RNA干擾endophilin A2抑制大鼠海馬神經元突觸囊泡內吞*
2016-06-06 03:34:30張吉鳳尹義臣楊萬勇譚明會郭國慶
中國病理生理雜志 2016年5期
張吉鳳, 尹義臣, 楊萬勇, 譚明會, 郭國慶
(暨南大學醫學院 1解剖學教研室, 2中西醫結合博士后流動站,廣東 廣州 510632; 3廣州市紅十字會醫院,暨南大學附屬第四醫院神經內科,廣東 廣州 510220; 4暨南大學附屬第一醫院神經內科,廣東 廣州 510632)
?
RNA干擾endophilin A2抑制大鼠海馬神經元突觸囊泡內吞*
張吉鳳1,2,尹義臣3,楊萬勇4,譚明會1,2,郭國慶1△
(暨南大學醫學院1解剖學教研室,2中西醫結合博士后流動站,廣東 廣州 510632;3廣州市紅十字會醫院,暨南大學附屬第四醫院神經內科,廣東 廣州 510220;4暨南大學附屬第一醫院神經內科,廣東 廣州 510632)
[摘要]目的: 探討內吞蛋白(endophilin)A2對大鼠海馬神經元突觸囊泡內吞的影響。方法: 設計干擾endophilin A2的小分子干擾片段,通過Western blot的方法篩選出針對endophilin A2有效的特異性干擾片段;用免疫熒光的方法驗證干擾片段對神經元內源性endophilin A2的干擾效果;利用雙全細胞膜片鉗的方法,記錄不同刺激方案下的興奮性突觸后電流(excitatory postsynaptic currents,EPSCs)的情況,以明確endophilin A2對突觸囊泡循環的影響。結果: (1)免疫熒光結果顯示endophilin A2的干擾片段能顯著減少海馬神經元內源性endophilin A2的表達(P<0.05);(2)雙全細胞膜片鉗結果顯示,0.1 Hz的低頻刺激下,干擾endophilin A2表達的神經元EPSCs大小與陰性對照組沒有明顯差異;而無論是單串高頻刺激下還是多串高頻刺激下,干擾endophilin A2表達的神經元標準化EPSCs的下降程度都明顯增強(P<0.05)。結論: (1)成功篩選出特異性干擾endophilin A2的有效干擾片段;(2)干擾endophilin A2的表達不影響海馬神經元突觸囊泡胞吐;(3)干擾endophilin A2的表達可抑制大鼠海馬神經元突觸囊泡內吞。
[關鍵詞]內吞蛋白 A2; 突觸囊泡; 內吞; RNA干擾
網格蛋白(clathrin)介導的細胞內吞作用是真核細胞內在化營養、抗原、生長因子、受體和囊泡的重要機制之一[1],尤其在神經元突觸前膜突觸囊泡的內吞以及突觸后膜受體的內吞中都發揮著重要的作用[2]。內吞蛋白(endophilin) A是網格蛋白依賴型內吞中的重要成員,其3個亞型endophilin A1、endophilin A2和endophilin A3都存在于腦組織中,且在突觸前后膜均豐富表達[3-4]。結構上endophilin A蛋白家族成員之間具有很高的同源性,擁有共同的結構特點,均包含N端的BAR結構域、中間的可變區及C端的SH3結構域,這些結構域是endophilin A蛋白參與蛋白質相互作用,執行生理功能的基礎[5]。目前endophilin A1被證實在突觸囊泡內吞和膜表面受體內吞中發揮著正性調節的作用[6-7],而結構相似的endophilin A3則被證實在受體內吞中發揮著負性調節的作用[8]。我們關于endophilin A另外一個亞型endophilin A2的研究發現,其能與N型鈣離子通道鈣離子依賴性結合來調節突觸囊泡的內吞[9]。為了進一步明確endophilin A2在內吞中的作用,本實驗利用小分子RNA干擾原代培養大鼠海馬神經元內源性endophilin A2的表達,利用雙全細胞膜片鉗檢測突觸傳遞的變化,來進一步探討endophilin A2在突觸囊泡內吞中的作用。
材料和方法
1動物
SPF級新生1 d內的SD大鼠,購于中山大學實驗動物中心,合格證編號為 SCXK (粵) 2011-0029:44008500008204、44008500008259、44008500008420、44008500008743、44008500008826、44008500008897、44008500009088和44008500009242。
2主要試劑
細胞培養用相關試劑:DMEM培養基、DMEM/F12培養基、谷氨酰胺、Neurobasal-A培養基和B27添加劑均為Gibco產品;胎牛血清購自HyClone;胰蛋白酶購自USB;多聚賴氨酸購自BD Biosciences公司。
抗體:抗endophilin A2多克隆抗體購自Santa Cruz;免疫印跡用Ⅱ抗購自ZYMED;免疫熒光用Ⅱ抗DyLight 549為Jackson產品。
免疫印跡及免疫熒光試劑:化學發光底物試劑及PVDF膜購自Pierce;封片劑購自EMS;預染蛋白Marker 購自NEB;封閉用驢血清購自Jackson;多聚甲醛購自Sigma。
電生理用試劑:NaCl、葡萄糖、KCl、CaCl2、MgCl2、HEPES、MgATP、Na2-phosphocreatine、Na2GTP、EGTA和EDTA均購自Sigma。
3主要方法
3.1siRNA序列設計利用在線軟件(www.promega.com/siRNADesigner/)針對endophilin A2亞型共設計了2條干擾片段,每個片段含19個核苷酸,分別為 si2-1(5’-GAGGTTCTATTACCTTTCT-3’)和 si2-2(5’-GCTTCGTCATCATTTAGAT-3’)二者均通過BLAST 驗證其特異性。2條干擾片段及陰性對照(negative control,NC)均由上海吉瑪制藥技術有限公司合成。
3.2HEK293細胞的培養、轉染及Western blot 檢測傳代后的HEK293細胞用含10%胎牛血清的DMEM培養基接種,置37 ℃、5%CO2培養箱內培養。待細胞達到70%~80%融合度時,將100 pmol siRNA 或 NC 與2 μg 的endophilin A2-pEGFPC1表達載體共轉染HEK293細胞。細胞轉染采用磷酸鈣的方法,首先轉染前2 h換成無血清培養基饑餓細胞,然后制備磷酸鈣-DNA沉淀,方法是將等量的質粒氯化鈣混合液逐滴加入到用移液管一直吹打著的等量的2×HEPSE緩沖液中,室溫避光靜置15 min,以形成細小的磷酸鈣-DNA沉淀,將沉淀溶液逐滴均勻加入無血清的培養基中,輕輕晃動,使DNA沉淀均勻分布在細胞培養皿中。37°C培養箱孵育4 h后,洗去沉淀,換含5%胎牛血清的DMEM培養基繼續培養,24~48 h后,用Suspension Buffer收集細胞。取出100 μL細胞裂解液用BCA法進行定量。 利用Western blot對蛋白表達進行檢測,endophilin A2 Ⅰ抗1∶1 000稀釋,Ⅱ抗1∶10 000稀釋。
3.3原代海馬神經元的培養和轉染新生1 d內的SPF級SD大鼠,去除顱骨和大腦皮質,取海馬并用解剖液清洗剪成小塊,入15 mL離心管中,加0.125%的胰酶37℃、5% CO2中消化10 min。終止消化,用拋光的玻璃吸管吹打10次,800 r/min離心5 min,棄上清,用含10%胎牛血清DMEM/F12培養液重懸細胞,400目篩網過濾,調整細胞密度并接種到PDL包被過的載玻片上,24 h后全量換2% B27的Neurobasal-A無血清培養液,以后每隔3 d半量換液。待神經元生長到8~9 d, 用磷酸鈣方法進行轉染,每孔加入1 μg GFP及30 pmol siRNAs或者NC,待48~72 h后用于后續實驗。所有實驗過程符合暨南大學醫學院實驗動物倫理規定的要求。
3.4endophilin A2的免疫熒光染色神經元轉染48~72 h后用4%PFA固定,洗后用含3%驢血清的TBS封閉1 h,用endophilin A2 Ⅰ抗按1∶100稀釋后過夜孵育,donkey anti-goat IgG (H+L) Dylight 549Ⅱ抗按1∶500稀釋孵育神經元1 h,最后用Fluoro Gel II with DAPI封片。用LSM710 63×油鏡進行拍照,GFP和endophilin A2熒光的采集通道分別為488 nm和543 nm,分辨率為1 024×1 024。
3.5電生理記錄所用電極為10 cm長,外徑為1.5 mm,內徑為0.86 mm的玻璃毛細管,在P-97水平拉制儀上用四步法拉制記錄用的微電極。尖端直徑一般為2.5~3.0 μm,入液阻抗3~5 MΩ。取轉染48~72 h后的神經元進行實驗。把培養液換成標準細胞外液后,置于OLYMPUS IX71倒置相差顯微鏡載物臺上。在400倍視野下找到胞體飽滿均勻,邊界清楚,突起較長,距離合適的2個神經元。所用放大器為HEKA EPC 10.0,軟件為PatchMaster。所得數據用FitMaster進行整理和分析。
4統計學處理
所有數據用均數±標準誤(mean±SEM)表示,用SPSS 13.0 軟件進行t檢驗,以P<0.05為差異有統計學意義。
結果
1Endo2 siRNA可有效抑制內源性endophilin A2的表達
我們針對endophilin A2設計了2條小分子干擾片段,為了篩選出有效干擾片段,我們將endophilin A2過表達載體與其對應的干擾片段共轉染HEK293細胞, 用Western blot的方法來檢測它們的干擾效果。Endophilin A2的2條干擾片段中,si2-1并沒有有效抑制endophilin A2的表達,而si2-2明顯抑制了endophilin A2的表達,于是我們選擇si2-2命名為Endo2 siRNA用于后續實驗。
為明確干擾片段是否有效干擾了培養神經元內endophilin A2的表達,我們將Endo2 siRNA與GFP共同轉染培養成熟的海馬神經元,共轉GFP的目的是用于標記轉染了干擾片段的海馬神經元,這種共轉方法已經得到了廣泛認同,共轉效率在90%以上[10]。我們將轉染48~72 h后的神經元固定后,用endophilin A2特異性的抗體對其進行免疫熒光染色。Endophilin A2在轉染了陰性對照的海馬神經元及未轉染神經元均有表達,紅色熒光均勻分布在各神經元的胞體及突起部位,而轉染了Endo2 siRNA干擾片段的海馬神經元,endophilin A2的染色并不明顯,看不見或者僅見較弱的紅色熒光分布在胞體部位,這些結果表明內源性endophilin A2的表達受到特異性siRNA的干擾。為進一步明確Endo2 siRNA對內源性endophilin A2的干擾效果,我們將轉染了陰性對照和干擾片段的細胞的平均熒光強度與其周圍未轉染神經元平均熒光強度進行了對比分析,轉染Endo2 siRNA的相對熒光強度明顯低于陰性對照組Endo2(P<0.05)。以上結果表明,endophilin A2干擾片段有效降低了培養海馬神經元內源性endophilin A2的表達,見圖1。
2干擾endophilin A2的表達不影響海馬神經元的胞吐過程
神經遞質的釋放與突觸囊泡的活動密切相關,所以通過記錄不同刺激模式下興奮性突觸后電流(excitatory postsynaptic currents,EPSCs)的大小,可以分析突觸囊泡胞吐和內吞的情況。為檢測突觸囊泡活動的情況,我們首先建立了雙全細胞膜片鉗的方法。圖2A顯示的是熒光相差顯微鏡下存在突觸聯系的2個神經元,其上為與2個神經元胞體形成緊密封接的玻璃電極,帶有綠色熒光的神經元是轉染了干擾片段或者陰性對照的細胞,作為突觸前細胞,未轉染的神經元作為突觸后,當給予突觸前神經元200~300 pA、 2 ms的刺激時,能夠引起突觸前神經元爆發一個動作電位,從而能夠在突觸后神經元記錄到EPSCs。囊泡內吞緊隨胞吐之后發生,我們首先要明確endophilin A2的干擾是否影響了胞吐過程,當給予突觸前細胞0.1 Hz的低頻刺激時,我們發現轉染NC組的神經元所激發的EPSCs幅度為(433.289±128.131) pA,而轉染Endo2 siRNA干擾片段的神經元所激發的EPSCs幅度為(416.886±113.522) pA,二者所激發的EPSCs強度沒有差異,這些結果表明干擾endophilin A2的表達并沒有影響到胞吐的過程,見圖2。
3干擾endophilin A2的表達抑制了海馬神經元突觸囊泡的內吞
高頻刺激可以使待釋放囊泡池(readily relea-sable pool,RRP)的突觸囊泡由于耗竭而產生短時程突觸抑制(short-term synaptic depression,STD),突觸抑制的程度與突觸囊泡回收的速率相關,通過比較突觸抑制的程度可以了解內吞是否受到影響。為檢測endophilin A2干擾后是否影響了突觸囊泡的內吞,我們給予了突觸前神經元5 Hz、20 s即100個高頻刺激,并記錄下這100個刺激產生的EPSCs的大小,我們將記錄到的100個EPSCs對第1個EPSCs進行標準化處理,可見轉染Endo2 siRNA的神經元短時程突觸抑制的程度明顯大于陰性對照組。同時我們對每組第81~100個的EPSCs進行平均化,結果顯示Endo2 siRNA組的平均的EPSCs的大小為 0.271±0.026,明顯低于NC組平均的EPSCs大小(0.577±0.032)(P<0.05)。以上結果表明,干擾endophilin A2的表達明顯影響突觸囊泡的內吞進程,見圖3。
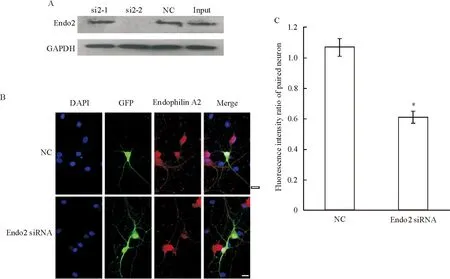
Figure 1.Screening and verification of the efficacy of endophilin A2 siRNAs.A: HEK293 cells were co-transfected with endophilin A2-pEGFPC1 plasmids and endophilin A2 siRNAs or NC, and 48 h later, endophilin A2 and GAPDH as a control in the cell lysates were probed by the designated antibodies; B:images of neurons co-transfected with GFP (green) and NC or Endo2 siRNA (scale bar=20 μm); C: fluorescence intensity of endophilin A2 in the cytoplasm transfected with Endo2 siRNA or NC was normalized to that of a paired untransfected neuron.Mean±SEM.n=10.*P<0.05vsNC.
圖1Endophilin A2 siRNAs有效片段的篩選及干擾效果驗證
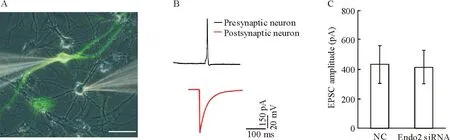
Figure 2.The impact of endophilin A2 isoform knockdown on neurotransmitter release. A: image of a pair of neurons obtained from dual patch recording transfected with siRNA or NC (green) and non-transfected neuron (gray). Scale bar=50 μm. B: representative traces from pairs of neurons. Presynaptic neuron exhibited an action potential (AP) at 2 ms, 200~300 pA current injection (black line); excitatory postsynaptic current (EPSC) was evoked in postsynaptic neuron by single AP of presynaptic neuron (red line). C: EPSC amplitude evoked in postsynaptic neuron by single AP of presynaptic neuron transfected with NC and Endo2 siRNA. Mean±SEM.n=9~12.
圖2干擾endophilin A2的表達對神經元遞質釋放的影響
如果突觸囊泡的內吞受損,那么待釋放囊泡池的再填充也將受到影響。為進一步明確endophilin A2在突觸囊泡內吞中的作用,我們采取了多串高頻刺激方案。 首先記錄在0.1 Hz刺激3 min時間內,突觸后神經元產生的平均EPSCs的大小,緊接著給予5 Hz、20 s即100個高頻刺激,然后再記錄0.1 Hz刺激3 min時間內,突觸后神經元產生的平均EPSCs,如此反復,記錄到4個高頻串刺激后的平均EPSCs,4個高頻串刺激后的平均EPSCs分別都對第1個高頻串刺激前的平均EPSCs進行均一化,通過比較各組高頻刺激后EPSCs的大小來判斷突觸囊泡的再填充情況。 多串高頻刺激方案的結果顯示,在每個高頻刺激后記錄到的EPSCs都呈現出穩定的下降趨勢, Endo2 siRNA組從第1個串刺激到第4個串刺激的EPSCs都比NC組降低得更為明顯(P<0.05)。串刺激后明顯降低的EPSCs表明, Endo2 siRNA組神經元囊泡待釋放池的囊泡再填充過程受到了阻抑,見圖4。以上這些結果表明干擾endophilin A2能明顯抑制大鼠海馬神經元突觸囊泡的內吞過程。
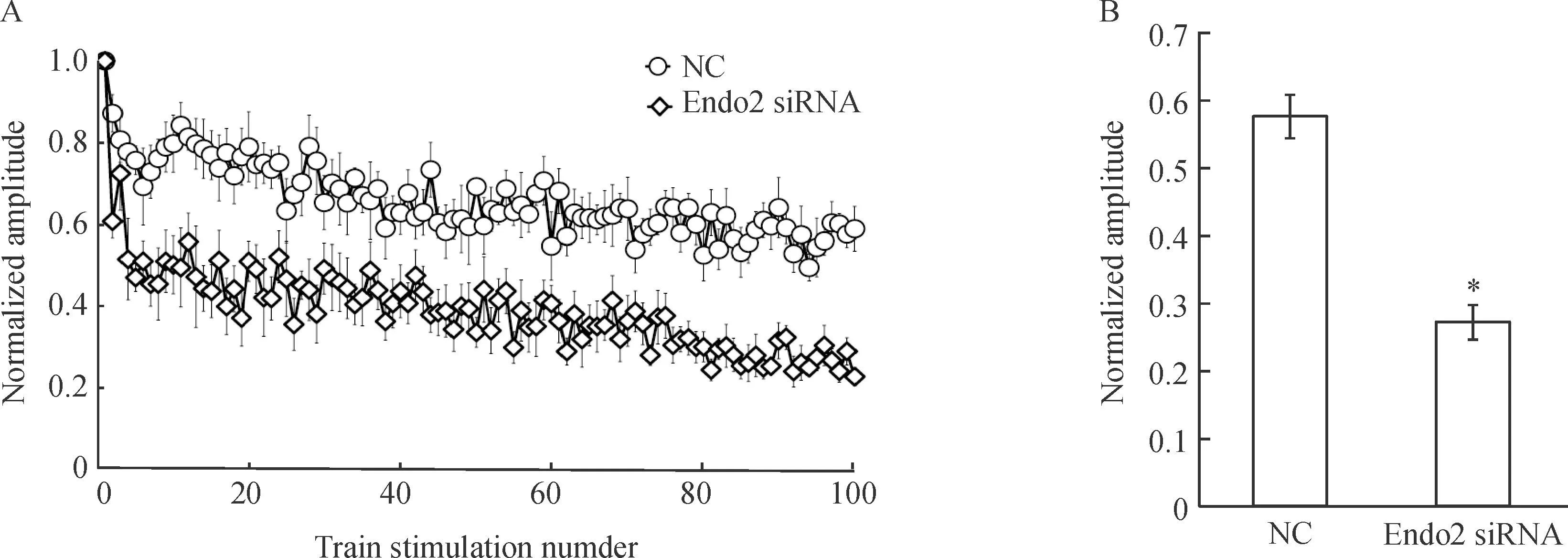
Figure 3.The impact of endophilin A2 isoform knockdown on short-term synaptic depression (STD). A:the changes of STD in postsynaptic neurons evoked by 100 pulses at 5 Hz from presynaptic neurons transfected with NC and Endo2 siRNA; B: the average normalized amplitude of the last 20 points from Endo2 siRNA compared with NC.Mean±SEM.n=7.*P<0.05vsNC.
圖3干擾endophilin A2對短時程突觸抑制的影響
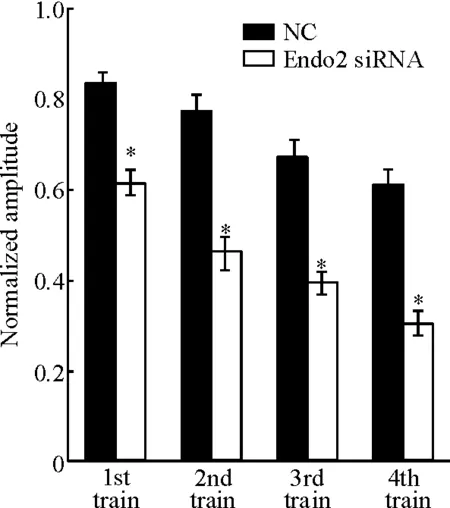
Figure 4.The impact of endophilin A2 isoform knockdown on the replenishment of the readily releasable pool(RRP). Normalized amplitude of EPSCs evoked by multiple train stimulations in neurons transfected with NC or Endo2 siRNA. Mean±SEM.n=11~13.*P<0.05vsNC.
圖4干擾endophilin A2對待釋放囊泡池再填充影響
討論
Endophilin A蛋白家族在哺乳類動物細胞中存在3種亞型endophilin A1、A2和A3,三者在中樞神經系統都豐富表達,在亞細胞水平,它們在突觸前后膜均有分布[3-4]。Milosevic等[11]為研究endophilin A在哺乳類動物突觸囊泡內吞中的功能,將小鼠endophilin A 3個亞型的基因全部敲除掉,在基因敲除老鼠體外培養的神經元上進行電生理實驗,結果發現基因敲除小鼠體外培養神經元突觸傳遞的效能明顯減弱,在給予突觸前神經元1 Hz 30個刺激的情況下,基因敲除組相比對照組就表現出明顯的突觸抑制。我們為了明確endophilin A2亞型是否在突觸前突觸囊泡內吞中發揮作用,采用了小分子干擾手段在培養的海馬神經元內特異性干擾endophilin A2的表達,同時轉染和雙全細胞膜片鉗的有效結合,使我們能夠保證干擾掉endophilin A2亞型的細胞作為突觸前,而其周圍未轉染且與轉染細胞有突觸聯系的神經元作為突觸后。當給予突觸前神經元5 Hz 100個電刺激時,突觸后記錄到的100個EPSCs隨著刺激個數的增加而逐漸下降,表現出明顯的突觸抑制,而干擾了endophilin A2亞型表達的神經元其突觸抑制的程度比對照組明顯增強,表明干擾endophilin A2的表達抑制了突觸前突觸囊泡的內吞。Milosevic等[11]由于將突觸前后膜的所有endophilin A亞型都敲除掉了,所以在他們的研究中應用更低頻率更少的刺激個數,實驗組與對照組就表現出了明顯的差異,而在我們的研究中僅僅是干擾了突觸前endophilin A2的表達。
研究顯示, endophilin A蛋白的3個亞型具有很高的同源性,均含有N段的BAR結構域、中間的可變區和C端的SH3結構域,這2個結構域被認為是endophilin A調控內吞的分子基礎[5]。例如BAR結構域能夠引起和穩定膜的彎曲,而SH3結構域能夠與dynamin等其它內吞蛋白相互作用,在囊泡剪切和脫包被中發揮作用[5, 12]。然而目前的一些報道顯示這3個亞型的功能并不一致,科學家通過向七鰓鰻的巨型軸突顯微注射endophilin A1抗體,發現突觸囊泡膜內陷反應停滯[7]。過表達endophilin A1增加了受體介導的多巴胺D2受體的內吞,相比而言,過表達endophilin A3則阻止了多巴胺D2受體的內吞[8]。在293T細胞中過表達endophilin A2促進了表皮生長因子受體(epidermal growth factor receptor,EGFR)的內吞[13]。我們的結果顯示在神經元內干擾endophilin A2的表達,抑制了突觸囊泡的內吞。從這些研究可以看到endophilin A1和endophilin A2在內吞中發揮了正性調控的作用,而endophilin A3發揮的是負性調控的作用。有分析顯示它們的不同功能可能源于其可變區的結構不同,endophilin A1和endophilin A2的可變區含有短α螺旋,然而endophilin A3的可變區則為β折疊[8]。如此,我們是否可以認為雖然endophilin A的BAR結構域和SH3結構域是內吞中發揮重要作用的分子基礎,然而位于二者之間的可變區對于調控endophilin A是否發揮功能以及發揮怎樣的功能上具有更關鍵的作用呢?由于這些研究都是在不同細胞的實驗中得出的結論,具體這3個亞型是不是存在這樣的差異,還有待我們在同一細胞體系中用同樣的實驗方法對三者的功能加以證實。
總而言之,我們的研究證實了在培養的大鼠海馬神經元上干擾endophilin A2的表達能夠抑制突觸前膜突觸囊泡的內吞。
[參考文獻]
[1]Doherty GJ, Mcmahon HT. Mechanisms of endocytosis[J]. Annu Rev Biochem,2009,78:857-902.
[2]Jung N, Haucke V. Clathrin-mediated endocytosis at sy-napses[J]. Traffic,2007,8(9):1129-1136.
[3]Giachino C, Lantelme E, Lanzetti L, et al. A novel SH3-containing human gene family preferentially expressed in the central nervous system[J]. Genomics,1997,41(3):427-434.
[4]Chowdhury S, Shepherd JD, Okuno H, et al. Arc/Arg3.1 interacts with the endocytic machinery to regulate AMPA receptor trafficking[J]. Neuron,2006,52(3):445-459.
[5]Kjaerulff O, Brodin L, Jung A. The structure and function of endophilin proteins[J]. Cell Biochem Biophys,2011,60(3):137-154.
[6]Majumdar S, Gong EM, Di Vizio D, et al. Loss of Sh3gl2/endophilin A1 is a common event in urothelial carcinoma that promotes malignant behavior[J]. Neoplasia,2013,15(7):749-760.
[7]Ringstad N, Gad H, Low P, et al. Endophilin/SH3p4 is required for the transition from early to late stages in clathrin-mediated synaptic vesicle endocytosis[J]. Neuron,1999,24(1):143-154.
[8]Sugiura H, Iwata K, Matsuoka M, et al. Inhibitory role of endophilin 3 in receptor-mediated endocytosis[J]. J Biol Chem,2004,279(22):23343-23348.
[9]Zhang J, Fan J, Tian Q, et al. Characterization of two distinct modes of endophilin in clathrin-mediated endocytosis[J]. Cell Signal,2012,24(11):2043-2050.
[10]Song B, Lai B, Zheng Z, et al. Inhibitory phosphorylation of GSK-3 by CaMKII couples depolarization to neuronal survival[J]. J Biol Chem,2010,285(52):41122-41134.
[11]Milosevic I, Giovedi S, Lou X, et al. Recruitment of endophilin to clathrin-coated pit necks is required for efficient vesicle uncoating after fission[J]. Neuron,2011,72(4):587-601.
[12]Gortat A, San-Roman MJ, Vannier C, et al. Single point mutation in Bin/Amphiphysin/Rvs (BAR) sequence of endophilin impairs dimerization, membrane shaping, and Src homology 3 domain-mediated partnership[J]. J Biol Chem,2012,287(6):4232-4247.
[13]Lua BL, Low BC. Activation of EGF receptor endocytosis and ERK1/2 signaling by BPGAP1 requires direct interaction with EEN/endophilin II and a functional RhoGAP domain[J]. J Cell Sci,2005,118(Pt 12):2707-2721.
(責任編輯: 林白霜, 羅森)
RNA interference of endophilin A2 inhibits synaptic vesicle endocytosis in rat hippocampal neurons
ZHANG Ji-feng1, 2, YIN Yi-chen3, YANG Wan-yong4, TAN Ming-hui1,2, GUO Guo-qing1
(1Department of Anatomy,2Postdoctoral Station of Integrated Traditional and Western Medicine, School of Medicine, Jinan University, Guangzhou 510632, China;3Department of Neurology, Guangzhou Red Cross Hospital, The Fourth Affiliated Hospital of Jinan University, Guangzhou 510220, China;4Department of Neurology, The First Affiliated Hospital of Jinan University, Guangzhou 510632, China. E-mail: tgqguo@jnu.edu.cn)
[ABSTRACT]AIM: To investigate the effect of endophilin A2 on synaptic vesicle endocytosis in rat hippocampal neurons.METHODS: The siRNAs for endophilin A2 isoform were designed, and the efficacy and specificity of these siRNAs were determined by Western blot. Immunostaining was performed to verify the interference efficiency of the siRNA for endogenous endophilin A2 in the hippocampal neurons. Excitatory postsynaptic currents (EPSCs) were recorded on cultured hippocampal neurons using dual whole-cell recordings by evoking the transfected presynaptic neurons in various stimulation patterns. RESULTS: The result of immunostaining in cultured hippocampal neurons demonstrated that the siRNA of endophilin A2 inhibited the expression of endogenous endophilin A2 significantly (P<0.05). The results of dual whole-cell recordings showed that no significant difference in the EPSCs amplitude evoked between endophilin A2 knockdown neurons and control was observed when stimulated at low frequency (0.1 Hz). The amplitude of normalized EPSCs evoked in endophilin A2 knockdown neurons decreased significantly for both single-train and multiple-train stimulations (P<0.05).CONCLUSION: An effective siRNA of endophilin A2 was screened out successfully. Knockdown of endophilin A2 isoform does not affect synaptic vesicle exocytosis, but inhibits synaptic vesicle endocytosis in rat hippocampal neurons.
[KEY WORDS]Endophilin A2; Synaptic vesicle; Endocytosis; RNA interference
[文章編號]1000- 4718(2016)05- 0819- 06
[收稿日期]2015- 08- 27[修回日期] 2016- 03- 07
*[基金項目]國家自然科學基金資助項目(No.31300885; No.81571191);廣東省自然科學基金資助項目(No.S2013040013256);廣東省科技計劃(No.2013B021800036);中國博士后科學基金資助項目(No.2015M570750);2015年暨南大學第一臨床醫學院科研培育專項基金資助項目(No.2015109)
通訊作者△Tel: 020-85220251; E-mail: tgqguo@jnu.edu.cn
[中圖分類號]R338.1+3; R363
[文獻標志碼]A
doi:10.3969/j.issn.1000- 4718.2016.05.008
雜志網址: http://www.cjpp.net
