高寒草甸退化對土壤昆蟲多樣性的影響
2016-07-21 01:25:40高艷美吳鵬飛
生態學報 2016年8期
高艷美,吳鵬飛
西南民族大學生命科學與技術學院,成都 610041
?
高寒草甸退化對土壤昆蟲多樣性的影響
高艷美,吳鵬飛*
西南民族大學生命科學與技術學院,成都610041
摘要:土壤昆蟲是陸地生態系統的重要組成部分,在物質循環和能量轉化過程中起著重要的作用。為了查明高寒草甸生態系統退化對土壤昆蟲群落的影響,于2011年的4、5、7和10月份別對青藏東緣的若爾蓋高寒草甸的沼澤草甸、草原草甸、中度退化草甸和重度退化草甸的土壤昆蟲群落進行了調查。共捕獲土壤昆蟲4172只,隸屬于8目35科,共46類。優勢類群有尖眼蕈蚊科幼蟲(Sciaridae larvae)、搖蚊科幼蟲(Chironomidae larvae)和象甲科幼蟲(Curculionidae larvae),其中尖眼蕈蚊科幼蟲為各退化階段的共同優勢類群。重度退化草甸的土壤昆蟲密度和多樣性指數均顯著低于其它退化階段(P<0.01)。各退化階段間的Sorenson相似性和Morisita-Horn相似性指數變化趨勢表明退化對土壤昆蟲的類群組和優勢類群的個體數量影響較大。而土壤昆蟲的群落密度和多樣性指數的季節動態在不同退化階段間也存在差異。此外,高寒草甸的退化還可影響昆蟲群落優勢類群的時空分布,但不同類群間存在差異。相關分析結果表明土壤昆蟲多樣性與土壤pH值呈顯著負相關(P<0.01),與地下生物量和磷含量呈顯著正相關(P<0.01),而密度僅與pH值呈顯著負相關(P<0.01)。研究結果表明高寒草甸退化可通過改變植物群落及土壤等環境因子影響土壤昆蟲群落組成和多樣性的空間分布和季節動態。
關鍵詞:土壤昆蟲;多樣性;高寒草甸;退化;若爾蓋
昆蟲種類多,數量巨大,適應性較強,是生態系統的重要組成部分[1- 2],在草地生態系統的物質循環和能量流動過程中起著重要的調控作用。與脊椎動物相比,昆蟲對氣候、植被、土壤等環境因子的變化具有高度敏感性,能在一定程度上能指示環境變化[3- 4],在生態恢復和環境監測方面,昆蟲作為生物指標之一正在得到更多地關注和應用[5- 6]。
青藏高原上的高寒草地面積約為1.2×108hm2,占全國草地總面積的1/3[7]。位于青藏東緣的若爾蓋草地總面積為7.08×l04km2,是我國重要的畜牧業基地之一,也是黃河的發源地和我國重要的生態屏障[8]。自20世紀70年代以來,在全球變化和放牧干擾的綜合作用下,若爾蓋草地也呈現出明顯的退化趨勢[9],沙化草地面積也逐漸擴大[10]。高寒草地退化后,植物群落[11]、土壤理化性質[12]都發生明顯變化。昆蟲群落與賴以生存的植物群落之間有著極其密切的聯系,植物群落的組成及變化決定著昆蟲群落的特征[13- 14]。
目前關于高寒草甸生態系統昆蟲的研究非常少[15],因此對高寒草甸退化過程中土壤昆蟲群落組成及變化趨勢等基本情況都不清楚,查明這些問題對揭示高寒草甸生態系統結構和功能的變化趨勢具有重要的科學意義。本文以若爾蓋高寒草甸不同退化階段土壤昆蟲為研究對象,目的是查明:高寒草甸不同退化階段土壤昆蟲群落組成特征和高寒草甸退化對土壤昆蟲群落多樣性的影響。研究結果不僅可以提供高寒草甸生態系統土壤昆蟲多樣性方面的基礎資料,還能為高寒草甸生態系統的病蟲害防治提供科學依據。
1研究區概況
若爾蓋縣位于青藏高原東部邊緣,地理坐標東經102°08′—103°39′、北緯 32°56′—34°19′,地處四川省阿壩藏族羌族自治州北部。海拔在2400—4500 m之間,平均海拔3500 m。該區屬于大陸性季風高原型氣候,具有寒溫帶氣候特征。日照充足,晝夜溫差大,年日照2507 h。年平均氣溫0.612 ℃,極端最高溫度24.6 ℃,極端最低溫度-29.5 ℃。最大凍土深72 cm,無絕對無霜期。年平均降水量約為600—750 mm,蒸發量1212.7 mm,相對濕度68%。區域內主要地貌類型有高原、丘陵、山原和高山峽谷,主要土壤類型有草甸土、沼澤土和泥炭土。
2研究方法
2.1樣地設置
在若爾蓋縣境內根據植物的蓋度、群落組成和土壤類型選取沼澤草甸、草原草甸、中度退化草甸和重度退化草甸4種典型生境作為固定樣地,樣地面積為50 m×50 m。各個樣地的土壤類型分別為沼澤土、草甸土、草甸土和風沙土。沼澤草甸的主要植物種類有:垂穗披堿草(Elymusnutans)、高原毛茛(Ranunculustanguticus)、火絨草(Leontopodiumleontopodioides)、珠芽蓼(Polygonumviviparum)、蒲公英(Taraxacummaurocarpum)、紫菀(Astertataricus)、毛莨狀金蓮花(Trolliusranunculoides)、報春花(Primulamalacoides)、老鸛草(Herbaerodii)等,高度10—30 cm,蓋度90%以上。草原草甸的主要植物種類有:藏羊茅(Festucawallichanica)、四川嵩草(Kobresiasetchwanensis)、高原毛茛、葛縷子(Carumcarvi)、狼毒(Stellerachamaejasme)、乳白香青(Anaphalislactea)、條葉銀蓮花(Anemonetrullifolia)、矮金蓮花(Trolliusfarreri)、垂穗披堿草、草地早熟禾(Poapratensis)、棘豆(OxytropisKansuensis)等,高度20—40 cm,蓋度90%以上。中度退化草甸主要植物種類有:鵝絨委陵菜(Potentillaansrina)、嵩草(Kobresiabellardii)、黃帚橐吾(Ligu1ariavirgaurea)、蒲公英、高原毛莨、棘豆等,高度0—5 cm,蓋度50%—70%以上。重度退化草甸的主要植物種類有:垂穗披堿草,葛縷子,高度0—20 cm,蓋度10%以下[15]。不同退化階段的植物種類、地上生物量、地下生物量、土壤容重、pH、含水量、有機質、全氮、全磷、全鉀、速效氮、有效磷和速效鉀共13個環境因子的基本情況見文獻[15]。
2.2土壤昆蟲調查及鑒定
2011年的4、5、7月和10月,對若爾蓋高寒草甸進行4次采樣,共采集96個土樣(4個階段×4次調查×6個重復)。在每種樣地內隨機選取6個50 cm×50 cm的樣方,每個樣方之間距離為5 m以上,采取0—15 cm高度范圍內的土壤。體長在2 mm以上的土壤昆蟲采用手揀法獲取,并用75%酒精溶液保存。另取土樣1份,在實驗室內用干漏斗法(Tullgren法)分離橫截面直徑為2 mm以下的昆蟲,溫度控制在37 ℃左右,分離時間為48 h。
土壤昆蟲的鑒定,把收集到的土壤昆蟲在實驗室內依據《中國土壤動物檢索圖鑒》[16]、《昆蟲分類檢索》[17]和《幼蟲分類學》[18]等在體式鏡(Olympus SZX16)和光學顯微鏡(Leica DM4000B)下對土壤昆蟲進行鑒定,除部分鑒定到屬外其余均鑒定到科,統計個體數量,并換算成個/m2。
2.3數據處理
(1)多度劃分
個體數占總捕獲量10%以上的為優勢類群,個體數占總捕獲量1%—10%之間的為常見類群,個體數占總捕獲量的1%以下的為稀有類群[19]。
(2)群落多樣性特征
采用豐富度、Shannon-Wiener多樣性指數H′和Simpson優勢度指數C來分析多樣性。其中以土壤昆蟲的類群數表示豐富度,而Shannon-Wiener多樣性指數和Simpson優勢度指數計算方法如下。
Shannon-Wiener多樣性指數[20]:
Simpson優勢度指數[21]:
式中,S為類群數,Pi為第i類群的百分比。
(3)土壤昆蟲群落相似性分析
分別采用Sorensen相似性系數和Morisita-Horn相似性系數對群落相似性進行定性和定量分析[22]。
Sorensen相似性系數:
式中,c為A和B兩種生境共有的類群數,a為A生境的總類群數,b為B生境的總類群數。該相似性指數主要反映群落組成類群的變化,即定性相似性。值在0.75—1.00之間為極相似,0.50—0.74之間為中等相似,0.25—0.49之間為中等不相似,0—0.24之間為極不相似。
Morisita-Horn 相似性系數:
式中,Na為群落A的物種數量,Nb為群落B的物種數量,ai和bi分別為A和B群落中第i個體數目。
(4)統計分析
對于服從正態分布的數據,利用單因素方差分析(One-Way ANOVA)和重復測量方差分析(Repeated measures ANOVA)分別對各退化階段季節間和不同退化階段間土壤昆蟲群落密度和多樣性的差異顯著性進行檢驗。如果差異顯著則用Tukey HSD法(方差齊性)和Tamhane′s T2(M)法(方差不齊)。對于不服從正態分布的數據,則用log(x+1)轉換,如若轉換后仍不服從正態分布的數據則利用 Kruskal-Wallis Test (H)和Mann-Whitney進行非參檢驗。土壤昆蟲群落與環境因子間的相關性用回歸分析進行檢驗。用SPSS 20.0對數據進行統計分析。
3結果與分析
3.1土壤昆蟲群落組成特征
2011年4、5、7、10月4個月份在若爾蓋高寒草甸共捕獲土壤昆蟲4172個,隸屬于8目,35科,共46類(表1)。優勢類群有尖眼蕈蚊科幼蟲(Sciaridae larvae)、搖蚊科幼蟲(Chironomidae larvae)和象甲科幼蟲(Curculionidae larvae ),分別占總捕獲量的35.84%、13.04%和13.09%;常見類群共15類,共占總捕獲量的34.24%。稀有類群共28類,僅占總捕獲量的3.80%。
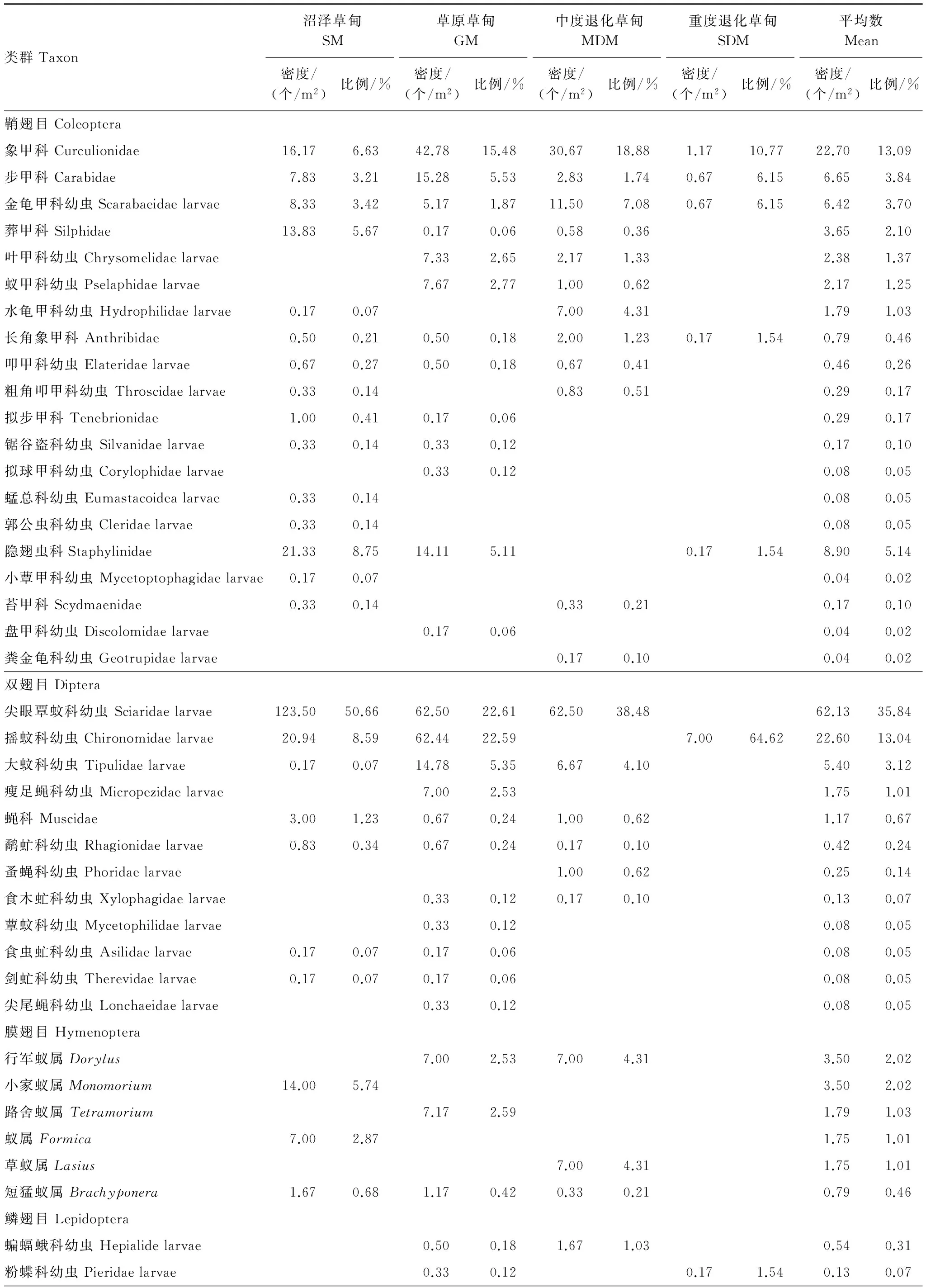
表1 若爾蓋高寒草甸不同退化階段土壤昆蟲群落組成
SM、GM、MDM、SDM分別代表沼澤草甸、草原草甸、中度退化草甸和重度退化草甸
土壤昆蟲類群數和密度高底順序均為沼澤草甸、草原草甸、中度退化草甸和重度退化草甸,類群數分別為27、31、26類和8類,密度分別為243.77、276.39、163.41、10.83 m2。不同退化階段,各類群的個體數在群落中所占比例存在一定的差異。沼澤草甸優勢種為尖眼蕈蚊科幼蟲(Sciaridae larvae),占其總個體數的50.66%,常見種共8類占其總個體數的37.37%,稀有種共18類占其總個體數的11.96%;草原草甸優勢種有搖蚊科幼蟲(Chironomidae larvae)、尖眼蕈蚊科幼蟲和象甲科幼蟲(Curculionidae larvae),占其總個體數的60.68%,常見種共有10類,占其總個體數的36.78%,稀有種共18類,占其總個體數2.53%;中度退化的優勢種有尖眼蕈蚊科幼蟲(Sciaridae larvae)和象甲科幼蟲(Curculionidae larvae ),占其總個體數的57.36%,常見種共10類,占其總個體數38.28%,稀有種共14類占其總個體數4.36%;重度退化優勢種有搖蚊科幼蟲(Chironomidae larvae )和象甲科幼蟲(Curculionidae larvae),共占其總個體數的75.38%,常見種共6類占其總個體數24.62%,沒有稀有類群。 以上分析表明草原草甸適宜較多類群的土壤昆蟲生存,群落結構復雜;重度退化僅有少數類群土壤昆蟲生存,群落結構較簡單。
3.2土壤動物群落相似性
不同退化階段間土壤昆蟲群落Morisita-Horn相似性系數變化范圍在0.10—0.86之間,而Sorenson相似性系數的變化范圍在0.27—0.57之間(表2),表明退化對群落的組成類群和優勢類群的相對個體數量影響均較大。其中,重度退化草甸與其它3個階段間的Morisita-Horn相似性系數平均值為0.29(范圍在0.10—0.58),而Sorenson相似性系數的平均值0.31(0.27—0.35之間),表明重度退化對土壤昆蟲群落的組成類群及各類群的個體數量均有較大影響。沼澤草甸、草原草甸和中度退化草甸三者間Morisita-Horn相似性系數在0.71—0.86之間,而Sorenson相似性系數在0.50—0.57之間,表明中度退化前土壤昆蟲群落的組成類群和優勢類群的相對個體數量差異較小。
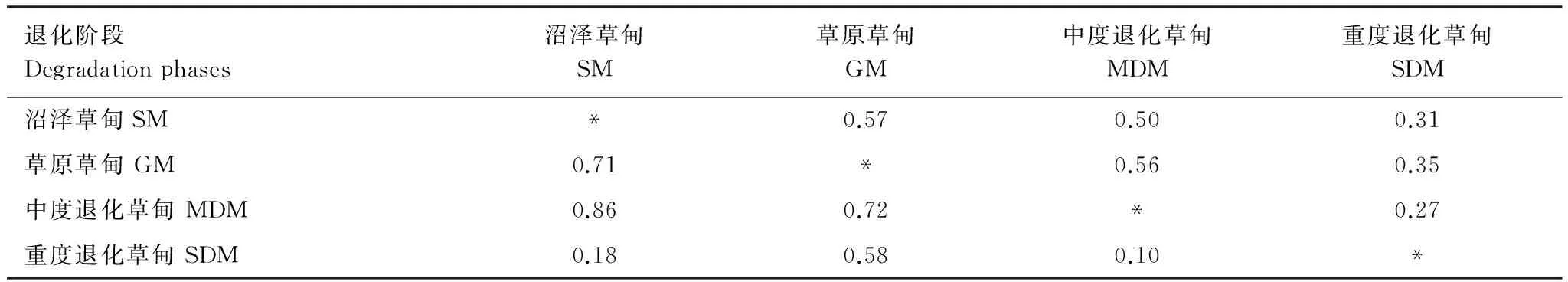
表2 各退化階段間土壤昆蟲群落相似性
對角線之上為Sorenson相似性系數,對角線之下為Morisita-Horn相似性系數
3.3土壤昆蟲的密度及多樣性動態
若爾蓋高寒草甸4種階段中的草原草甸、沼澤草甸和中度退化草甸的土壤昆蟲密度變化無顯著差異但其均顯著高于重度退化草甸(圖1和表3)。重復測量分析結果表明高寒草甸的土壤昆蟲密度有顯著的季節變化(表3),但進一步的分析表明僅有沼澤草甸的土壤昆蟲密度有顯著季節差異(圖1),其它階段的季節變化均不顯著(圖1),表明高寒草甸的退化可以影響土壤昆蟲密度的季節動態。以上結果表明高寒草甸的退化對土壤昆蟲群落的密度及其季節動態均有顯著影響。
草原草甸、沼澤草甸和中度退化草甸的土壤昆蟲的類群數(豐富度)、Shannon-Wiener多樣性指數和Simpson優勢度指數亦均無顯著差異,但均顯著高于重度退化草甸(圖1和表3)。季節變化對4個階段的土壤昆蟲類群數均有顯著的影響(圖1),而生境×季節交互作用對土壤昆蟲類群數也有顯著影響(表3)。重復測量結果表明Shannon-Wiener多樣性指數和Simpson優勢度指數有顯著季節變化(表3),但單因素方差分析表明僅有中度退化草甸和重度退化草甸中的Shannon-Wiener多樣性指數和Simpson優勢度指數的季節變化顯著(圖1)。以上結果表明高寒草甸的退化不僅可顯著降低土壤昆蟲群落多樣性,還可影響其季節變化。
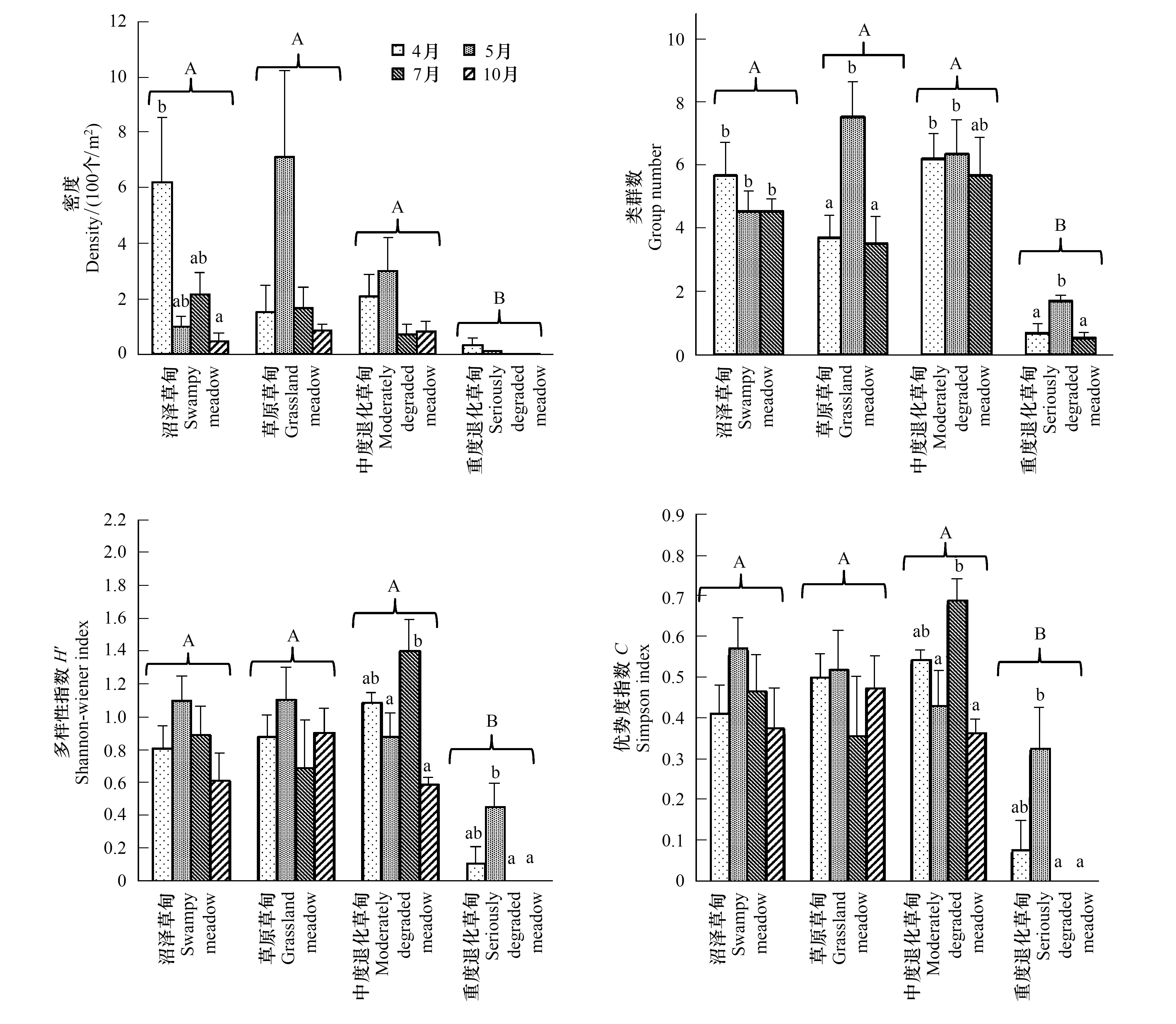
圖1 不同退化階段土壤昆蟲群落多樣性(平均值±標準誤)Fig.1 Diversity index of soil insect communities at different degradation phases (Mean±S.E.)不同大寫字母表示不同生境間差異顯著(P<0.05),不同小寫字母代表同一生境不同季節間的顯著差異(P<0.05)
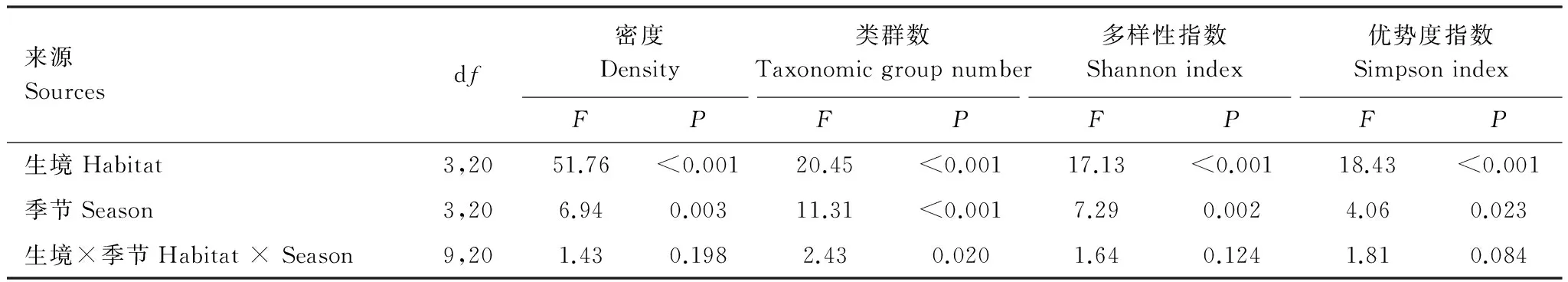
來源Sourcesdf密度Density類群數Taxonomicgroupnumber多樣性指數Shannonindex優勢度指數SimpsonindexFPFPFPFP生境Habitat3,2051.76<0.00120.45<0.00117.13<0.00118.43<0.001季節Season3,206.940.00311.31<0.0017.290.0024.060.023生境×季節Habitat×Season9,201.430.1982.430.0201.640.1241.810.084
3.4主要類群動態
若爾蓋高寒草甸土壤昆蟲主要類群為雙翅目和鞘翅目昆蟲,兩者的密度的變化順序均為草原草甸>沼澤草甸>中度退化草甸>重度退化草甸(圖2)。統計分析結果表明雙翅目昆蟲密度在不同退化階段間無顯著差異,而鞘翅目昆蟲密度在不同退化階段間存在顯著差異,且重度退化草甸鞘翅目昆蟲密度顯著低于其它退化階段(圖2和表4)。
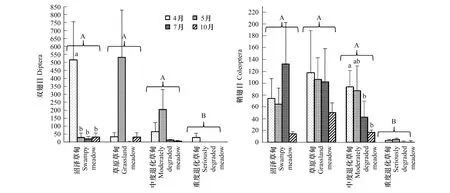
圖2 不同退化階段間雙翅目和鞘翅目的密度(平均值±標準誤)Fig.2 Densities of Diptera and Coleoptera at different degradation phases of Zoigê alpine (Mean±S.E.)
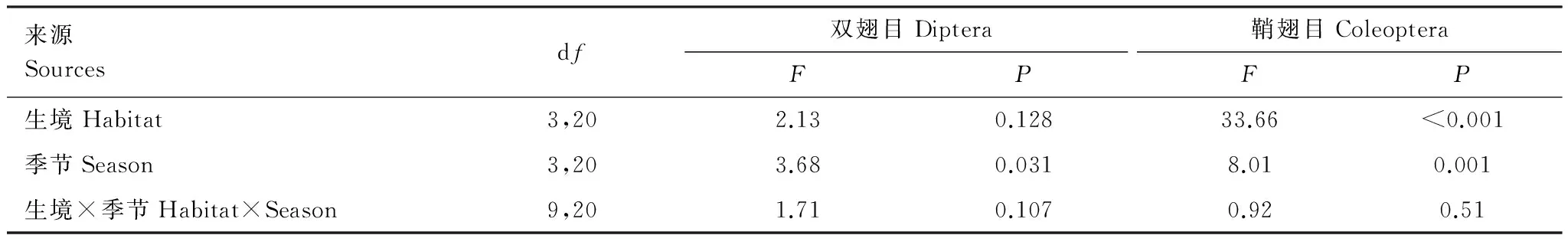
來源Sourcesdf雙翅目Diptera鞘翅目ColeopteraFPFP生境Habitat3,202.130.12833.66<0.001季節Season3,203.680.0318.010.001生境×季節Habitat×Season9,201.710.1070.920.51
此外,重復測量方差分析結果表明雙翅目和鞘翅目昆蟲密度均有顯著的季節變化(表4),但單因素方差分析表明雙翅目密度僅在沼澤草甸有顯著季節差異(F=6.08,P<0.01),鞘翅目密度僅在中度退化階段有顯著季節差異(F=3.28,P<0.05)。表明高寒草甸的退化還可影響昆蟲群落的優勢類群的空間分布及季節動態,但對不同類群的影響存在差異。
3.5土壤昆蟲群落與環境因子的關系
土壤昆蟲群落各參數與環境因子的回歸分析結果如表5。在13個環境因子中,僅有地下生物量、pH值、全磷和有效磷對土壤昆蟲有顯著影響(表5)。其中,地下生物量僅與鞘翅目密度呈顯著正相關(P<0.01);pH值與土壤昆蟲群落的密度、類群數、Shannon-Wiener指數以及雙翅目昆蟲密度呈顯著負相關(P<0.01或P<0.05);全磷含量僅與土壤昆蟲優勢度指數有著正相關性(P<0.01),而有效磷含量則與類群數、Shannon-Wiener指數及鞘翅目昆蟲密度呈顯著正相關性(P<0.01或P<0.05)。回歸分析結果表明土壤pH值的增加將會降低土壤昆蟲群落密度和多樣性,而土壤磷含量的增加則能提高其多樣性。

表5 土壤昆蟲與環境因子間的關系
括號內為非標準化系數;*表示顯著性水平為P<0.05;**表示P<0.01,雙尾檢驗
4討論
4.1高寒草甸退化對土壤昆蟲群落組成和多樣性的影響
土壤昆蟲群落組成具有鮮明的空間異質性特征。隨著若爾蓋高寒草甸退化的加劇,土壤昆蟲群落組成也隨之發生變化(表2),尤其是重度退化草甸的土壤昆蟲密度顯著低于其它草甸。已有研究表明植被和土壤理化性狀與土壤昆蟲密度有密切的關系[23],且在本文研究發現土壤昆蟲密度與pH值呈顯著性負相關。在高寒草甸退化過程中,植物群落密度、蓋度和高度明顯下降,且pH值呈顯著增加趨勢[24- 25]。因此,隨著高寒草甸退化程度的加劇,土壤昆蟲密度隨之降低。在各退化階段間,土壤昆蟲稀有類群的組成差異較大,且個體數量在群落中所占比例均在6%以下,而優勢類群的個體數量在50%以上。由于Morisita-Horn大小主要由優勢類群的數量決定,Sorenson則主要由相同類群的個數決定[25],因此4個退化階段間的Morisita-Horn相似性相對較高,而Sorenson相似性相對較低。
多樣性指數、優勢度指數以及豐富度指數(類群數)作為衡量群落多樣性的重要指標被廣泛應用[27]。若爾蓋高寒草甸的4個退化階段中,重度退化草甸土壤昆蟲的豐富度指數、Shannon指數和Simpson優勢度指數均顯著小于其它退化階段,表明重度退化可顯著降低土壤昆蟲群落多樣性。有關研究表明土壤環境是引起土壤昆蟲多樣性變化的重要因素[28],其中磷含量及pH值分別對土壤昆蟲多樣性有正、負影響[29- 30]。本文的研究結果發現土壤昆蟲的類群數和Shannon指數與土壤pH值呈顯著負相關,與磷含量呈顯著正相關,說明在高寒草甸生態系統中土壤pH值及磷含量與土壤昆蟲多樣性有間也存在密切的關系。
雙翅目和鞘翅目昆蟲密度在各種生境中占有量均較大,且雙翅目為4個退化階段的共同優勢類群,可能是因為若爾蓋是黃河流域的多雨區,其潮濕多水的環境為雙翅目昆蟲的生長發育提供了優越的條件[26]。
已有研究表明雙翅目幼蟲中的尖眼蕈蚊科幼蟲和搖蚊科幼蟲受水分影響較大[31- 32]。此外,本研究還發現雙翅目昆蟲密度僅與pH值呈顯著負相關性。隨著退化程度的加重,土壤pH值逐漸升高,這也會對雙翅目產生不利的影響。本文中數量較大的鞘翅目幼蟲,如象甲科幼蟲、金龜甲科幼蟲及葉甲科幼蟲均為植食性昆蟲[32],因此他們易受植物群落的影響。而本文的研究表明鞘翅目密度與地下生物量和有效磷含量呈顯著正相關(表5),表明鞘翅目昆蟲除了受植物群落的影響外,還受土壤環境的影響。在重度退化草甸中植被種類、蓋度及生物量等均顯著低于其他退化階段[15],因此重度退化草甸的鞘翅目昆蟲密度顯著低于其他退化階段。
4.2高寒草甸退化對土壤昆蟲季節動態的影響
在高寒草甸不同退化階段土壤昆蟲密度、類群數和多樣性均有顯著的季節變化,但不同退化階段間的季節動態不同。草原草甸和中度退化草甸土壤昆蟲的密度和類群數均以5月份最高,10月最低,與內蒙古草原土壤昆蟲的季節動態相似[33]。但沼澤草甸的土壤昆蟲密度和類群數則以4月份最高,可能是因為沼澤草甸地勢低洼、地表聚集較多水分,尤其是在5至9月間該區降雨較多[34],土壤水分處于飽和狀態,造成一些土壤昆蟲直接淹沒死亡或因土壤空氣缺氧窒息死亡[30]。在本文的研究中,重度退化草甸實際上是已經沙化了的草甸,其土壤有機質及養分含量非常低,而且長期干燥,地上基本沒有植物,無法為土壤昆蟲提供食物資源。所以該退化階段的土壤昆蟲個體數量非常少,以至于觀察不到明顯的季節變化。
致謝:西南民族大學崔麗巍、張洪芝、何先進、于曉飛、李占洋、師方等同學參與野外采樣及室內工作。
參考文獻(References):
[1]Fu S L, Zou X M, Coleman D. Highlights and perspectives of soil biology and ecology research in China. Soil Biology & Biochemistry, 2009, 41(5): 868- 876.
[2]Biaggini M, Consorti R, Dapporto L, Dellacasa M, Paggetti E, Corti C. The taxonomic level order as a possible tool for rapid assessment of arthropod diversity in agricultural landscapes. Agriculture Ecosystems & Environment, 2007, 122(2): 183- 191.
[3]陳瑜, 馬春森. 氣候變暖對昆蟲影響研究進展. 生態學報, 2010, 30(8): 2159- 2172.
[4]Nahmani J, Lavelle P, Rossi J P. Does changing the taxonomical resolution alter the value of soil macroinvertebrates as bioindicators of metal pollution?. Soil Biology & Biochemistry, 2006, 38(2): 385- 396.
[5]歐曉紅, 秦瑞豪, 王林. 滇池李居山生態恢復區昆蟲多樣性調查與分析. 云南環境科學, 2004, 23(增刊): 129- 131.
[6]王義平, 吳鴻, 徐華潮. 以昆蟲作為指示生物評估森林健康的生物學與生態學基礎. 應用生態學報, 2008, 19(7): 1625- 1630.
[7]孫鴻烈. 青藏高原的形成演化. 上海: 上海科學技術出版社, 1996.
[8]丁明軍, 張鐿鋰, 劉林山, 王兆峰. 1982—2009年青藏高原草地覆蓋度時空變化特征. 自然資源學報, 2010, 25(12): 2114- 2122.
[9]雍國瑋, 石承蒼, 邱鵬飛. 川西北高原若爾蓋草地沙化及濕地萎縮動態遙感監測. 山地學報, 2003, 21(6): 758- 762.
[10]胡光印, 董治寶, 魏振海, 逯軍峰, 顏長珍. 近30a來若爾蓋盆地沙漠化時空演變過程及成因分析. 地球科學進展, 2009, 24(8): 908- 916.
[11]王乾, 包維楷, 晏兆莉, Timo K, Alfred C, Angela M. 若爾蓋西部草甸的基本類型、特點及近幾十年來的變化. 應用與環境生物學報, 2002, 8(2): 133- 141.
[12]田應兵, 熊明標, 宋光煜. 若爾蓋高原濕地土壤的恢復演替及其水分與養分變化. 生態學雜志, 2005, 24(1): 21- 25.
[13]Stacey D A. The effects of climate change on invasive insect pests in the UK. Antenna, 1999, 23(4): 254- 256.
[14]Gray D R. The relationship between climate and outbreak characteristics of the spruce budworm in eastern Canada. Climatic Change, 2008, 87(3/4): 361- 383.
[15]吳鵬飛, 張洪芝, 崔麗巍, 鐘紅梅, 王永. 大型土壤動物群落對高寒草甸退化的響應. 土壤學報, 2013, 50(4): 786- 799.
[16]尹文英. 中國土壤動物檢索圖鑒. 北京: 科學出版社, 1998.
[17]李鴻興, 隋敬之, 周士秀, 周勤, 孫洪國. 昆蟲分類檢索. 北京: 農業出版社, 1987.
[18]鐘覺民. 幼蟲分類學. 北京: 農業出版社, 1990.
[19]劉任濤, 趙哈林, 趙學勇. 半干旱區草地土壤動物多樣性的季節變化及其與溫濕度的關系. 干旱區資源與環境, 2013, 27(1): 98- 101.
[20]Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1949.
[21]Simpson E H. Measurement of diversity. Nature, 1949, 163(4148): 688- 689.
[22]Rempala G A, Seweryn M. Methods for diversity overlap analysis in T-cell receptor populations. Journal of Mathematical Biology, 2013, 67(6/7): 1339- 1368.
[23]Koehler H. Secondary succession of soil mesofauna: A thirteen year study. Applied Soil Ecology, 1998, 9(1/3): 81- 86.
[24]吳鵬飛, 華翠, 魏雪. 若爾蓋高寒草甸不同功能群植物數量特征對退化演替的響應. 西南民族大學學報, 2014, 40(3): 1- 9.
[25]吳鵬飛, 楊大星. 若爾蓋高寒草甸退化對中小型土壤動物群落的影響. 生態學報, 2011, 31(13): 3745- 3757.
[26]顧偉, 馬玲, 丁新華, 張靜, 韓爭偉. 扎龍濕地不同生境的昆蟲多樣性. 應用生態學報, 2011, 22(9): 2405- 2412.
[27]傅必謙, 陳衛, 董曉暉, 邢忠民, 高武. 北京松山四種大型土壤動物群落組成和結構. 生態學報, 2002, 22(2): 215- 223.
[28]谷艷芳, 張莉, 丁圣彥, 欽繩武. 長期施肥對麥田大型土壤動物群落結構的影響. 生態學報, 2011, 31(17): 4900- 4906.
[29]肖能文, 劉向輝, 戈峰, 歐陽志云. 高黎貢山自然保護區大型土壤動物群落特征. 生態學報, 2009, 29(7): 3576- 3584.
[30]李娜, 張雪萍, 張利敏. 三種溫帶森林大型土壤動物群落結構的時空動態. 生態學報, 2013, 33(19): 6236- 6245.
[31]蘇蘭, 黃俊浩, 吳明, 吳鴻. 濕地植被演替中昆蟲多樣性變化研究進展. 生態學雜志, 2012, 31(6): 1577- 1584.
[32]林琳, 鄔天媛, 李景科, 張雪萍. 大慶草甸草原區大型土壤動物功能類群. 地理研究, 2013, 32(1): 41- 54.
[33]興安, 羅瑞芳, 劉新民. 內蒙古典型草原大型土壤動物群落特征. 內蒙古師范大學學報: 自然科學漢文版, 2011, 40(6): 622- 628.
[34]郭潔, 李國平. 若爾蓋氣候變化及其對濕地退化的影響. 高原氣象, 2007, 26(2): 422- 428.
Effects of alpine meadow degradation on soil insect diversity in the Qinghai-Tibetan Plateau
GAO Yanmei, WU Pengfei*
CollegeofLifeScienceandTechnology,SouthwestUniversityforNationalities,Chengdu610041,China
Abstract:Soil insects are an important component and play a key role in the material cycling and energy flow processes in grassland ecosystems. However, soil insect populations are easily influenced by changes in environmental factors. The Qinghai-Tibetan Plateau is the highest and largest plateau in the world, and is mainly dominated by alpine meadow vegetation. Alpine meadow degradation has become an important, widespread, and growing ecological problem in the Qinghai-Tibetan Plateau due to the effects of climate change and increasing overgrazing. The Zoigê wetland, which is located on the eastern edge of the Qinghai-Tibetan Plateau, has also degraded due to climate change and increased overgrazing over the last 50 years. Currently, there are four typical habitats (swamp meadow, grassland meadow, moderately degraded meadow, and seriously degraded meadow) in this area. However, the changes in the structure and diversity of the soil insect community among the four habitats are unknown. Six plots of 50 cm × 50 cm were selected in each habitat to investigate the effects of alpine meadow degradation on the soil insect community in April, May, July, and October of 2011. A total of 4,172 insect individuals were captured and classified into 8 orders, 35 families, and 46 taxonomic groups. The Sciaridae larvae, Chironomidae larvae, and Curculionidae larvae were the dominant groups in soil insect communities, and Sciaridae larvae comprised the most dominant group at all four degradation phases. The density and diversity indices of the soil insects were significantly lower in the seriously degraded meadow compared with the other degradation phases (P < 0.01). The Sorenson and Morisita-Horn similarity indices in the different degradation stages indicated that degradation of the alpine meadow had a strong effect on the taxonomic composition and dominant group abundances of soil insect communities. Moreover, the seasonal dynamics in the density and diversity of soil insect communities also differed among the different degradation stages. Degradation of the alpine meadow also changed the spatio-temporal distributions of the dominant taxonomic groups of soil insect communities. Correlation analysis showed that the diversity indices of soil insect communities were negatively correlated to soil pH (P < 0.01) and were positively correlated to below-ground biomass and phosphorus contents (P < 0.01), while the community density was only negatively correlated to soil pH (P < 0.01). Our results indicated that degradation of the alpine meadow influenced the taxonomic composition and spatio-temporal distributions of the soil insect communities by altering plant communities and soil properties.
Key Words:soil insect; diversity; alpine meadow; degradation; Zoigê
基金項目:國家自然科學基金資助項目(41371270, 40801092);四川省教育廳創新團隊資助項目(14TD0049);中央高校優秀團隊及重大孵化項目(2014NZYTD01資助和西南民族大學研究生創新型科研項目(CX2014SZ114)
收稿日期:2014- 11- 04; 網絡出版日期:2015- 05- 18
*通訊作者
Corresponding author.E-mail: wupf@swun.cn
DOI:10.5846/stxb201411042173
高艷美,吳鵬飛.高寒草甸退化對土壤昆蟲多樣性的影響.生態學報,2016,36(8):2327- 2336.
Gao Y M, Wu P F.Effects of alpine meadow degradation on soil insect diversity in the Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2016,36(8):2327- 2336.
