不同地理來源的甘蔗梢腐病菌營養體親和性研究
2016-05-30 07:42:03郭強崔一平徐世強張木清
熱帶作物學報 2016年10期
郭強 崔一平 徐世強 張木清

摘 要 為探討甘蔗梢腐病病原菌菌株的群體結構及種內的分化,以采自廣西、云南、福建和海南等不同地區的30個甘蔗梢腐病菌株為材料,通過初步的分子生物學鑒定,發現30個甘蔗梢腐病菌株分屬于2個不同的病原種。利用KClO3培養基誘導篩選獲得Nit突變體,其中1個菌株沒有產生Nit突變體,剩下的29個菌株共產生266個Nit突變體。營養體親和群的測定結果表明:9個菌株的VCG沒有同一性,1個菌株組成1個VCG;不同地理來源的甘蔗梢腐病菌株間互不親和,分屬于不同的VCG;同一地理來源的甘蔗梢腐病菌株間亦不親和;暗示引起甘蔗梢腐病的串珠鐮刀菌屬群體內存在豐富的VCG多樣性,在不同的栽種地區群體結構變化很大,并且營養體親和性與菌株的地理來源及寄主品種都沒有明顯的相關性。
關鍵詞 甘蔗梢腐病;串珠鐮刀菌;Nit突變體;營養體親和群
中圖分類號 S432.44 文獻標識碼 A
Abstract Sugarcane pokkah boeng disease has become an important disease in china. In order to analysis the population structure and the genetic differentiation in species of the pathogen. The 30 strains of Fusarium moniliforme were collected individually from the pokkah boeng samples in Guangxi, Yunan, Fujian and Hainan provinces in China. Based on the molecular biology analysis, 30 isolateds divide into two species. After single spore isolation, 266 nitrate-nonutilizing(Nit)mutants were got from 29 strains after these colony transplanted on potato sucrose agar medium with 1.5% KClO3(KPS); 1 strains can't produce nit mutant. Furthermore, vegetative compatibility of these strains were analyzed on MM plates. In this study, the results show that all of the 9 isolates don't have VCG with each other,or 1 strains correspond to 1 VCG. This phenomenon occur not only in the strains from different geographic origins but also in the strains from the different host varieties in the same geographic origins. All these results indicate there was abundant diversity of VCG in the population of F. moniliforme, and there was no obvious correlation between the vegetative compatibility and geographic origins or host varieties.
Key words Sugarcane Pokkah Boeng Disease; Fusarium moniliforme; Nit mutant; Vegetative compatibility groups
doi 10.3969/j.issn.1000-2561.2016.10.017
真菌營養體親和性(Vegetative compatibility)又稱體細胞親和性(Somatic compatibility),是指真菌菌株間菌絲能融合并交換細胞質或核物質的特性;根據營養體親和性劃分的群體稱為營養體親和群(Vegetative compatibility group, VCG)[1-2]。自1923年Cayley等[3]首次報道發現了植物病原真菌營養體親和與不親和現象。目前,VCG已經逐漸成為研究病原真菌遺傳變異、種群地理分布、生理小種鑒定等重要手段[4-5]。在中國,VCG方法已經廣泛應用在栗疫病、黃瓜枯萎病、向日葵黃萎病和水稻紋枯病等病原菌的分類上[6-9]。而涉及串珠鐮刀菌屬真菌的研究則較少,有關甘蔗梢腐病病原菌的營養體親和性研究尚未見報道。
甘蔗梢腐病是甘蔗上一種重要的真菌病害,對甘蔗生產造成了巨大的經濟損失[10]。近年來,甘蔗梢腐病在中國廣西、云南、福建以及海南等蔗區呈現出逐漸加重的趨勢。過去的研究發現,甘蔗梢腐病主要發生在高溫多濕的夏季,病原菌主要屬于F.verticillioides[12]。本實驗室在冬季進行田間調查時發現,冬季的甘蔗上也出現了梢腐病的危害;通過分離鑒定發現引起冬季梢腐病的病原菌屬于F. proliferatum[12]。同時發現同一病原小種在不同的甘蔗品種上的危害性也不同。出現這種現象的原因,除了品種抗感性不同外,還有可能是病原種內也存在著分化現象。為了進一步解析這種現象,同時對新發現的甘蔗梢腐病病原種F. proliferatum與F.verticillioides是否具有營養體親和性進行進一步的研究。本文期望通過甘蔗梢腐病菌株營養體親和性的研究,旨在明確同一地理位置不同甘蔗品種以及不同地理來源的甘蔗梢腐病菌的營養體親和群和病原菌群體內的VCG結構,探索甘蔗梢腐病菌的種內分化、病害流行規律,更好地了解其種群組成,為甘蔗梢腐病菌遺傳變異的機制研究提供參考,并為其綜合治理和抗病育種研究提供依據。
1 材料與方法
1.1 材料
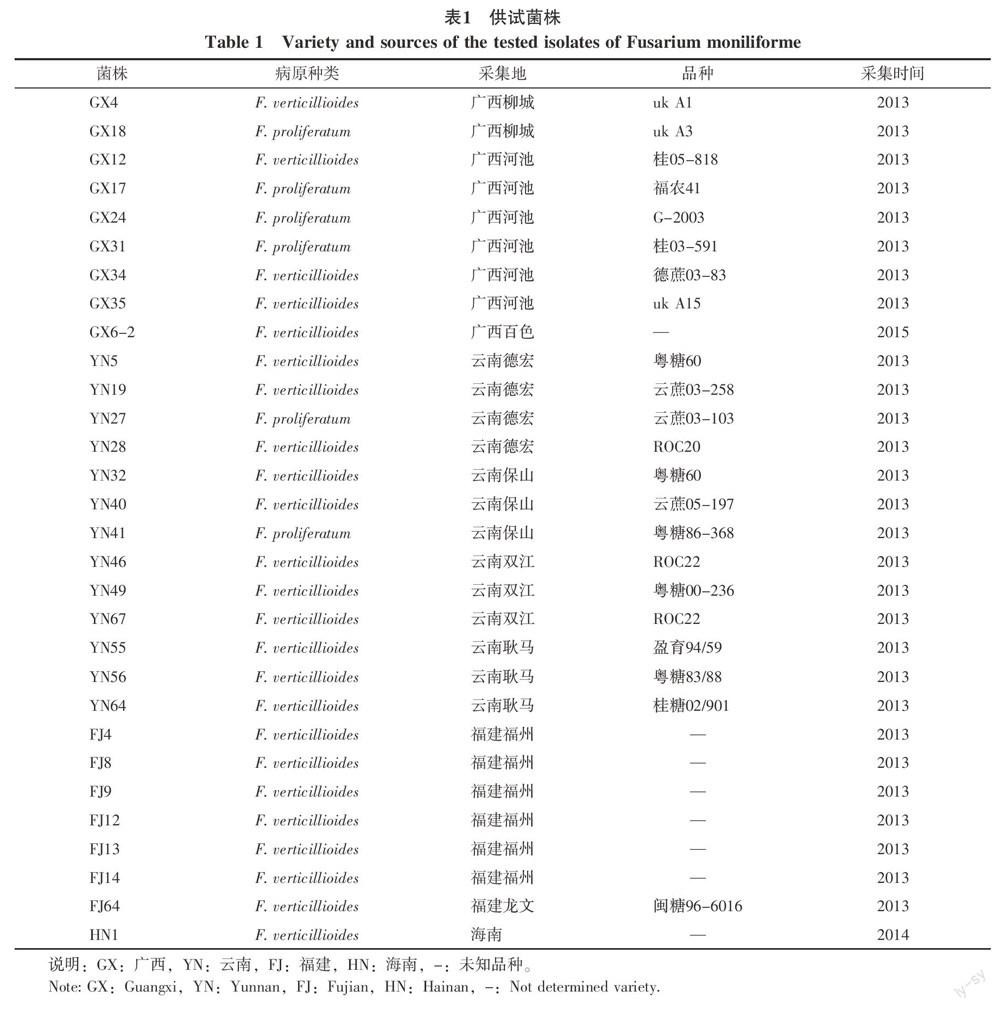
1.1.1 供試菌株 30個供試菌株(Fusarium moniliforme)于2013~2015年期間分別采集于廣西、云南、福建、海南等地,經單孢分離鑒定后獲得(表1)。
1.1.2 培養基 培養基及其組成成分見表2。
1.2 方法
1.2.1 Nit突變體誘導 將供試菌株接種于PDA平板上,于28 ℃培養箱中活化4 d。挑取活化好的菌落邊緣直徑約5 mm的菌塊,轉接于KPS(1.5%KClO3)培養基中,每個培養皿接一塊,每個菌株5個重復,于28 ℃恒溫暗培養箱中培養,觀察菌落生長情況。若形成不規則菌落(非放射狀菌落或扇形角變區),則為抗氯酸鹽突變體。
在不規則菌落的不同突變區挑取菌落邊緣的幼嫩菌絲轉接于MM培養基中,野生型菌株作為陰性對照。于28 ℃培養箱中培養4 d,觀察菌落生長情況。若產生氣生菌絲,則說明其為抗氯酸鹽突變體,而未形成硝酸鹽利用缺陷型突變體,應將其丟棄。若不產生氣生菌絲或菌落很稀薄,說明形成了硝酸鹽利用缺陷型突變體,即Nit突變體,應將其轉入MM斜面,4 ℃保存備用。
1.2.2 Nit突變體穩定性鑒定 將獲得的Nit突變體轉接于PDA培養基中,于28 ℃培養箱中培養4 d后,挑取菌落邊緣的幼嫩菌絲轉接于MM培養基中,28 ℃培養箱中培養4 d,并觀察其生長情況。若產生氣生菌絲,則說明Nit突變體不穩定,應將其丟棄。若仍不產生氣生菌絲或菌落很稀薄,則說明形成了穩定的Nit突變體,應將其轉入MM斜面,4 ℃保存備用。
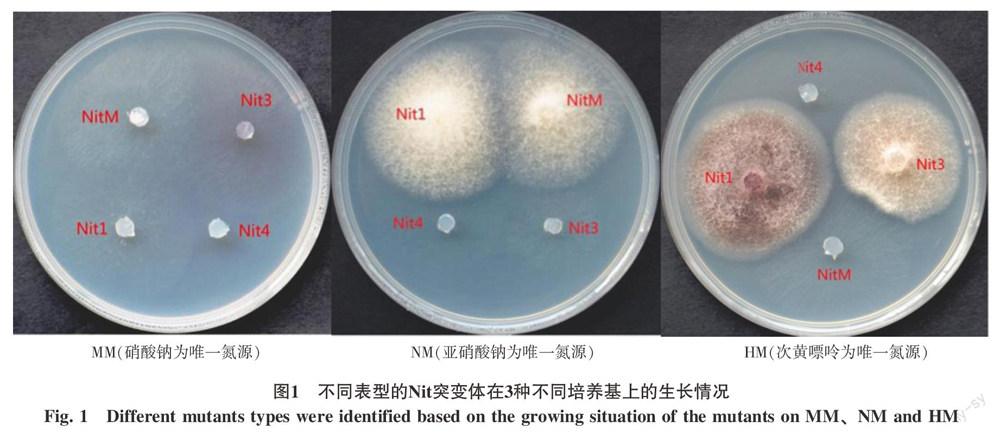
1.2.3 Nit突變體生理表現型鑒別 將穩定的Nit突變體分別轉接于MM(硝酸鈉為唯一氮源)、NM(亞硝酸鈉為唯一氮源)和HM(次黃嘌呤為唯一氮源)3種培養基中,于28 ℃培養箱中培養4 d。根據菌落生長情況將Nit突變體劃分為4種生理表型;在MM培養基中不產生氣生菌絲或菌落很稀薄,而在NM和HM培養基中均能正常生長的的突變菌株為Nit1;在MM和HM培養基中不能生長或菌落很稀薄,而在NM培養基中能正常生長的突變菌株為NitM;在MM和NM培養基中不能生長或菌落很稀薄,而在HM培養基中能正常生長的突變菌株為Nit3;在MM、NM和HM三種培養基中均不能生長或菌落很稀薄的突變菌株為Nit4(圖1)。
1.2.4 Nit突變體異核體自身親和性測定 挑取NitM突變體菌落邊緣的幼嫩菌絲直徑約5 mm的菌塊,放置于MM培養基中間,周圍等近距離放置同一菌株的Nit1和Nit3突變體,進行互補配對實驗,兩菌株間距2 cm,每平板放置4~6個菌株,于28 ℃恒溫暗培養箱中培養4~7 d。若在NitM與Nit1或Nit3突變菌株菌落交界處出現野生型生長,形成一條濃密的氣生菌絲帶,則說明該菌株具有異核體自身親和性(HSC)。反之,若菌株間沒有形成明顯的氣生菌絲帶,只有稀薄的菌絲,則為異核體自身不親和性(HSI)。
1.2.5 營養體親和群(VCG)的測定 根據異核體自身親和性鑒定結果,各菌株分別選取2~3株具有強親和力的NitM突變體,挑取其菌落邊緣的幼嫩菌絲直徑約5 mm的菌塊,放置于MM培養基中間,周圍等近距離放置不同菌株的Nit1和Nit3突變體,進行互補配對培養,兩株菌間距2 cm,每平板接4~6個菌株,于28 ℃恒溫暗培養箱中培養4~7 d。若在NitM與Nit1或Nit3突變菌株菌落交界處出現野生型生長,形成一條濃密的氣生菌絲帶,則說明這兩個菌株屬于同一營養體親和群(VCG);若菌絲交界處沒有形成氣生菌絲帶,則兩菌株不屬于同一個營養體親和群(VCG)。
2 結果與分析
2.1 甘蔗梢腐病菌Nit突變體的篩選與鑒定
甘蔗梢腐病菌在PDA培養基上活化4 d后,移入KPS培養基中誘導培養;菌株在KPS培養基中1~2 d內基本不生長,受到了嚴重的抑制;第4天開始出現菌絲的緩慢生長,第5天在菌落的一側出現旺盛的氣生菌絲生長,形成扇形角變區(圖2-A),即抗氯酸鹽突變體。這些抗氯酸鹽突變體轉移到MM培養基中培養,其菌落形態和顏色較野生型菌株在PDA培養基上有較大差異;在MM培養基上,菌絲呈無色、匍匐狀生長,無氣生菌絲生成,生長速度與野生型菌株在PDA上的生長速度大致相近,說明該菌株不能利用硝酸鹽,即為Nit突變體。一些抗氯酸鹽突變體在MM培養基上仍以野生型生長,說明該菌株即抗氯酸鹽,又能利用硝酸鹽,不屬于Nit突變體。
將獲得的Nit突變體轉移到PDA培養基中培養4 d后,再回接于MM培養基。Nit突變體中絕大部分仍不產生氣生菌絲或菌落很稀薄,即保持了Nit突變體的生長特征,但也有少數Nit突變體在MM培養基中恢復了野生型生長,即為不穩定的Nit突變體。由此可見,絕大部分Nit突變體是穩定的,只有小部分Nit突變體不穩定。
30個供試的甘蔗梢腐病菌株只有YN32未獲得Nit突變體,其余的菌株均獲得了Nit突變體。依據Nit突變體在MM、NM和HM 3種培養基上的生長特征,可將Nit突變體分為Nit1、NitM、Nit3和Nit4共4種類型的突變體(圖1)。其中大部分為Nit1突變體,占到了總突變體數的80%以上,其余3種類型的突變體較少;Nit1、NitM、Nit3和Nit4分別占突變體總數的86.09%、7.89%、5.26%和0.75%(表3)。
2.2 Nit突變體異核體自身親和性測定
30個供試菌株中只有9個菌株獲得了NitM突變體(表3),對這9個菌株進行異核體自身親和性鑒定,即菌株自身的NitM與Nit1或Nit3配對培養。結果顯示,9個菌株均能發生營養體自身親和,即兩稀疏生長的菌落交界處形成旺盛的氣生菌絲帶,這表明9個菌株都具有異核體自身親和性(圖2-B);而其余菌株未獲得NitM突變體,未能進行自身親和性測定。
2.3 甘蔗梢腐病菌營養體親和群的劃分
由于只有GX4、GX18、GX17、GX31、GX6-2、YN41、YN49、FJ9和FJ64等9個菌株獲得了NitM突變體,所以只能利用這9個菌株進行菌株間的互補配對實驗。實驗結果表明(表4),來源于廣西的GX4、GX18、GX17、GX31、GX6-2,云南的YN41和YN49,福建的FJ9和FJ64分別與不同地區的其他菌株進行互補配對實驗,均未發生營養體親和性,說明它們分屬于不同的VCG;同一地區內不同區域來源的菌株間進行互補配對實驗,亦未發生營養體親和性,其中廣西區內的柳州、河池和百色區域的菌株均獲得了NitM突變體,可進行不同區域內的菌株間的互補配對實驗,但均未發生營養體親和性,說明不同區域內的菌株來自于不同的VCG;有趣的是,同一區域內的不同菌株間,同樣沒有發生營養體親和性。由此可見,甘蔗梢腐病菌基本上為1個菌株組成1個VCG,也就是說甘蔗梢腐病菌的VCG基本上沒有同一性。
3 討論
串珠鐮刀菌引起的甘蔗梢腐病對甘蔗生產造成了重大的損失,本文利用營養體親和性方法解釋了串珠鐮刀菌內一定程度的群體結構。過去的研究表明,營養體親和性與菌株生長的環境有一定的關系,特別是培養基的pH[6]。本文中培養30個菌株所使用培養基的pH一致,所以可以排除環境因素,僅就菌株本身的營養親和性進行說明。本研究利用KClO3對甘蔗梢腐病菌株進行誘導突變,獲得Nit突變體,并通過在不同營養缺陷培養基上的篩選確定突變體的類型(Nit1、NitM、Nit3和Nit4),進而對甘蔗梢腐病菌株進行營養親和性研究。其中,NitM突變體具有較強的親和能力,而Nit1和Nit3的親和能力較弱[13];研究發現,菌株與自身的NitM都具有非常強的親和能力,而少數幾個菌株的Nit1和Nit3也具有一定的親和能力。
較早的研究報道[14-15]認為,菌株間的親緣關系取決于地理分布,來源于相同或相鄰地區的菌株具有更大的遺傳相似性。但據潘月梅[16]、余霞等[17]的研究表明,VCG與地理分布間并不存在明顯的關系,同一地域可能存在多個 VCG或同一VCG分布于多個地理區域;同時Sidhu[18]、Klittich[19]、潘月梅等[16]的研究也表明,串珠鐮刀菌的田間群體結構非常復雜。本研究中,分離自廣西、云南、福建和海南等4個地區的30個梢腐病菌株群體間的VCGs互不相同,沒有屬于同一個VCG的菌株,同時存在于4個地區或其中2個或3個地區的群體中;而來自于同一地區的甘蔗梢腐病菌株亦未發現相同的VCG。說明引起甘蔗梢腐病的串珠鐮刀菌群體內存在豐富的VCG多樣性,田間群體結構變化很大;同時也證明VCG與菌株的地理來源并沒有明顯的直接關系
據Elena等[20]的研究結果表明,同一地理區域中同一寄主品種的菌株通常為同一VCG群。本研究中,除部分菌株未能確定寄主品種外,同一地理區域的供試菌株幾乎采自不同的甘蔗品種,只有2對菌株采自相同的甘蔗品種,即YN5和YN32采自粵糖60,YN46和YN67采自ROC22;但由于本研究中YN46和YN67菌株沒有獲得NitM突變體,未能進行親和性實驗,所以不能確定它們是否屬于同一個VCG。但是本研究中發現采自同一地區不同甘蔗品種上的甘蔗梢腐病菌株沒有出現營養體親和的現象,說明營養體親和性與寄主品種也沒有相關性。
本文共分析了30個甘蔗梢腐病菌株,初步表明該群體內存在豐富的VCG多樣性,但由于樣品的局限性,沒有對不同地理區域的串珠鐮刀菌菌群結構進行詳細的分析和總結。在以后的研究中需要采集更多地理區域,更多甘蔗品種上的梢腐病菌株進行分析,以便更全面地分析甘蔗梢腐病菌的VCGs多樣性狀況以及VCG與菌株地理來源以及寄主品種之間的相互聯系。同時由于在研究串珠鐮刀菌營養體親和方面沒有成熟的技術可以借鑒,本研究也存在著一定的不足,雖然30個菌株幾乎都能夠獲得nit突變體,但是在獲得NitM突變體的篩選概率上偏低,仍需對該病原菌菌株的培養條件進行進一步的優化。
參考文獻
[1] 裘維蕃. 菌物學大全[M]. 北京: 科學出版社, 1998.
[2] Leslie J F. Fungal vegetative compatibility[J]. Annu Rev Phytopathol, 1993, 31: 127-150.
[3] Cyaley D M. The phenomenon of mutua1 aversion between mono-spore mycelium of the same fungus(Diaporthe penicosa Marchal)with a diseussion of sex heterothallism in fungi[J]. Joumal of Geneties, 1923, 13: 1 353-1 370.
[4] 吳小芹. 林木病原真菌的群體分化研究[J]. 林業科學研究, 2000, 13(4): 423-430.
[5] 龔國淑, 余 霞, 葉華智, 等. 新月彎孢菌營養體親和群鑒定方法探討[J]. 植物病理學報, 2004, 34(5): 400-408.
[6] 余功明, 史國英, 魏源文, 等. 影響水稻紋枯病菌營養體親和性分化的因子[J]. 植物病理學報, 2015, 45(3): 225-231.
[7] 劉 東, 代麗婷, 蒲子婧, 等. 黃瓜枯萎病菌毒力、營養體親和性及ISS R分析[J]. 植物病理學報, 2012, 42(5): 456-465.
[8] 景 嵐, 裴 旭, 楊柳青, 等. 向日葵黃萎病菌營養親和性及致病力分化[J]. 中國油料作物學報, 2014, 36(1): 0106-0112.
[9] 喬廣行, 丁 平, 李 濤, 等. 營養體不親和性誘導栗疫病菌菌株間程序性細胞死亡[J]. 南京農業大學學報, 2010, 33(2): 110-114.
[10] 張玉娟. 甘蔗梢腐病病原分子檢測及甘蔗組合、 品種的抗病性評價[D]. 福州: 福建農林大學, 2009.
[11] 李 明, 錢月珍, 楊雪靜, 等. 串珠鐮刀菌的營養體親和性[J].南京農業大學學報, 2000, 23(2): 31-34.
[12] Lin Z, Xu S. Species-specific detection and identification of fusarium species complex, the causal agent of sugarcane pokkah boeng in China[J]. PloS One, 2014, 9(8): e104 195.
[13] 余仲東, 曹支敏, 高愛琴, 等. 真菌營養體親和性的研究[J]. 西北林學院學報, 2002, 17(3): 80-85.
[14] Puhalla JE, Hummel M. Vegetative compatibility groups within Verticillium dahliae[J]. Phytopathology, 1983, 73: 1 305-1 308.
[15] Bell A A. Vegetative compatibility and virulence to cotton among isolates of Verticillium dahlia[J]. Phytopathology, 1992, 81: 159-162.
[16] 潘月梅, 高智謀, 營金鳳, 等. 不同地理及寄主來源的串珠鐮孢的營養體親和性研究[J]. 菌物學報, 2009, 28(2): 205-212.
[17] 余 霞, 龔國淑, 葉華智, 等. 新月彎孢菌Curvularia lunata 營養體親和性的進一步研究[J]. 菌物學報, 2006, 25(2): 292-301.
[18] Sidhu G S. Characteristics and natural occurrence of Gibberella fujikuroi mating groups A and D on sorghum and corn hosts[J]. Canadian Journal of Botany, 1985, 63: 562-566.
[19] Klittich C J R. Chlorate-resistant, nitrate-utilizing mutants of Fusarium moniliforme[J]. Journal of General Microbiology, 1989, 135: 721-727.
[20] Elena K, Paplonatas E J. Vegetatine compatibility groups with in Verticillium dahlliae isolates from different hosts in Greece[J]. Plant Pathology, 1998, 47: 635-640.
