擬南芥AP3基因植物表達(dá)載體的構(gòu)建及在煙草中的遺傳轉(zhuǎn)化
2016-05-30 10:48:04劉彩霞代麗娟劉軼葛曉蘭曲冠證
山東農(nóng)業(yè)科學(xué) 2016年2期
關(guān)鍵詞:煙草
劉彩霞 代麗娟 劉軼 葛曉蘭 曲冠證
摘要:擬南芥花器官B類特征基因?qū)儆贛ADS-box基因家族,其中APETALA3(AP3)基因在花瓣和雄蕊中特異性地表達(dá)。為探討AP3基因在花瓣和雄蕊發(fā)育中的功能,本研究從擬南芥(Arabidop.sis thaliana)花序中克隆AtAP3基因構(gòu)建植物表達(dá)載體,并轉(zhuǎn)化煙草(Nicotiana tobacum)。通過PCR及qRT-PCR分析,表明AtAP3基因成功轉(zhuǎn)入煙草基因組并表達(dá)轉(zhuǎn)錄。表型觀測顯示轉(zhuǎn)基因煙草雄蕊與野生型相比明顯變短,說明AP3基因特異性地參與雄蕊的發(fā)育并起著至關(guān)重要的作用。
關(guān)鍵詞:擬南芥;APETALA3(AP3);轉(zhuǎn)基因;煙草;雄蕊發(fā)育
中圖分類號:S188+.1
文獻(xiàn)標(biāo)識號:A 文章編號:1001-4942(2016)02-0007-05
花是被子植物的重要生殖器官。花器官的發(fā)育模型最早源于Coen和Meverowitz對擬南芥和金魚草的研究,并提出了花發(fā)育的“ABC”模型,隨后該模型被后人逐步發(fā)展為“ABCD”或“ABCE”模型。典型的花發(fā)育具體可分為四輪,分別為萼片、花瓣、雄蕊和心皮,每一輪的發(fā)育都受相應(yīng)基因的特異性調(diào)控。
隸屬于MADS-box家族的植物花器官B類特征基因在雄蕊及花瓣發(fā)育中行使功能。研究表明,B類功能基因在花器官發(fā)育時(shí)與C類功能基因共同控制雄蕊發(fā)育,同時(shí)又與A類功能基因共同調(diào)控花瓣發(fā)育,當(dāng)B類功能基因缺失,雄蕊會轉(zhuǎn)化為心皮,花瓣則轉(zhuǎn)化為萼片,形成只有心皮和萼片的不完整花器官。B類功能基因在植物進(jìn)化過程中發(fā)生了兩次基因重復(fù)事件,結(jié)果在這個(gè)亞家族中產(chǎn)生了四個(gè)不同的進(jìn)化系,分別為DEF/AP3(paleoAP3)、GLO/PI、TM6和euAP3。其中擬南芥經(jīng)歷基因重復(fù)后,其B類功能基因產(chǎn)生了屬于euAP3進(jìn)化系成員的APETAIA3(AP3)基因和屬于GLO/PI進(jìn)化系成員的PISTELIATA(PI)基因。
目前對花器官發(fā)育的研究大多以“ABC”模型為理論基礎(chǔ)來豐富其他物種的花器官發(fā)育機(jī)制,關(guān)于擬南芥AP3基因在煙草中異源表達(dá)的研究報(bào)道較少。本試驗(yàn)根據(jù)擬南芥B類功能基因的高度保守性及組織表達(dá)特異性,從擬南芥花序中克隆得到AtAP3基因,構(gòu)建植物表達(dá)載體并進(jìn)行煙草遺傳轉(zhuǎn)化。不僅對植物花器官B類特征基因AP3的功能進(jìn)行了新的補(bǔ)充,還增加了對植物花發(fā)育“ABC”模型的新認(rèn)識。
1 材料與方法
1.1 試驗(yàn)材料
野生型擬南芥(Arabidopsis thaliana Col-0)、野生型煙草(Nicotiana tobacum)種子均為本實(shí)驗(yàn)室保存。
1.2 菌株及主要試劑
大腸桿菌Transl-T1感受態(tài)購自Transgen(中國);農(nóng)桿菌EHA105由本實(shí)驗(yàn)室保存;植物表達(dá)載體pROKII質(zhì)粒,由山東師范大學(xué)張慧教授惠贈(zèng);質(zhì)粒提取、膠回收試劑盒購自O(shè)MEGA公司(美國);PCR相關(guān)試劑、DNA marker、限制性內(nèi)切酶、T4 DNA ligase、PrimeScriptrrM RT reagent Kit購自TaKaRa公司(中國大連);EasyPureTM PlantRNA Kit、pEASY-T1購自Transgen(中國);其他試劑為進(jìn)口或國產(chǎn)分析純。
1.3 試驗(yàn)方法
1.3.1 擬南芥花序總RNA提取及AtAP3基因的克隆 用EasyPure'rM Plant RNA Kit試劑盒提取擬南芥花器官的總RNA,采用PrimeScriptTM RT rea-gent Kit試劑盒進(jìn)行cDNA合成。以cDNA為模版,利用引物AtAP3-F和AtAP3-R(表1)進(jìn)行PCR擴(kuò)增。反應(yīng)程序:94℃預(yù)變性4min;94℃變性30s,55℃退火30s,72℃延伸1min,共35個(gè)循環(huán);72℃再延伸7min,PCR結(jié)束后全部產(chǎn)物進(jìn)行1%瓊脂糖凝膠電泳分析。利用凝膠回收試劑盒回收目的片段,將回收后的片段根據(jù)操作手冊介紹方法連人pEASY-T1載體中,轉(zhuǎn)化大腸桿菌Transl-T1感受態(tài),涂LB抗性平板(卡那抗性),對獲得的單克隆進(jìn)行菌落PCR驗(yàn)證,篩選后的陽性克隆命名為pEASY-T1-AtAP3,送至哈爾濱博仕生物技術(shù)有限公司測序。
1.3.2 植物表達(dá)載體構(gòu)建利用限制性內(nèi)切酶Xba I、Kpn I雙酶切重組質(zhì)粒pEASY-T1-A-tAP3,同時(shí)將pROKII載體質(zhì)粒也用XbaI、Kpn I雙酶切,并分別回收目的片段,利用T4 DNA ligase過夜連接轉(zhuǎn)化大腸桿菌Transl-T1感受態(tài),涂LB抗性平板(卡那抗性),對獲得的單克隆進(jìn)行菌落PCR驗(yàn)證,篩選后的陽性克隆命名為pROKII-AtAP3,提取質(zhì)粒,送至哈爾濱博仕生物技術(shù)有限公司測序。選取測序正確的質(zhì)粒轉(zhuǎn)化農(nóng)桿菌EHA105,提取農(nóng)桿菌中的質(zhì)粒利用引物pROKII-F和pROKII-R(表1)進(jìn)行PCR檢測。
1.3.3 轉(zhuǎn)基因株系獲得及分子檢測 通過根瘤農(nóng)桿菌介導(dǎo)法進(jìn)行煙草的遺傳轉(zhuǎn)化,轉(zhuǎn)化過程如下:預(yù)培養(yǎng)1~2d;農(nóng)桿菌(OD=0.2)侵染3min;共培養(yǎng)1~2d;脫菌。一個(gè)月后獲得抗性株系。用CTAB方法提取抗性植株的總DNA,利用引物pROKII-F和pROKII-R進(jìn)行PCR,并用1%瓊脂糖凝膠檢測,分析是否成功獲得轉(zhuǎn)基因株系。
1.3.4 實(shí)時(shí)定量RT-PCR分析檢測 用EasvPureTM Plant RNA Kit試劑盒提取轉(zhuǎn)基因植株的總RNA,經(jīng)反轉(zhuǎn)錄試劑盒進(jìn)行cDNA合成。將合成的第一鏈cDNA加去離子水稀釋10倍后作為模板,進(jìn)行定量檢測,引物為AtAP3-RT-F和AtAP3-RT-R(表1)。反應(yīng)體系為20μL:其中2×SYBR Green實(shí)時(shí)熒光染料混合液10μL,ROXDye Ⅱ0.4μL,2μL cDNA模板,正、反向引物各0.8μL。以NtActin基因作為內(nèi)參,擴(kuò)增反應(yīng)條件為:95℃30s;95℃5s,60℃35s,40個(gè)循環(huán);95℃15s,60℃lmin,95℃15s。每個(gè)樣品進(jìn)行3次重復(fù),并通過2-△△Ct法進(jìn)行計(jì)算。
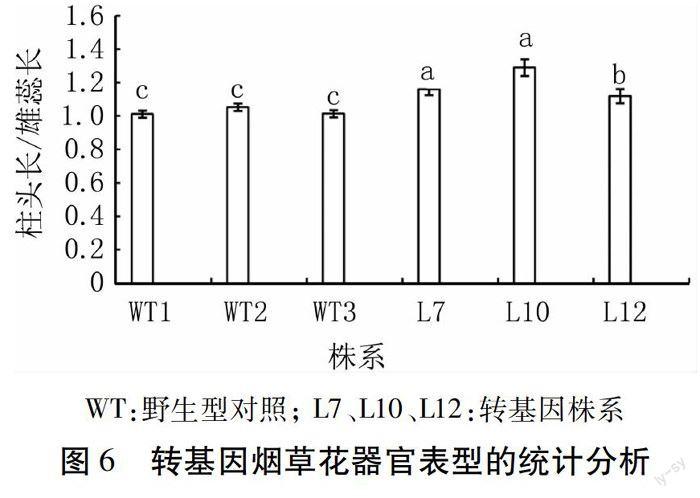
1.3.5 轉(zhuǎn)基因植株的表型觀測 挑選目的基因表達(dá)量較高的3個(gè)轉(zhuǎn)基因株系及對照株系分別移栽10棵至溫室,對其進(jìn)行定期觀察,待進(jìn)入盛花期后,在頂部花序中,每株隨機(jī)選取5朵剛剛開放的花,分別測量其花瓣、柱頭及雄蕊的長度,對測量結(jié)果利用SPSS 19.0軟件進(jìn)行SNK差異性統(tǒng)計(jì)分析。
2 結(jié)果與分析
2.1 擬南芥花序AtAP3基因的克隆
利用RNA提取試劑盒從擬南芥花序中提取總RNA,經(jīng)濃度及質(zhì)量檢測可用于下一步試驗(yàn)。將提取的RNA經(jīng)反轉(zhuǎn)錄試劑盒反轉(zhuǎn)錄為cDNA作為模板,利用引物AtAP3-F和AtAP3-R進(jìn)行PCR擴(kuò)增,1%瓊脂糖電泳檢測表明,擴(kuò)增條帶清晰且和預(yù)期大小一致(圖IA)。將目的條帶切膠回收后與克隆載體pEASY-T1連接,轉(zhuǎn)化大腸桿菌,經(jīng)菌落PCR驗(yàn)證(圖1B),將獲得的陽性克隆命名為pEASY-T1-AtAP3。
2.2 植物表達(dá)載體的構(gòu)建
對獲得的pEASY-T1-AtAP3質(zhì)粒和pROKII載體質(zhì)粒,同時(shí)利用限制性內(nèi)切酶XbaI、Kpn I進(jìn)行雙酶切,反應(yīng)結(jié)束用1%瓊脂糖凝膠電泳檢測(圖2A),分別回收目的片段。利用T4 DNA ligase進(jìn)行連接反應(yīng)后,轉(zhuǎn)化大腸桿菌Transl-T1感受態(tài),挑取單菌落進(jìn)行PCR檢測,獲得750 bp左右目的條帶(圖2B),測序結(jié)果顯示連接正確,基因未發(fā)生突變。提取質(zhì)粒轉(zhuǎn)化農(nóng)桿菌EHA105,挑取單克隆并進(jìn)行PCR檢測(圖2C),結(jié)果顯示所選單菌落均為陽性克隆,表明植物表達(dá)載體構(gòu)建成功。
2.3 轉(zhuǎn)基因植株的獲得及PCR檢測
用含有pROKII-AtAP3的農(nóng)桿菌EHA105侵染煙草葉片,經(jīng)4~6周的選擇培養(yǎng),從葉片周圍生成大量抗性芽。待抗性芽長出葉片后,將葉片切下放在含有Kan(50mg·L-l)抗生素的分生培養(yǎng)基上繼代培養(yǎng)。將經(jīng)兩次分化獲得的芽在生根培養(yǎng)基中進(jìn)行生根定植,最終共獲得18株轉(zhuǎn)基因煙草植株。提取抗性轉(zhuǎn)基因株系的基因組DNA,利用表1中的pROKII通用引物對其進(jìn)行PCR擴(kuò)增檢測(圖3),結(jié)果顯示抗性植株中均含有目的基因,說明外源基因已經(jīng)整合人煙草基因組中。
2.4 轉(zhuǎn)基因植株的熒光定量PCR檢測
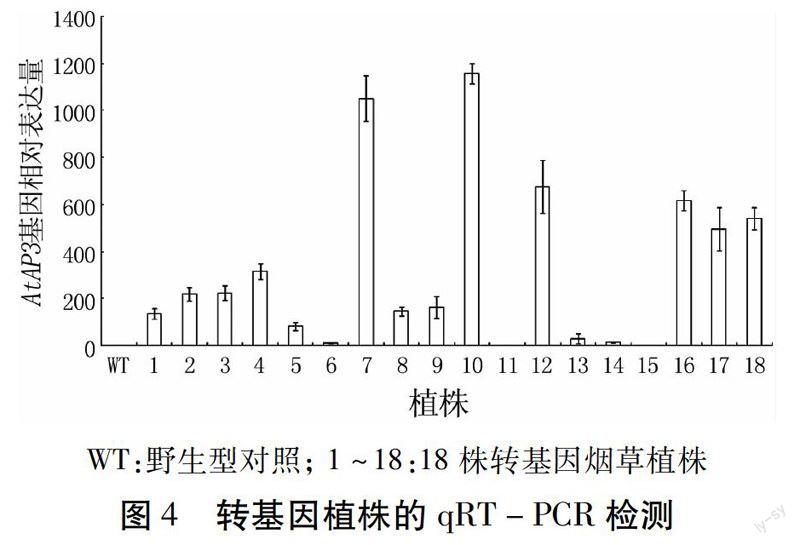
為了比較AtAP3基因在煙草植株中的表達(dá)情況,提取煙草葉片總RNA進(jìn)行熒光定量PCR檢測,結(jié)果如圖4。所有的轉(zhuǎn)基因株系中均有AtAP3的表達(dá),其中7、10、12、16、17、18號株系表達(dá)量最高;1、2、3、4、5、8、9號株系表達(dá)量低于以上幾個(gè)株系;表達(dá)量最低的株系是6、1 1、13、14、15。
2.5 轉(zhuǎn)基因株系表型觀察
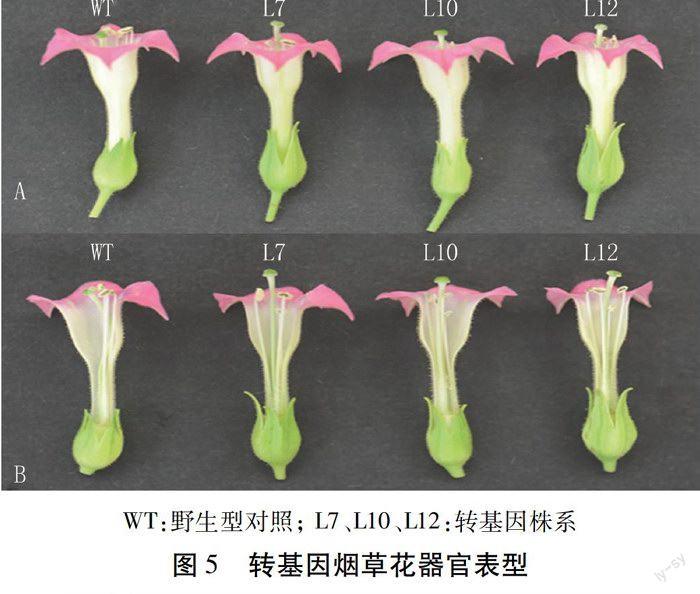
經(jīng)過定量PCR檢測,篩選目的基因表達(dá)量高的3個(gè)轉(zhuǎn)基因株系和野生型煙草,移栽到溫室中。表型觀測結(jié)果顯示,在營養(yǎng)生長期,轉(zhuǎn)基因植株與野生型相比無明顯性狀差異;待進(jìn)入花器官發(fā)育成熟期后,轉(zhuǎn)基因株系雄蕊變短(圖5)。分別選取3個(gè)轉(zhuǎn)基因株系和對照株系(每個(gè)株系頂端隨機(jī)取5朵花),測量柱頭和雄蕊的長度,用每一朵花的柱頭長度與其雄蕊長度做比值,再取比值的平均值,利用SPSS 19.0軟件,采用SNK方差齊性檢驗(yàn)的方法對其差異顯著性進(jìn)行分析,結(jié)果如圖6。轉(zhuǎn)基因株系與野生型株系比,雄蕊變短,且差異顯著;3個(gè)轉(zhuǎn)基因株系之間L10>L7>L12,與實(shí)時(shí)定量結(jié)果一致,表明AtAP3基因表達(dá)量越高雄蕊縮短越明顯。上述結(jié)果表明,過表達(dá)AtAP3基因影響轉(zhuǎn)基因煙草植株雄蕊的發(fā)育,進(jìn)而導(dǎo)致轉(zhuǎn)基因株系結(jié)種率下降。
3 討論
自“ABC”模型提出之后,許多人都從這三類特征基因著手,研究不同物種花器官發(fā)育機(jī)制,得到了許多類似的基因。基于植物之間的親緣遠(yuǎn)近、物種差異、進(jìn)化程度強(qiáng)弱等特點(diǎn),同一種功能基因會出現(xiàn)序列差異,但該類基因均保留有相同的保守區(qū)域,來保證行使相應(yīng)的功能。其中屬于MADS-box家族的B類特征基因在花瓣和雄蕊的發(fā)育中行使功能,其在擬南芥中包括PI和AP3兩個(gè)成員。被子植物的B類功能基因在具有高度保守性的同時(shí)隨物種的進(jìn)化等原因產(chǎn)生序列多態(tài)性,其中包括一個(gè)PI基因系和三個(gè)AP3-like基因系,且AP3/PI基因較其他功能基因的進(jìn)化速度快40%左右,這使得B類功能基因產(chǎn)生多元結(jié)構(gòu),主要表現(xiàn)為雄蕊及花瓣的形態(tài)多樣性。
本試驗(yàn)為探究AtAP3基因異源表達(dá)的功能,從擬南芥花序中克隆得到AtAP3基因,并將其轉(zhuǎn)人煙草,與野生型相比,轉(zhuǎn)基因煙草的花器官發(fā)生明顯變化,主要表現(xiàn)為雄蕊的長度短于對照植株,花冠長度也有變短現(xiàn)象,但變化不明顯。前人研究表明,花瓣是由A類功能基因與B類功能基因中的PI基因共同作用形成的,而雄蕊則是由B類功能基因中的AP3基因與C類功能基因共同作用形成的,本研究中雄蕊發(fā)育發(fā)生變化導(dǎo)致轉(zhuǎn)基因植株的育性下降甚至敗育,這與AP3基因調(diào)控雄蕊發(fā)育的理論相一致,表明AtAP3在異源調(diào)控植物的花器官發(fā)育方面有一定作用,AtAP3基因與煙草中的B類基因和C類基因共同作用從而抑制雄蕊發(fā)育。但外源AtAP3基因在轉(zhuǎn)基因煙草中是通過怎樣的方式調(diào)控雄蕊形態(tài)變化仍需進(jìn)一步深入探究,以期對該現(xiàn)象作出機(jī)理性的闡述,挖掘出其潛在的應(yīng)用價(jià)值。
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學(xué)報(bào)(2019年5期)2019-11-14 07:54:12
首都公共衛(wèi)生(2019年5期)2019-05-21 01:08:34
浙江中西醫(yī)結(jié)合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當(dāng)代化工研究(2016年9期)2016-03-20 16:22:08
自動(dòng)化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學(xué)林(2013年2期)2013-11-12 12:58:54
中國煙草學(xué)報(bào)(2012年5期)2012-04-12 06:21:18
