煙草茉莉素信號途徑相關基因
2016-06-29 13:01:38王文靜陳德鑫
中國煙草科學 2016年6期
王文靜,陳德鑫,張 潔
(中國農業科學院煙草研究所,青島 266101)
煙草茉莉素信號途徑相關基因
王文靜,陳德鑫,張 潔
(中國農業科學院煙草研究所,青島 266101)
茉莉素途徑是植物重要的信號途徑,對植物的生長發育過程和抗逆過程起重要的調控作用。有生物活性的茉莉酸衍生物JA-Ile(jasmonoyl-isoleucine)與茉莉素受體復合體SCFCOI1結合后,導致茉莉素途徑負調控因子JAZ蛋白由26S蛋白酶體(即蛋白泛素化降解途徑)降解,并釋放出茉莉素途徑轉錄激活因子MYC2,從而激活茉莉素信號途徑下游與植物生長發育相關和抗逆相關的功能基因表達和生理變化。對該信號途徑中3個重要基因COI1、JAZ、MYC2在煙草中的研究情況進行了綜述。
煙草;茉莉素信號途徑;COI1;JAZ;MYC2
茉莉酸及其揮發性衍生物茉莉酸甲酯(methyl jasmonate,MeJA)和氨基酸衍生物統稱為茉莉素類物質(jasmonates,JAs),是植物體內起整體性調控作用的植物生長調節物質。作為一種公認的新型植物激素,茉莉素不僅參與調節植物生長發育如根系生長、植物育性、塊莖形成、果實成熟以及衰老等生命活動[1-2],還在植物對生物脅迫(如昆蟲侵襲、病原菌侵染等)和非生物脅迫(如機械損傷、臭氧損傷、低溫、干旱以及鹽脅迫等)反應中起重要調控作用[3-5]。
幾十年來,通過對很多茉莉素不敏感突變體的研究,該信號途徑一些關鍵基因被克隆,結合其他生物化學手段,茉莉素信號轉導途徑逐漸清晰和完善起來。經典的茉莉酸信號轉導途徑為:當受到病原菌、機械傷害等外部刺激時,具有生物活性的茉莉酸衍生物JA-Ile(jasmonoyl-isoleucine)首先被受體復合體SCFCOI1結合識別后,原本與轉錄因子結合的蛋白質JAZs被泛素化標記并轉運至26S蛋白酶體中進行泛素化降解,釋放后的轉錄因子(以MYC2為主)得以轉錄啟動茉莉酸信號下游基因的表達,從而調控一系列由茉莉素介導的植物生長發育過程和抗性反應(圖1)[6-10]。在茉莉素信號途徑中,涉及到大量基因,如COI1、JAZ、MYC2等重要基因,這些基因大都克隆自模式植物擬南芥,其 次在煙草、水稻、番茄等植物中也有克隆。本文對來自煙草的這3個重要基因進行綜述。
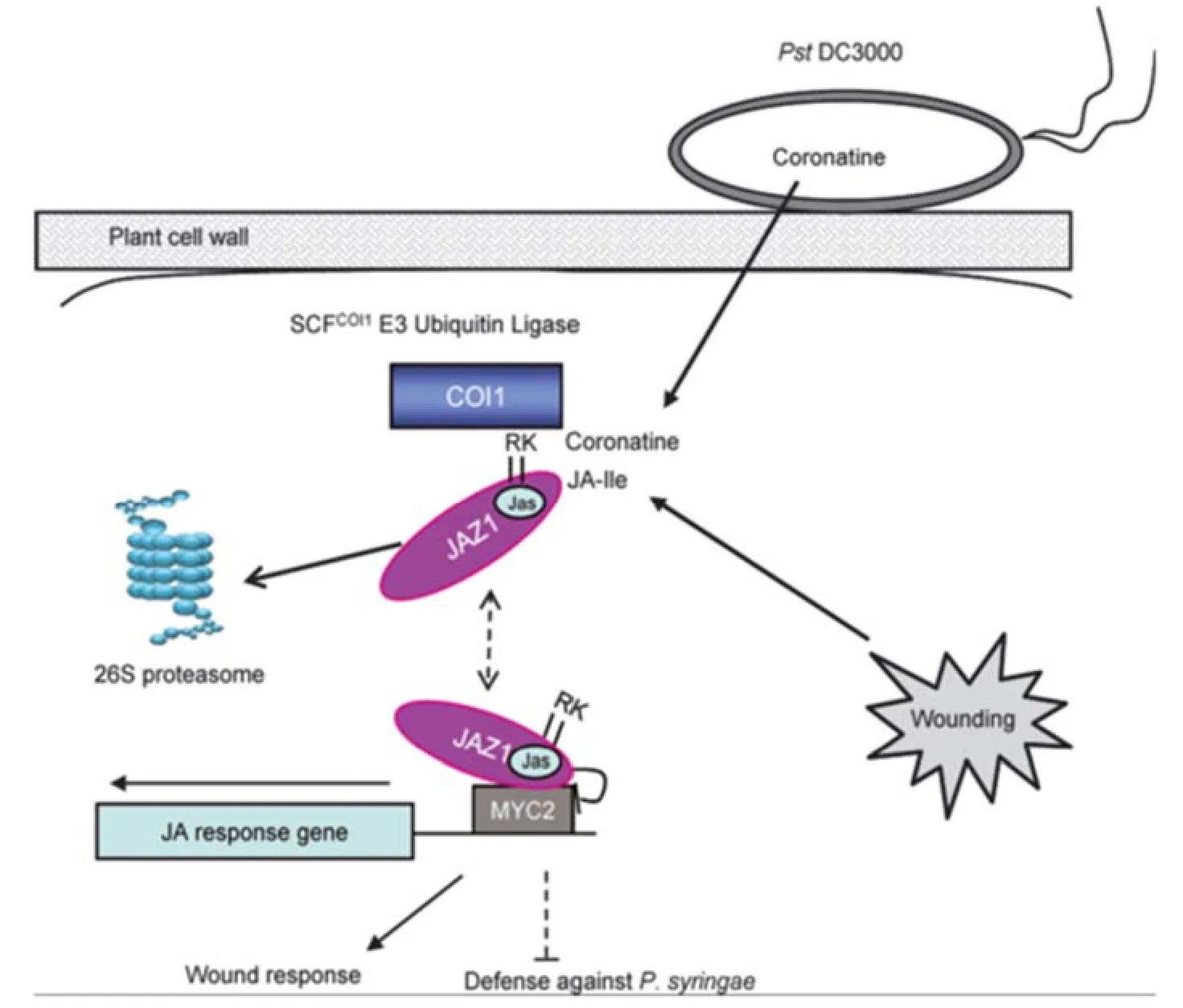
圖1 JA信號傳導途徑模式圖[10]Fig. 1 Model of JA signaling pathway[10]
1 煙草COI1基因
1994年,Feys等[11]篩選獲得了對植物毒素冠菌素(CORONATINE)不敏感的擬南芥突變體coi1(coronatine insensitivel),表現為雄性不育,易感病蟲害,而且失去了對茉莉素的正常反應,該突變體對茉莉酸信號傳導途徑的研究具有里程碑式的意義。1998年Xie等采用圖位克隆法分離出擬南芥COI1基因(CORONATINE INSENSITIVE 1)。COI1蛋白是茉莉素信號轉導途徑中的關鍵調控因子,該蛋白的缺失突變能夠導致植物喪失所有重要的茉莉素反應[12-13]。COI1基因編碼一個含有592個氨基酸的F-box蛋白,大小為66 kD,包含F-box和16個富含亮氨酸重復序列(leueine rich repeat,LRR)兩個特征結構域[12,6]。COI1的兩個特征結構域與調控茉莉酸轉導途徑的作用密切相關。F-box與Skpl、 Rbxl、Cdc53/Cullin相互結合,形成SCFCOI1復合體即E3泛素連接酶,最后被蛋白酶體降解[13-14]。
2007年,Paschold等[15]克隆了煙草的COI1基因,命名為ir-coi1,證明該基因對煙草天蛾幼蟲的防御作用。2014年,Wang等[16]克隆了煙草COI1保守區片段NtCOI1,研究了煙草COI1在花粉育性及次生代謝方面的調控機理。研究結果表明,沉默COI1煙草表現出與擬南芥coi1突變體高度類似的不育表型,即花絲縮短一半、花藥開裂延遲及花粉敗育。同時,沉默COI1煙草還喪失了花冠花色素苷合成及花蜜腺的類胡蘿卜素累積,并抑制了花冠的花色素苷合成基因及花蜜腺的類胡蘿卜素合成基因的表達水平。因類胡蘿卜素的合成與碳水化合物代謝密切相關,因此對花蜜腺的碳水化合物代謝進行了研究。試驗結果發現,沉默COI1煙草花蜜腺的淀粉累積受到顯著抑制,而且其淀粉代謝基因及與淀粉代謝相關的MYB305轉錄因子基因的表達水平都發生了改變。這些研究結果首次證明了茉莉素受體蛋白COI1對淀粉代謝的調控,深入揭示了碳水化合物代謝在茉莉素介導的次生代謝及花粉育性形成中的重要作用。
2 煙草JAZ基因
JAZ蛋白是SCFCOI1泛素連接酶復合體的一類直接底物,是茉莉素信號途徑中的一類抑制蛋白,在茉莉素信號途徑中發揮關鍵作用。JAZ蛋白為定位在細胞核中的鋅指蛋白,含2個結構域,分別是N端TIFY基序的ZIM結構域和C端與COI1互作的Jas結構域[6-7]。根據序列的相似性,目前在擬南芥中鑒定獲得了JAZ1-JAZ12共12個JAZ的蛋白[7,17-19]。
Oh等[20]報道,克隆了野生煙草中的12個 JAZ蛋白,通過反向遺傳學方法對其功能進行了研究。研究結果表明,NaJAZd的表達受昆蟲誘導,在其基因沉默后并未對煙草蟲害抗性造成明顯影響,但可導致花蕾脫落,從而影響煙草結實率。該研究結果揭示了JAZ蛋白在煙草花和果實發育中的調控作用,表明JAZ蛋白的功能多樣性。2008年,Shoji
等[21]克隆了煙草的COI1基因和JAZ基因,功能研究表明沉默COI1基因或增強JAZ負調控因子蛋白水平后,煙草對茉莉素不敏感,其煙堿合成不再應答茉莉素誘導,表明COI1基因和JAZ基因對煙草次生代謝的重要調控作用。
3 煙草MYC基因
MYC2轉錄因子是從茉莉素不敏感突變體jai1/jin1分離到的茉莉素途徑調控因子[22],屬于basic helix-loop-helix(bHLH)轉錄因子家族成員。前期對MYC2轉錄因子的大量研究證明,MYC2是植物茉莉素應答調控因子,在茉莉素途徑調控植物病害和蟲害防御應答中發揮著重要作用,并與乙烯信號傳導途徑的 ERF 轉錄因子(Ethylene-Response-Factor)拮抗性地調控下游蟲害和病害防御基因的表達[22-25]。
2012年,Zhang等[26]克隆了煙草中MYC2a、b、c 3個同源基因。編碼腐胺N-甲基轉移酶的NtPMT1a是尼古丁合成的關鍵酶。NtMYC2a/b/c編碼蛋白能特異結合NtPMT1a的啟動子核心元件G-box并激活茉莉素誘導的NtPMT1a表達。黃色熒光蛋白(YFP)熒光雙分子互補試驗表明,在無JA的情況下,NtMYC2a和NtMYC2b在細胞核內與茉莉素途徑負調控因子NtJAZ1互作。通過RNA干擾(RNAi)技術沉默NtMYC2基因后使JA誘導的NtPMT1a轉錄水平顯著降低,并抑制了其他煙堿合成基因的表達水平,其中,與合成相關的NtA662的表達水平下降了80%多。另一方面,過表達NtMYC2a和NtMYC2b未能改變NtPMT1a在有或無JA條件的表達特性。這些數據表明,NtMYC2a/b/c是多個JA誘導煙堿合成基因的關鍵表達調控因子。
煙草是我國和世界范圍內重要的經濟作物,也是重要的用于基因功能研究的模式植物,在茉莉素途徑調控植物生長發育相關研究方面具有獨特優勢。因此,對煙草茉莉素信號途徑的調控機理進行深入研究,不僅對煙草的功能基因組學研究有重要意義,同時對其他植物的基因功能研究也有重要的借鑒意義。
[1] Sembdner G, Parthier B. The biochemistry and the physiological and molecular Actions of jasmonates[J]. Annu Rev Plant Physiol Plant Mol Biol, 1993, 44(44): 569-589.
[2] Creelman R A, Mullet J E. Jasmonic acid distribution and action in plants: regulation during development and response to biotic and abiotic stress[J]. Proc Natl Acad Sci U S A, 1995, 92(10): 4114-4119.
[3] Browse J. Jasmonate: an oxylipin signal with many roles in plants[J].Vitam Horm, 2005, 72: 431-456.
[4] Jander G, Howe G. Plant interactions with arthropod herbivores: state of the field[J]. Plant Physiol, 2008, 146(3): 801-803.
[5] Browse J, Howe G A. New weapons and a rapid response against insect attack[J]. Plant Physiol, 2008, 146(3): 832-838.
[6] Chini A, Fonseca S, Fernández G, et al. The JAZ family of repressors is the missing link in jasmonate signalling[J]. Nature, 2007, 448(7154): 666-671.
[7] Thines B, Katsir L, Melotto M, et al. JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling[J]. Nature, 2007, 448(7154): 661-665.
[8] Yan J, Zhang C, Gu M, et al. The Arabidopsis CORONATINEINSENSITIVE1 protein is a jasmonate receptor[J]. Plant Cell, 2009, 21(8): 2220-2236.
[9] Sheard L B, Tan X, Mao H, et al. Jasmonate perception by inositol-phosphate-potentiated COI1-JAZ co-receptor[J]. Nature, 2010, 468(7322): 400-405.
[10] Melotto M, Mecey C, Niu Y, et al. A critical role of two positively charged amino acids in the Jas motif of Arabidopsis JAZ proteins in mediating coronatine- and jasmonoyl isoleucine-dependent interactions with the COI1 F-box protein[J]. Plant J, 2008, 55(6): 979-988.
[11] Feys B, Benedetti C E, Penfold C N, et al. Arabidopsis mutants selected for resistance to the phytotoxin coronatine are male sterile, insensitive to methyl jasmonate, and resistant to a bacterial pathogen[J]. Plant Cell, 1994, 6(5): 751-759.
[12] Xie D X, Feys B F, James S, et al. COI1: an Arabidopsis gene required for jasmonate-regulated defense and fertility[J]. Science, 1998, 280(5366): 1091-1094.
[13] Devoto A, Nieto-Rostro M, Xie D, et al. COI1 links jasmonate signalling and fertility to the SCF ubiquitinligase complex in Arabidopsis[J]. Plant J, 2002, 32(4): 457-466.
[14] Ellis C, Karafyllidis I , Wasternack C, et al. The Arabidopsis mutant cev1 links cell wall signaling to j asmonate and ethylene responses[J]. Plant cell, 2002, a) 14(7): 1557-1566.
[15] Paschold A, Halitschke R, Baldwin I T. Co(i)-ordinating defenses: NaCOI1 mediates herbivore- induced resistance in Nicotiana attenuata and reveals the role of herbivore movement in avoiding defenses[J]. Plant J, 2007, 51(1): 79-91.
[16] Wang W, Liu G, Niu H, et al. The F-box protein COI1 functions upstream of MYB305 to regulate primary carbohydrate metabolism in tobacco(Nicotiana tabacum L. cv. TN90)[J]. J Exp Bot, 2014, 65(8): 2147-2160.
[17] Browse J. Jasmonate passes muster: a rece+ptor and targets for the defense hormone[J]. Annu Rev Plant Biol, 2008, 60: 183-205.
[18] Chung H S, Howe G A. A critical role for the TIFY motif in repression of jasmonate signaling by a stabilized splice variant of the JASMONATE ZIM-domain protein JAZ10 in Arabidopsis[J]. Plant Cell, 2009, 21(1): 131-145.
[19] Kazan K, Manners J M. JAZ repressors and the orchestration of phytohormone crosstalk[J]. Trends Plant Sci, 2012, 17(1): 22-31.
[20] Oh Y, Baldwin I T, Galis I. A jasmonate ZIM-domain protein NaJAZd regulates floral jasmonic acid levels and counteracts flower abscission in Nicotiana attenuata plants[J]. PLoS One, 2013, 8(2): e57868.
[21] Shoji T, Ogawa T, Hashimoto T. Jasmonate-induced nicotine formation in tobacco is mediated by tobacco COI1 and JAZ genes[J]. Plant Cell Physiol, 2008, 49(7): 1003-1012.
[22] Lorenzo O, Chico J M, Sanchez-Serrano J J, et al. JASMONATE- INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis[J]. Plant Cell, 2004, 16(7): 1938-1950.
[23] Boter M, Ruiz-Rivero O, Abdeen A, et al. Conserved MYC transcription factors play a key role in jasmonate signaling both in tomato and Arabidopsis[J]. Genes Dev, 2004, 18(13): 1577-1591.
[24] Dombrecht B, Xue G P, Sprague S J, et al. MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis[J]. Plant Cell, 2007, 19(7): 2225-2245.
[25] Cheng Z, Sun L, Qi T, et al. The bHLH transcription factor MYC3 interacts with the Jasmonate ZIM-domain proteins to mediate jasmonate response in Arabidopsis[J]. Mol Plant, 2011, 4(2): 279-288.
[26] Zhang H B, Bokowiec M T, Rushton P J, et al. Tobacco transcription factors NtMYC2a and NtMYC2b form nuclear complexes with the NtJAZ1 repressor and regulate multiple Jasmonate-inducible steps in nicotine biosynthesis[J]. Mol Plant, 2012, 5(1): 73-84.
Jasmonate Signaling Pathway Genes in Tobacco (Nicotiana tabacum L.)
WANG Wenjing, CHEN Dexin, ZHANG Jie
(Tobacco Research Institute, Chinese Academy of Agricultural Sciences, Qingdao 266101, China)
Jasmonate signaling pathway plays an important role in the regulation of plant development and stress response. At the presence of Jasmonate, its biological active derivative JA-Ile (jasmonoyl-isoleucine) is perceived by the receptor complex SCFCOI1, which leads to the degradation negative regulator JAZ protein via 26S proteasome and then releases the transcriptional activator MYC2 to activate the expression of downstream genes and to activate physiological responses. This article will summarize the research progresses on the genes of the key Jasmonate pathway regulators COI1, JAZ and MYC2 in tobacco.
tobacco; jasmonate signaling pathway; COI1; JAZ; MYC2
S572
1007-5119(2016)06-0101-04
10.13496/j.issn.1007-5119.2016.06.018
中國農業科學院科技創新工程(ASTIP-TRIC04)
王文靜(1975-),助理研究員,主要從事病原菌與植物互作機理的研究。E-mail:wangwenjing@caas.cn
2016-09-01
2016-12-12
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
