
關鍵詞:小麥;重組自交系;旗葉性狀;SNP;QTL
小麥旗葉對小麥的產量具有重要的影響[1],在抽穗開花后對粒重的貢獻率超過30%,而在發育后期,旗葉的凈光合物質幾乎全部運輸到籽粒中去[2]。小麥旗葉的長、寬及葉面積等相關性狀與其產量具有顯著的正相關關系[3-7]。因此,對小麥旗葉相關性狀的研究將會對小麥產量育種提供理論指導。
目前,國內外有關旗葉性狀的QTL研究已有諸多報道。常 鑫等[8]和趙 朋等[9]分別利用小偃81/西農1376和春小麥寧春4號/寧春27構建的重組自交系(Recombinant inbred line,RIL)群體,在多環境中檢測出位于不同染色體的多個控制旗葉長、寬和面積的QTL;閆 雪等[10]利用旱選10號和魯麥14構建的包含150個家系的DH群體,分別在干旱脅迫與正常灌溉條件下檢測到多個控制旗葉長、寬和面積的QTL;Jia等[11]在5A染色體上發現一個控制旗葉寬的QTL;Xue等[12]利用以綿陽99-323 和 PH691為遺傳背景的近等基因系(NIL)衍生的次級F2群體將Jia等[11]發現的 Qflw.naw-5A精細定位并發現其與5A染色體上的 Fhb5緊密連鎖。然而,這些研究都是利用傳統的SSR等標記構建的低密度遺傳圖譜,標記間距離大,檢測方法費時、繁瑣,人工成本高,很難應用于育種實踐。基因芯片技術已經廣泛應用于醫學領域[13-15]以及水稻[16-17]、大麥[18]等谷類作物的研究中。Zhang等[19]利用SNP基因芯片定位到控制水稻旗葉寬的主效QTL, Bertholdsson等[20]利用SNP基因芯片在大麥中發現5個影響光量子產量的QTL。近年來,基于Illumina 技術平臺開發了小麥9K和90K SNP基因芯片[21]。Wu等[22]利用9K SNP芯片和SSR標記在燕大1817與北農6號組成的RIL群體中定位出與旗葉性狀有關的QTL。然而目前利用90K芯片對旗葉相關性狀的QTL檢測還鮮見報道。
本研究以利用周8425B和小偃81構建的包含102個家系的RIL群體為研究材料,采用小麥90K SNP基因芯片技術和SSR標記對其進行分子標記檢測,構建含有全基因組SNP和SSR 標記的高密度遺傳圖譜,并在4個環境下對小麥旗葉相關性狀QTL進行檢測,以期為小麥旗葉長、寬和面積等數量性狀的基因定位、分子標記開發、功能基因的精細定位以及株型育種提供理論依據。
1材料與方法
1.1試驗材料
供試材料為周8425B(母本,是由周口市農科院育成的高產、抗病新種質)與小偃81(父本,為李振聲院士育成的高產、優質、抗病、半冬性、多穗型中熟品種)雜交后經單粒傳法得到的含 102個家系的 F8~F9代RIL群體,由西北農林科技大學農學院王 輝教授課題組創制并保存。
1.2田間種植和性狀調查
2013-2014年度和2014-2015年度將RIL群體及其親本種植于陜西楊凌和河南安陽兩地。兩年均采用完全隨機區組設計,每個株系種植2行,2次重復,行長2 m,行距0.25 m,株距3 cm。田間管理按照當地大田生產管理標準進行。于小麥開花后20 d(灌漿期)測定其旗葉長、寬,每個株系隨機抽取6株進行測量,數據讀取至小數點后兩位,然后取其均值作為該品系旗葉長、寬值,旗葉面積采用宋荷仙等[23]的方法,旗葉面積=葉長×葉寬×0.83。
1.3DNA 的提取和分子標記檢測
采用CTAB法[24]提取102個RIL家系及其親本DNA,將其保存在TE中,用分光光度計定量,并將樣品用TE調到標準濃度50 ng·μL-1,然后用0.8%的瓊脂糖凝膠進行電泳。樣品電泳主帶清晰,大小在10 kb以上,沒有明顯降解,總量 1 μg 以上,A260/A280=1.7~2.1,為合格樣品。
SNP標記由北京博奧晶典生物技術有限公司利用 Illumina SNP Genotyping技術測試平臺,使用微珠芯片技術(BeadArray)進行檢測,其多態性使用 Genomestudio v1.0 軟件進行分析。
SSR引物由上海生工生物工程有限公司合成,引物序列信息從GrainGene 2.0(http://wheat.pw.usda.gov/GG2/index.shtml)獲得,利用兩親本進行多態性標記篩選,用多態性引物在RIL群體中進行檢測。
1.4遺傳連鎖圖的構建和QTL定位
利用軟件QTL IciMapping 4.0和QTL Mapchart 2.1構建分子標記的連鎖圖。利用IBM SPSS statistics 對表型數據進行偏峰度及相關性分析。在此基礎上,結合表型數據,使用QTL IciMapping 4.0軟件,采用完備區間作圖法(ICIM-ADD)[25]進行QTL檢測,其中,LOD閾值為2.5,掃描步長為1 cM,逐步回歸進入的概率(PIN)為0.001,缺失表型不用于QTL作圖(表型缺失的基因型數據仍會使用)。QTL 的命名參考 McIntosh等[26]的方法,按照性狀及染色體位置進行命名,旗葉長命名為Qfll,旗葉寬命名為Qflw,旗葉面積命名為Qfla,如 Qfll3A-1表示位于3A染色體上控制旗葉長且位于第一個標記區間的QTL。
2結果與分析
2.1遺傳圖譜構建
在親本周8425B和小偃81中進行標記檢測,共篩選出8 650對多態性標記,其中包含187對SSR標記和8 463對SNP標記。經過連鎖分析,共有6 949對標記構建到圖譜中,其中包含6 910對SNP標記和39對SSR標記,覆蓋染色體總長度4 839.9 cM,標記間平均距離0.7 cM。構建的遺傳圖譜中,A 染色體組有2 085對標記,占總標記數的30.0%,全長2 074.1 cM,標記間平均距離為1.0 cM;B染色體組多態性標記最多,共4 677對,占標記總數的67.3%,全長2 624.6 cM,標記間平均距離僅為0.6 cM;D染色體組標記數目最少,共187對,占標記總數的2.7%,染色體覆蓋長度最短,全長僅有141.2 cM,標記間平均距離0.8 cM。
2.2表型性狀變異分析
將2013-2014年度種植在陜西楊凌(E1)、河南安陽(E2)和2014-2015年度種植在陜西楊凌(E3)、河南安陽(E4)的各親本及RIL家系的表型性狀進行統計分析,結果(表1)表明,親本周8425B的旗葉長、寬和面積在多環境中表現均大于親本小偃81,且差異具有統計學意義。RIL群體各環境下表現出明顯的超親遺傳,各家系3個性狀表型變異豐富,且呈現連續變異,偏、峰度絕對值基本都小于1,符合正態分布,表現為多基因控制的數量性狀遺傳的特點,適合做QTL分析。雖然2014-2015年度種植于安陽的旗葉面積的峰度稍大于1,但根據李慧慧等[27]數量性狀作圖的要求,表型性狀的非正態性分布并不影響QTL作圖。
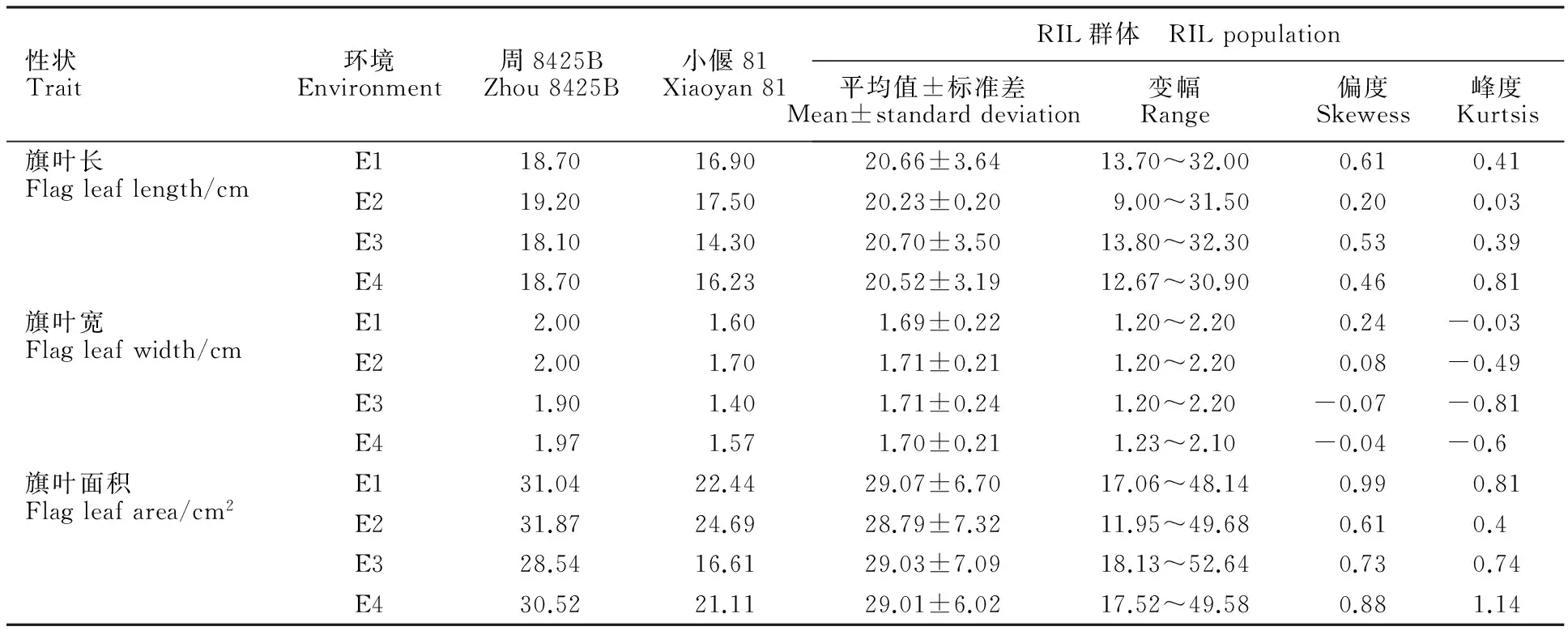
表1 不同環境下RIL群體及其親本的旗葉長、寬和面積
2.3旗葉長、寬和面積的QTL分析
在4個環境中共檢測到22個旗葉長、寬和面積相關的QTL,分別位于2A、3A、3B、4B、5A、5B、6B、7A和7B染色體上,其中3個QTL在多個環境中均檢測到。A、B、D基因組分別含有10、12和0個QTL;第1至7同源群上分別檢測到0、3、4、2、6、5和2個QTL(表3,圖1)。
2.3.1旗葉長QTL
4個環境中共檢測到10個控制旗葉長的QTL,分別位于2A、3B、4B、5A、6B和7B染色體上,解釋表型變異7.900%~24.080%。除 Qfll2A-1外,其他QTL只能在1個環境中檢測到,說明旗葉長的遺傳基礎復雜且受環境的影響很大。 Qfll2A-1在E3和E4兩個環境中檢測到,位于標記Tdurum_contig64391_231和RAC875_c58503_182之間,標記間距離僅為0.5 cM。
2.3.2旗葉寬QTL
4個環境中共檢測到4個控制旗葉寬的QTL,分別位于2A、3A和5B染色體上,解釋表型變異9.080%~16.540%。其中, Qflw2A-2在E2、E3、E4多個環境中均檢測到,位于標記RAC875_c58006_352和BS00094574_51之間,標記間距離為1.0 cM。
2.3.3旗葉面積QTL
4個環境中共檢測到8個控制旗葉面積的QTL,分別位于2A、3B、4B、5A、6B和7A染色體上,解釋表型變異9.310%~30.498%。其中, Qfla4B-1在E3、E4中均檢測到,位于標記RAC875_c18200_929和Excalibur_c47209_87之間,標記間距離為0.5 cM。此外,在E2和E4中檢測到 Qfla5A-1、 Qfla5A-2和 Qfla5A-3均位于5A染色體200.0 cM附近。
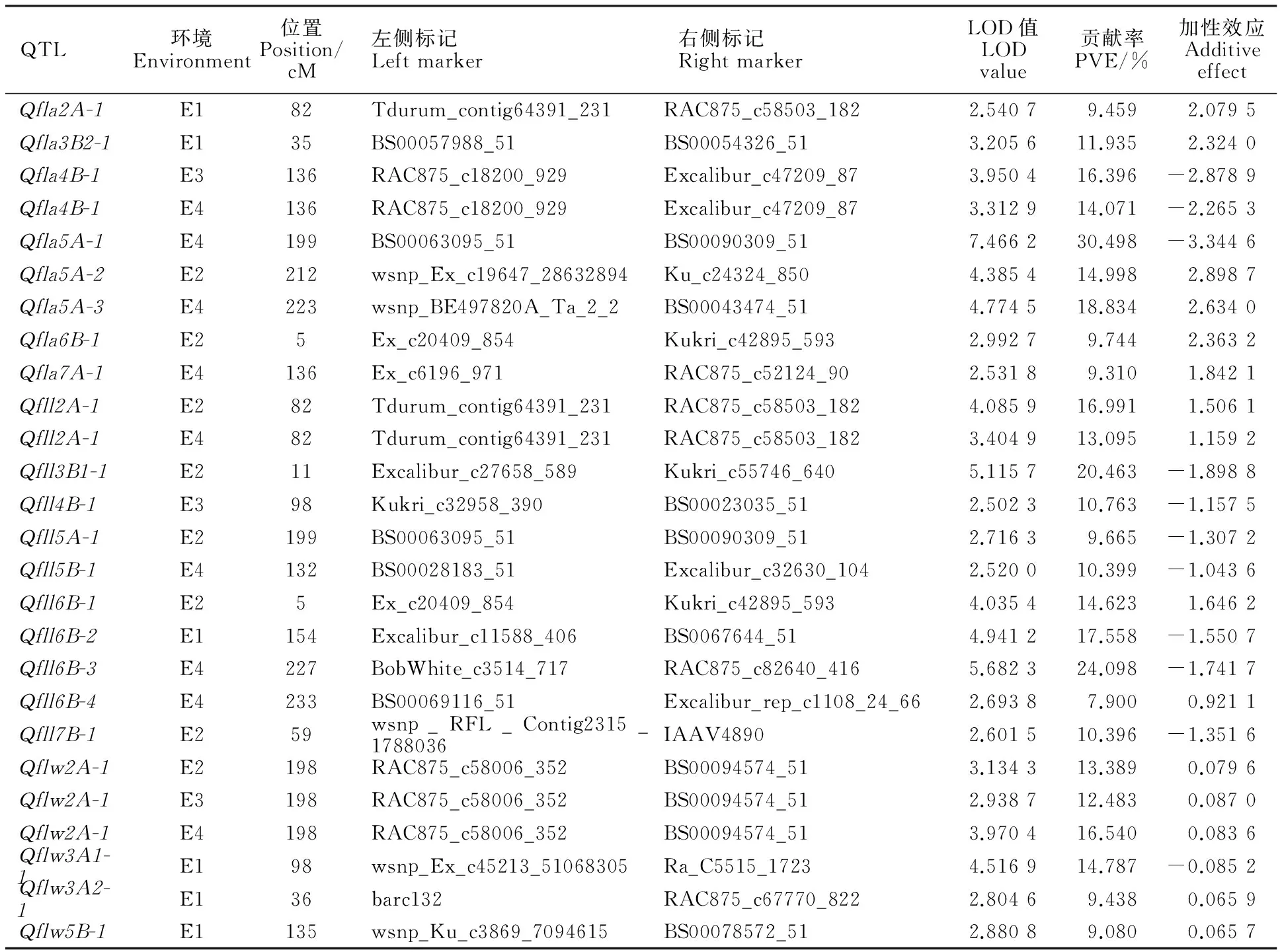
表2 不同環境中旗葉長、寬和面積的QTL定位結果



3討 論
3.1QTL定位結果一致性
數量性狀受多基因控制,且受環境影響較大,即使相同群體不同年份檢測出的QTL也存在較大差異。目前已有利用SSR標記和9K芯片對旗葉性狀的QTL定位的研究,但由于遺傳圖譜密度小,定位出的QTL標記間距離大,存在的假陽性QTL多,很難應用于育種實踐。然而,利用90K基因芯片對小麥旗葉性狀進行QTL的發掘卻很少有學者進行研究。本研究利用90K SNP芯片構建遺傳圖譜,所用標記和前人研究所用標記不同,很難用于比較是否為同一QTL,但仍具有一定的參考價值。
Qfla3B2-1是位于小麥3B染色體上控制旗葉面積的QTL,Wu等[22]和常 鑫等[8]也在該染色體上檢測出控制旗葉長的QTL,但位置相距較遠,應該不是同一QTL。在4B染色體上檢測到控制旗葉長的 Qfll4B-1,Fan等[28]在4B染色體上發現3個旗葉長QTL,但由于所用標記不同,不能確定是否存在相同QTL。位于5B染色體上的 Qfll5B-1與Fan等[28]在5B染色體上發現旗葉長相關QTL的表型貢獻率都較低,不是主效QTL。在3個環境中均檢測到位于6B染色體上的旗葉長QTL,其中3個QTL都在200.0 cM附近的位置,該區段應該為1個控制旗葉長的染色體簇,趙 朋等[9]也在該染色體上發現旗葉長QTL。在2A染色體的RAC875_c58006_352和BS00094574_51標記之間檢測到1個旗葉寬QTL,在3個環境中均穩定表達,解釋表型變異13.389%~16.540%,是1個主效QTL。 Qflw5B-1為在5B上控制旗葉寬的QTL,Fan等[28]和Jia等[11]也在同一染色體上發現旗葉寬QTL。 Qfla4B-1為在4B染色體上136.0 cM位置控制旗葉面積的QTL,在2個環境中均穩定表達,表型貢獻率為14.071%~16.396%,說明這是1個穩定的主效QTL,Fan等[28]也在4B染色體上檢測出旗葉面積QTL,但臨近的標記不同,不能確定是否為同一位點。在5A染色體200.0 cM附近存在3個控制旗葉面積的QTL,Wu等[22]在5A上200.0 cM處也檢測出1個旗葉面積QTL,且在多環境中均能檢測到,預測該位點存在穩定表達的QTL,但是否為同一位點還很難判斷。本研究發現的旗葉面積 Qfla6B-1與Fan等[28]在6B染色體上發現旗葉面積QTL均為微效QTL。位于7A染色體上的 Qfla7A-1與常 鑫等[8]發現旗葉面積QTL在7A上的位置相近,預測該位置附近存在影響旗葉面積的基因。 Qfll2A-1為在2A染色體上Tdurum_contig64391_231與RAC875_c58503_182標記之間存在的1個控制旗葉長的QTL,該位點在E2和E4兩個環境中均能檢測出來,是一個新發現的位點,標記間距離僅為0.5 cM,但目前只在一個群體檢測出來,需要在多個群體中進一步驗證,以期能夠早日應用于育種實踐。
3.2QTL聚集區及一因多效QTL
本研究在2A、5A和6B染色體上檢測出3個QTL聚集區及一因多效QTL。高度相關的性狀之間表現出區域化分布現象或存在一些共同的QTL[29]。位于2A染色體標記Tdurum_contig64391_231與RAC875_c58503_182之間的QTL為同時控制旗葉長和旗葉面積的一因多效QTL,且該QTL在多個環境中均能檢測到,為1個穩定的QTL;在5A染色體上3個相近的區間BS00063095_51-BS00090309_51、wsnp_Ex_c19647_28632894-Ku_c24324_850和wsnp_BE497820A_Ta_2_2-BS00043474_51包含1個旗葉長和3個旗葉面積的QTL,其中,BS00063095_51-BS00090309_51之間的QTL為同時控制旗葉長和旗葉面積的一因多效QTL;在6B染色體上Ex_c20409_854-Kukri_c42895_593區間的QTL同時影響旗葉長和旗葉面積,為一因多效QTL。在5A染色體上檢測到3個旗葉面積QTL,且3個QTL距離在24.0 cM內,推測這一區段為控制旗葉面積的基因簇。此外,6B染色體上檢測出4個旗葉長相關QTL。
本研究發現控制旗葉相關性狀的QTL存在QTL聚集區和一因多效現象,這說明旗葉不同性狀之間具有一定的相關性。控制相關性狀的基因更傾向于形成基因聚合區域,而非零散分布,這也許是物種進化帶來的結果,同時也對聚合育種提供了很大的便利。
參考文獻:
[1]Saha C S,鄭康樂.小麥旗葉節以上不同結構的光合面積與籽粒產量的關系 [J].麥類作物學報,1982(5):28-31.
Saha C S,Zheng K L.The relationship between photosynthetic area and grain yield of wheat with different structure of flag leaf [J].JournalofTriticeaeCrops,1982(5): 28-31.
[2]Simón M R.Inheritance of flag-leaf angle,flag-leaf area and flag-leaf area duration in four wheat crosses [J].TheoreticalandAppliedGenetics,1999,98(2):310-314.
[3]Stahli D,Perrissin-Fabert D,Blouet A,etal.Contribution of the wheat(TriticumaestivumL.) flag leaf to grain yield in response to plant growth regulators [J].PlantGrowthRegulation,1995,16(3):293-297.
[4]Mcneal F H,Berg M A.Flag leaf area in five spring wheat crosses and the relationship to grain yield [J].Euphytica,1977,26(3):739-744.
[5]成東梅,彭 濤,高 燕,等.高產小麥旗葉與穗粒重關系的研究 [J].安徽農業科學,2007,35(36): 11798-11799.
Cheng D M,Peng T,Gao Y,etal.Study on the relationship between the flag leaves and ear grain weight of high-yield wheat [J].JournalofAnhuiAgriculturalSciences,2007,35(36):11798-11799.
[6]傅兆麟,馬寶珍,王光杰,等.小麥旗葉與穗粒重關系的研究 [J].麥類作物學報,2001,21(1):92-94.
Fu Z L,Ma B Z,Wang G J,etal.Relationship between the flag leaf and the grain weight per spike in wheat [J].JournalofTriticeaeCrops,2001,21(1):92-94.
[7]郭翠花,高志強,苗果園.花后遮陰對小麥旗葉光合特性及籽粒產量和品質的影響 [J].作物學報,2010,36(4): 673-679.
Guo C H,Gao Z Q,Miao G Y.Effect of shading at post flowering on photosynthetic characteristics of flag leaf and response of grain yield and quality to shading in wheat [J].ActaAgronmicaSinica,2010,36(4):673-679.
[8]常 鑫,李法計,張兆萍,等.小麥旗葉長、寬及面積的QTL分析 [J].西北植物學報,2014,34(5):896-901.
Chang X,Li F J,Zhang Z P,etal.Mapping QTLs for flag leaf length,width and area in wheat [J].ActaBotanicaBoreali-OccidentaliaSinica,2014,34(5):896-901.
[9]趙 朋,徐 鳳,蔣文慧,等.春小麥旗葉長度、寬度及葉綠素含量QTL分析 [J].麥類作物學報,2015,35(5):603-608.
Zhao P,Xu F,Jiang W H,etal.Quantitative trait loci analysis of flag leaf length width and chlorophyll content of spring wheat [J].JournalofTriticeaeCrops,2015,35(5):603-608.
[10]閆 雪,史雨剛,梁增浩,等.小麥旗葉形態相關性狀的QTL定位 [J].核農學報,2015,29(7):1253-1259.
Yan X,Shi Y G,Liang Z H,etal.QTL mapping for morphological traits of flag leaf in wheat [J].JournalofNuclearAgriculturalSciences,2015,29(7):1253-1259.
[11]Jia H,Wan H,Yang S,etal.Genetic dissection of yield-related traits in a recombinant inbred line population created using a key breeding parent in China’s wheat breeding [J].TheoreticalandAppliedGenetics,2013,126(8):2123-2139.
[12]Xue S,Xu F,Li G,etal.Fine mapping TaFLW1,a major QTL controlling flag leaf width in bread wheat(TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2013,126(8):1941-1949.
[13]Arnold M,Hartsperger M L,Baurecht H,etal.Network-based SNP meta-analysis identifies joint and disjoint genetic features across common human diseases [J].BioMedCentralGenomics,2012,13:490.
[14]Wang Y,Sung P,Lin P,etal.A multi-SNP association test for complex diseases incorporating an optimal P-value threshold algorithm in nuclear families [J].BioMedCentralGenomics,2015,16(1):381-481.
[15]Chen X,Gong X,Yan N,etal.RNASET2 tag SNP but not CCR6 polymorphisms is associated with autoimmune thyroid diseases in the Chinese Han population [J].BioMedCentralMedicalGenetics,2015,16(1):11.
[16]Shen B,Yu W,Zhu Y,etal.Fine mapping of a major quantitative trait locus, qFLL6.2,controlling flag leaf length and yield traits in rice(OryzasativaL.) [J].Euphytica,2012,184(1):57-64.
[17]Duan D,Zhang H.A single SNP in NRT1.1B has a major impact on nitrogen use efficiency in rice [J].ScienceChinaLifeSciences,2015,58(8):827-828.
[18]Xue D W,Chen M C,Zhou M X,etal.QTL analysis of flag leaf in barley(HordeumvulgareL.) for morphological traits and chlorophyll content [J].JournalofZhejiangUniversity,2008,9(12):938-943.
[19]Zhang B,Ye W,Ren D,etal.Genetic analysis of flag leaf size and candidate genes determination of a major QTL for flag leaf width in rice [J].Rice,2015,8(1):2-12.
[20]Bertholdsson N,Holefors A,Macaulay M,etal.QTL for chlorophyll fluorescence of barley plants grown at low oxygen concentration in hydroponics to simulate waterlogging [J].Euphytica,2015,201(3):357-365.
[21]Wang S,Wong D,Forrest K,etal.Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array [J].PlantBiotechnologyJournal,2014,12(6):787-796.
[22]Wu Q,Chen Y,Fu L,etal.QTL mapping of flag leaf traits in common wheat using an integrated high-density SSR and SNP genetic linkage map [J].Euphytica,2016,208(2):337-351.
[23]宋荷仙,李躍建,馮君成,等.小麥收獲指數和源、庫性狀的遺傳研究 [J].中國農業科學,1993,26(3):21-26.
Song H X,Li Y J,Feng J C,etal.A study on gengtics of harvest index and source-sinnk of wheat [J].ScientiaAgriculturaSinica,1993,26(3):21-26.
[24]Porebski S,Bailey L G,Baum B R.Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components [J].PlantMolecularBiologyReporter,1997,15(1):8-15.
[25]王建康.數量性狀基因的完備區間作圖方法 [J].作物學報,2009,35(2):239-245.
Wang J K.Inclusive composite interval mapping of quantitative trait genes [J].ActaAgronmicaSinica,2009,35(2):239-245.
[26]Mccouch S R,Cho Y G,Yano M,etal.Report on QTL nomenclature [J].RiceGeneticsNewsletters,1997,11(14):11-13.
[27]李慧慧,張魯燕,王建康.數量性狀基因定位研究中若干常見問題的分析與解答 [J].作物學報,2010,36(6):918-931.
Li H H,Zhang L Y,Wang J K.Analysis and answers to frequently asked questions in quantitative trait locus mapping [J].ActaAgronmicaSinica,2010,36(6):918-931.
[28]Fan X,Cui F,Zhao C,etal.QTLs for flag leaf size and their influence on yield-related traits in wheat(TriticumaestivumL.) [J].MolecularBreeding,2015,35(1):24.
[29]陳建省,陳廣鳳,李青芳,等.利用基因芯片技術進行小麥遺傳圖譜構建及粒重QTL分析 [J].中國農業科學,2014,47(24):4769-4779.
Chen J S,Chen G F,Li Q F,etal.Construction of genetic map using genotyping chips and QTL analysis of grain weight [J].ScientiaAgriculturaSinica,2014,47(24):4769-4779.
收稿日期:2016-01-17修回日期:2016-03-07
基金項目:國家重點基礎研究計劃項目(2014CB138100);陜西省自然科學基金項目(2015JM3094);陜西省重點科技創新團隊項目(2014KCT-25)
通訊作者:馮 毅(E-mail:fengyi1455@126.com);孫道杰(E-mail:chinawheat@163.com)
中圖分類號:S512.1;S330
文獻標識碼:A
文章編號:1009-1041(2016)06-0689-10
QTL Mapping of Flag Leaf Traits Using an Integrated High-density 90K Genotyping Chip
LIAN Junfang,ZHANG Deqiang,WU Bingjin,SONG Xiaopeng,MA Wenjie,ZHOU Limin,Feng Yi,SUN Daojie
(College of Agronomy,Northwest A&F University,Yangling,Shaanxi 712100,China)
Abstract:In order to carry out the quantitative trait loci(QTLs) of wheat flag leaf traits,in this study,QTLs for flag leaf traits were mapped using an available high-density 90K wheat SNP and SSR genetic linkage map developed from a recombinant inbred line(RIL) population of Zhou 8425B × Xiaoyan 81.A genetic map covering 21 wheat chromosomes was constructed,which contains 6 949 polymorphism markers(6 910 SNP and 36 SSR),with a total genetic distance of 4 839.9 cM and an average interval distance of 0.7 cM.A,B and D chromosomes posseed 2 085,4 677 and 187 polymorphism markers,accounting for the proportion of 30.0%,67.3% and 2.7%,respectively.And the average interval distance of the markers were 1.0,0.6 and 0.8 cM,respectively.Using phenotypic data at two locations(Yangling and Anyang) in two years(2013-2014 and 2014-2013 growing seasons),22 QTLs for flag leaf length(FLL),flag leaf width(FLW) and flag leaf area(FLA) were detected by inclusive composite interval mapping(ICIM)(LOD ≥ 2.5). Among those QTLs,ten QTLs for FLL were mapped on chromosome 2A,3B,4B,5A,6B and 7B with phenotypic variations ranging from 7.900% to 24.098%.A stable and major QTL associated with FLL, Qfll2A-1,was detected in two environments,while the other nine QTLs can be detected in only one environment.Four QTLs for FLW were found on chromosome 2A,3A and 5B with phenotypic variations from 9.080% to 16.540%. A stable and major QTL associated with FLW, Qflw2A-1,was detected in three environments with phenotypic variations ranging from 12.483% to 16.540%.Eight QTLs for FLA were mapped on chromosome 2A,3B,4B,5A,6B and 7A with phenotypic variations ranging from 9.310% to 30.498% and three of them were anchored on 5A chromosome.Three QTL enrichment sections of 2A,5A and 6B chromosomes were identified.
Key words:Wheat; RILs; Flag leaf traits; SNP; QTL
網絡出版時間:2016-05-30
網絡出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160530.1535.004.html
第一作者E-mail:18829785748@163.com
