油棕花序性別分化過程觀察
2016-05-30 14:52:53楊靜李哲曾憲海林位夫鄒積鑫潘登浪李煒芳劉釗周建南劉潔瓊張春紅
南方農業學報 2016年9期
楊靜 李哲 曾憲海 林位夫 鄒積鑫 潘登浪 李煒芳 劉釗 周建南 劉潔瓊 張春紅
摘要:【目的】了解油棕雌、雄花序的發育規律及在形態學和細胞組織學水平上的差異,為進一步研究油棕花序性別分化機理打下基礎。【方法】取油棕不同發育時期的花序,測量其長度和質量,制作石蠟切片后利用體視顯微鏡觀察其細胞組織學特點。【結果】油棕花序的整個發育過程可分為兩個階段:雌雄未顯性期和雌雄顯性期,且雌雄未顯性期發育時間比雌雄顯性期長得多,約占總時期的80%。雌雄未顯性期與雌雄顯性期間存在一個花序長度和質量呈指數式迅速增長的時期,即花序發育指數增長期(EGSI)。油棕花性別分化之初,花序開始迅速發育,進入EGSI。油棕雌、雄花序在發育過程中的不同之處表現為:雌花有一個兩性期階段,由一個有功能的雌花及兩側的伴隨雄花組成,伴隨雄花選擇性敗育,雌花成為單性花;而性別分化后,雄花不存在兩性期階段,直接進入單性期階段。油棕雌、雄花序每個小穗軸上花原基分生組織數目明顯不同,雄花花原基的數量遠多于雌花花原基,具有數量級差別。【結論】油棕花序發育過程中存在EGSI。幼嫩雌、雄花序的花原基分生組織數目顯著不同,根據花原基分生組織的數量可判斷尚未從葉片基部露出的幼嫩花序性別。
關鍵詞: 油棕;花序;雌花;雄花;發育分化;細胞組織學
中圖分類號: S565.9 文獻標志碼:A 文章編號:2095-1191(2016)09-1533-09
Abstract:【Objective】Developmental law of oil palm female and male inflorescences were studied, and the differences between pistillate flowers and staminate flowers in morphology and cytohistology were investigated, in order to lay foundation for further sex differentiation research. 【Method】The inflorescences were collected at different developmental stages, their length and mass were measured. Then, the paraffin sections were made, so as to observe its cytohistology characters with microscope. 【Result】The whole developmental process of oil palm inflorescences could be divided into two stages viz., sex unrevealed stage and sex revealed stage. The development at sex unrevealed stage took more time than that at sex revealed stage, accounting for 80% of total period. There was a exponential growth stage to be found between sex unrevealed stage and sex revealed stage, at this stage, both length and mass of inflorescences increased rapidly and exponentially, it was denominated exponential growth stage of inflorescence(EGSI). At beginning of floral sex differentiation, the oil palm inflorescences started to develop rapidly, entering EGSI. The remarkable differences between pistillate flowers and staminate flowers during developmental process were as follows: There was a bisexual period during developmental process of pistillate flower, and pistillate flower was composed of a single pistillate flower flanked by two accompanying staminate flowers, then accompanying staminate flowers selectively aborted and the pistillate flower transformed into an unisexual flower; however, there was not a bisexual period during developmental process of a staminate flower, and staminate flower directly entered unisexual stage after sex differentiation. Moreover, there were some significant differences in number of floral primodium meristem between male and female inflorescences, and that of male inflorescences was more than that of female, with some differences in order of magnitude. 【Conclusion】There exist EGSI during developmental process of oil palm inflorescence. And there are some significant differences in number of floral primodium meristem between male and female inflorescences. According to numbers of floral primodium meristem, the gender of a young inflorescence which has not emerged from the base of the relevant leaf can be judged.
Key words: oil palm; inflorescence; pistillate flower; staminate flower; development and differentiation; cytohistology
0 引言
【研究意義】植物花序性別分化是有花植物發育過程中極其重要的階段,是植物生命延續的關鍵環節,也是一種特殊的器官發生現象(趙密珍等,2016)。植物花序性別分化過程包括細胞組織學特征及相關內源激素含量的變化,了解其變化規律對揭示其發育生物學和生殖生物學特性、在生產中實現花期和性別比例調控、單性材料的繁殖及雜交、雜種優勢利用等具有重要意義。油棕是重要的木本油料作物,其單位面積產油量遙遙領先于世界上其他油料作物,被稱為世界油王(高尚士,1994;林秀香和陳振東,2007;李靜等,2014)。油棕花序發育及性別分化過程決定能否形成優質的雌、雄花序和果實,是獲得高產優質棕櫚油的前提條件。因此,開展油棕花序性別分化過程研究對獲得高產優質的棕櫚油具有重要意義。【前人研究進展】植物花序性別的不同主要表現在花器官上,即雌蕊和雄蕊的不同。目前,關于花序性別分化的研究可分為四個方面:花序結構發育過程研究、環境因子影響、植物生長調節劑影響和分子生物學研究,其中花序發育細胞組織學結構研究是上述研究的基礎。細胞組織學石蠟切片是研究植物花序性別分化的重要技術,是觀察細胞組織形態結構、研究其形態變化的主要方法之一。胡青等(2009)通過組織學石蠟切片技術觀察平基槭花性別分化過程,發現雄花中雌蕊原基的選擇性敗育發生在大孢子母細胞減數分裂為4個大孢子時期。高英(2011)以核桃雄花芽為材料,用形態觀察和組織切片的方法研究了雄花芽與樹體的變化,結果發現雄花芽的外部形態及內部結構變化與核桃樹的物候期間存在密切相關性。王秀榮等(2011)利用組織學石蠟切片技術對麻瘋樹花進行觀察,發現其雌花在發育進程中會經歷一個兩性期,隨后雄蕊停止發育,形成功能上的單性花,無兩性期階段。油棕只有一個營養型頂端分生組織,每張葉子的腋下均有一個花序。油棕花序為雌雄同株異序,從花序開始發育至開放需經歷2~3年的時間,過程漫長,顯著區別于其他大多數植物。油棕雌花序近頭狀,密集;雄花序由多個指狀的穗狀花序組成。雌花序的花序軸由150個左右小穗軸組成,而雄花序的花序軸由100~300個小穗軸組成(Jacquemard,1995)。成熟雄花序花序軸下的花梗約40 cm,成熟雌花序花序軸下的花梗20~30 cm。2005年,Adam等對油棕花序進行細胞組織學研究,發現其雌花序由一個有功能的雌花及兩側的敗育雄花組成,而雄花序由單一的有功能的雄花組成。【本研究切入點】至今,國內尚無油棕花序性別分化細胞組織學研究的相關報道。【擬解決的關鍵問題】取油棕不同發育時期的花序,測量其長度和質量,并制作細胞組織石蠟切片進行顯微結構觀察,找出雌、雄花序的發育規律及在形態學和細胞組織學水平的差異,為進一步研究油棕花序性別分化機理打下基礎。
1 材料與方法
1. 1 試驗材料
油棕花序取自中國熱帶農業科學院橡膠研究所農業部油棕品種區域適應性種植基地(海南省儋州市),品種為從馬來西亞引種的熱油13號,樹齡16年。
1. 2 試驗方法
1. 2. 1 取材與固定 取樣時將整棵油棕成齡樹砍倒,以莖頂端分生組織記為0,最靠近的葉片記為N1,依次記為N2、N3……,然后按順序對葉片及其葉腋的花序或花序原基細胞進行排序編號。將采集的花芽立即投入FAA固定液中浸泡至少24 h,也可長期保存。
1. 2. 2 花芽長度和質量測量 較大的花芽直接用刻度尺依次測量。序號N1~N29的花序小,用裝有Leica DFC 450C攝像頭的Leica M205 FA熒光體視顯微鏡觀察和拍照,再根據照片中的標尺用軟件Image J計算長度。
1. 2. 3 制作石蠟切片 將固定好的材料依次在30%、50%、70%和100%的脫水劑中脫水2 h,然后進行透明、浸蠟、包埋。利用Leica RM2235切片機進行切片,厚度7~10 μm。用Schiff試劑和苯胺藍黑染色后(趙志虎,2007),用裝有Leica DFC 500攝像頭的Leica DMLB顯微鏡觀察和拍照。
1. 2. 4 花組織切片觀察和花原基分生組織數量統計
將較大花芽的小穗軸縱切,全部雌花及雄花橫切,對連續切片進行觀察;并根據連續切片中花原基分生組織的位置和形態統計其數量。
2 結果與分析
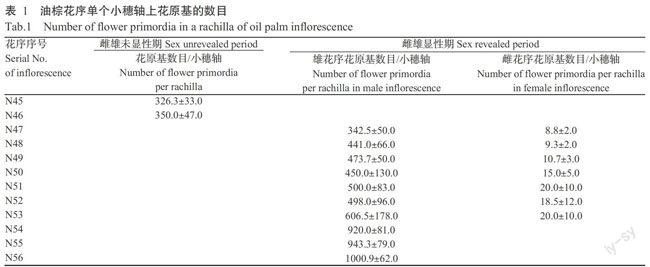
油棕花序的發育過程可分為兩個主要階段:雌雄未顯性期和雌雄顯性期。雌雄未顯性期發育時間比雌雄顯性期長得多,約占總時期的80%。
2. 1 花序大小
以油棕花序全長和質量為參數繪制油棕雌雄未顯性期和雌雄顯性期花序的生長曲線。
從圖1~4可以看出,N1~N44階段的花序發育速度非常緩慢,從N45開始花序迅速發育,花序長度和質量呈指數式顯著增長,故稱這一時期為花序發育指數增長期(The exponential growth stage of the inflorescence,EGSI)。油棕花性別分化之初,花序開始迅速發育,進入EGSI。雌、雄花序的生長曲線大致相同,整體上表現為雄花序的長度大于雌花序。作為花序的保護性器官,苞片長度和質量的增長在花序生長中占據重要位置;去除苞片的花序質量增長相對于苞片的質量增長有遲滯現象。隨著花序不斷發育,苞片持續生長并木質化。
2. 2 花序發育的細胞學特征
2. 2. 1 雌雄未顯性期 N1階段,在葉原基下存在一簇細胞,為花序原基(圖5-1),大小為0.30 mm,是在光學顯微鏡下看到的花序發育最早時期;此時細胞含有1個較大的細胞核。N6階段,花序原基起始發育出先出葉原基(圖5-2),整個花序大小為0.85 mm;此時花序原基的中央為圓頂狀,兩側為先出葉原基,隨后先出葉伸長(圖5-3)。N10階段,先出葉完全包裹住花序原基,此時整個花序大小為1.15 mm,且花序原基兩側出現花梗苞片原基(圖5-4)。花梗苞片原基細胞繼續分裂、生長成為花梗苞片(圖5-5);先出葉出現維管化,花梗苞片繼續伸長。N14階段,花梗苞片完全包裹住花序分生組織(圖5-6),此時花序長度為2.20 mm。當先出葉和花梗苞片包裹住花序分生組織后,花序軸兩側出現小穗軸苞片原始細胞,此時為N15花芽階段(圖5-7),花序長度仍為2.20 mm。小穗軸苞片原始細胞繼續發育成小穗軸苞片原基、小穗軸苞片(圖5-8),此時花梗苞片出現維管化。隨著小穗軸苞片的生長及數目的增多,在小穗軸苞片基部出現小穗軸分生組織(圖5-9),即進入N27花序階段,此時花序長度為10.00 mm。小穗軸分生組織細胞有較大的核質比,其發育方式為自頂部向基部發育(圖5-10),此時在花序軸頂部的小穗軸苞片基部已出現小穗軸分生組織,但基部的小穗軸苞片尚未出現小穗軸分生組織。小穗軸分生組織呈圓錐形(圖5-11),圍繞花序軸呈螺旋狀排列(圖5-12)。小穗軸分生組織繼續發育,至N36階段時,小穗軸分生組織發育出花苞片,此時花序大小為26.50 mm(圖5-13),且頂部小穗軸的發育速度快于基部的小穗軸(圖5-14)。
2. 2. 2 雌雄顯性期
2. 2. 2. 1 正常雌花的發育 在雌花序中,小穗軸上每個花苞片下會長出一朵三基花。三基花由中間一朵有功能的雌花及其兩側的伴隨雄花組成,發育到一定階段,伴隨雄花會敗育脫落。三基花呈蝎尾狀聚傘花序。
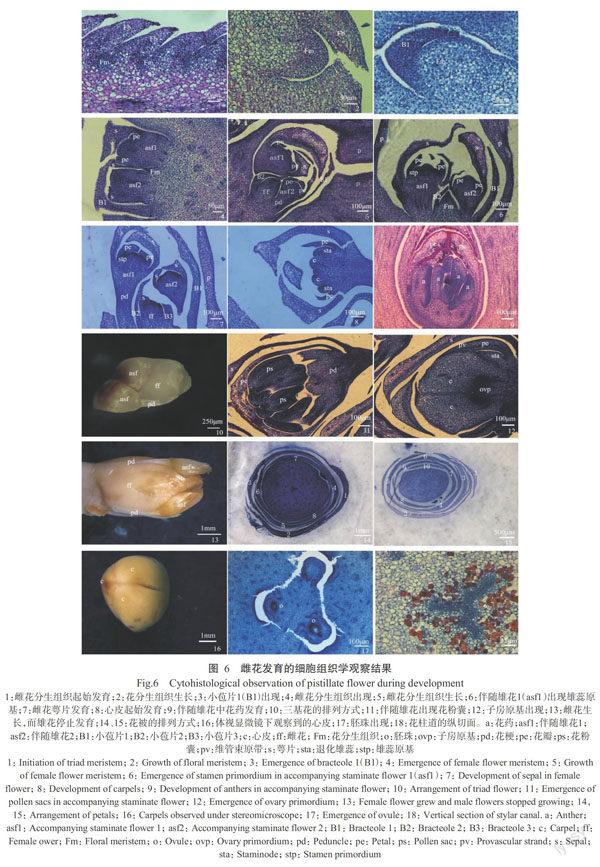
約至N47階段,小穗軸分生組織形成的三基花苞片腋下出現花分生組織(圖6-1),此時花分生組織的細胞含有一個較大的細胞核,處于活躍狀態。花分生組織由三部分組成(圖6-2):最外層是一層明顯的細胞(L1),將來發育成表皮細胞;L1層下是數層活躍的等徑細胞,含有較大的細胞核;等徑細胞下的區域內,細胞核質比較小,細胞中有液泡出現(L3)。花分生組織分化出小苞片1(B1),B1腋下發育出伴隨雄花1(asf1)(圖6-3)。在B1腋下與asf1間出現一簇細胞,將來發育出小苞片2(B2),B2腋下靠近B1的位置隨后發育出伴隨雄花2(asf2)。位置排列順序為B1→asf2→B2→asf1。asf1與asf2的發育并不同步,表現為asf1發育早于asf2。
N48階段,在B2腋下與asf2間出現一簇分生細胞(圖6-4),隨后發育出小苞片3(B3),B3腋下發育出雌花(ff),ff位于B2和B3之間。此時asf1已發育出萼片和花瓣原基,asf2的萼片原基也已出現。這種發育方式符合蝎尾狀聚傘花序的特征。
N49階段,伴隨雄花的萼片和花瓣均已出現(圖6-5),且部分伴隨雄花已發育出雄蕊原基(圖6-6)。雄花萼片末端的細胞液泡化,此階段雄花的花梗伸長,直至高于雌花。雌花的分生組織生長變大,但只發育出B3,此時的雌花小于雄花。
N51階段,雌花的分生組織開始發育,而此時雄花保持原樣(圖6-7)。隨后,雌花的生殖器官繼續發育,發育出雌蕊群和雄蕊群(圖6-8),其萼片末端的細胞液泡化。同時,伴隨雄花發育出花藥(圖6-9)。花藥的一側細胞出現液泡化,另一側細胞則有一個較大的細胞核。
N52階段,雌花發育速度快于雄花。雌花迅速變大,而伴隨雄花大小保持不變(圖6-10)。伴隨雄花的萼片、花瓣及花藥末端的一些細胞中積累多酚,花瓣的一些細胞空泡化。同時,花藥中出現花粉囊(圖6-11)。雌花中,心皮基部已發育出子房原基(圖6-12);雌花萼片、花瓣末端的細胞出現液泡化現象。
N53階段,雌花繼續發育,雄花仍保持不變(圖6-13)。伴隨雄花會在雌花成熟前脫落。絕大多數雌花有3片萼片、3片花瓣(圖6-14,其中1、2為B2和B3),個別雌花有4片萼片、4片花瓣(圖6-15,其中1、2為B2和B3);其外形和質地相同,均呈淡黃色。剝去花瓣和萼片,在體視顯微鏡下觀察,可觀察到雌花有3個自由心皮(圖6-16)。根據雌花橫切面可以看出,子房為3心室,每個心室均有一個胚珠(圖6-17)。在心皮的基部和末端間有一個通道,為花柱道(圖6-18),可能是花粉管在受精前的通道。
2. 2. 2. 2 正常雄花的發育 通過觀察具有完整雄花發育過程的成齡油棕樹,發現正常雄花的發育特征如下:
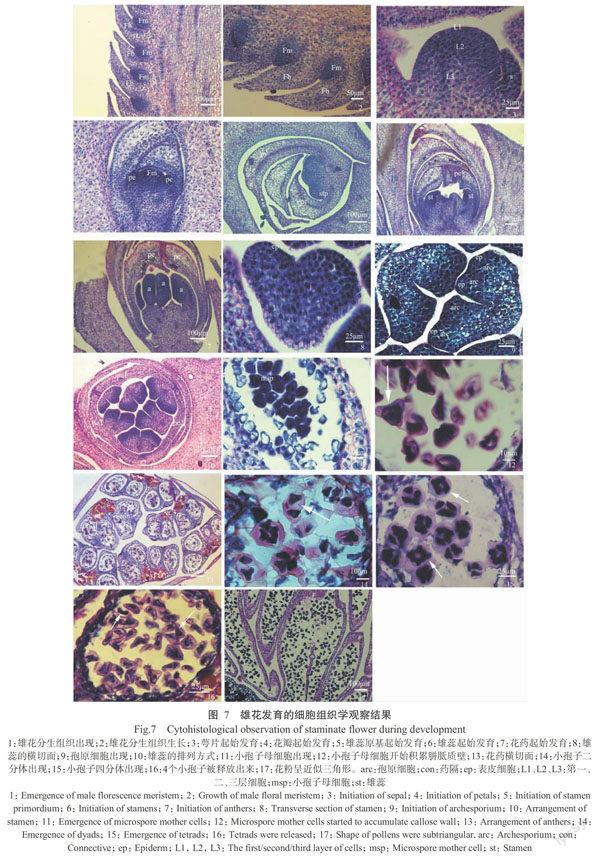
約N47階段,花苞片下長出花分生組織(圖7-1)。花分生組織呈圓錐形,細胞內均有一個較大的細胞核(圖7-2 )。
N49階段,花分生組織一側長出萼片(圖7-3)。雄花的分生組織也可分為3層:最外層是一層明顯的細胞(L1),將來發育成表皮細胞;L1層下是數層活躍的等徑細胞,含有較大的細胞核(L2);等徑細胞下的區域內,細胞核質比較小,細胞中有液泡出現(L3)。
N50階段,花分生組織兩側起始發育出花瓣原基(圖7-4),此時萼片末端的細胞出現液泡化。
N51階段,雄花的雄蕊原基開始發育(圖7-5),此時萼片及花瓣末端區域的細胞液泡化。
N52階段,雄蕊出現(圖7-6)。
N53階段,雄花中花藥出現(圖7-7),此時花藥的細胞內均含有一個較大的細胞核,表明發育初期的花藥由分裂活躍的細胞組成。由花藥縱切面可以看出,花藥由最外層的一層表皮和內部的同型細胞組成(圖7-8)。隨著花藥的發育,表皮下出現4組孢原細胞,孢原細胞的細胞核大于表皮內部其他細胞的細胞核(圖7-9)。由雄花橫切面可以看出:每朵雄花有3片萼片、3片花瓣,6個雄蕊(圖7-10)。孢原細胞通過1次平周分裂分為內外兩層,內層為造孢細胞,將來發育成小孢子母細胞,外層為初生壁細胞。
N55階段,花粉囊內出現小孢子母細胞且開始進行減數分裂。小孢子母細胞較大,細胞質濃(圖7-11)。
N56階段,小孢子母細胞開始積累胼胝質壁,且小孢子母細胞的細胞核較大(圖7-12),此時觀察花藥橫切面可看到其橫切面似蝶形(圖7-13)。隨后,小孢子母細胞開始進行減數分裂,第一次減數分裂后產生一個小孢子二分體(圖7-14),第二次減數分裂后形成4個單倍體細胞,即4個小孢子。減數分裂剛完成時,4個小孢子仍被包裹在胼胝質內,排列成四面體型(圖7-15)。隨著胼胝質壁的消失,4個小孢子被釋放出來(圖7-16),剛釋放出來的小孢子形狀不規則。小孢子繼續發育,最終形成似三角形的花粉粒(圖7-17)。
2. 3 雌雄未顯性期的花序性別判斷
成熟花中,雌花序每個小穗軸上長有5~30朵三基花,而雄花序每個小穗軸上長有400~500朵雄花。由此可見,每個小穗軸上,雄花花原基的數量遠多于雌花花原基數量,具有數量級差別,是由基因表達所決定。因此,根據每個小穗軸上花原基的數量可斷定花序性別。本研究通過細胞組織學切片觀察,發現雌雄未顯性期中,N45、N46階段每個小穗軸上的花原基數目均遠多于雌花花原基數目,而處于雄花花原基數目的區域(表1),與雌花花原基數目具有數量級差別,且均處于生長狀態,花原基的數目仍在增加,因此斷定所采的N45、N46花序為雄花序。
3 討論
油棕花序為雄雌同株異序,生殖生長時間很長,從花原基分生組織形成到花序完全開放需經歷2年以上的時間。根據花序的發育狀態,其發育過程可分為兩個階段:雌雄未顯性期和雌雄顯性期。雌雄未顯性期所占時間比雌雄顯性期長得多,根據本研究測算結果可知,前者約占總時期的80%。本研究發現,油棕雌雄未顯性期與雌雄顯性期間存在花序長度和質量呈指數式顯著增長的EGSI,油棕花性別分化之初,花序開始迅速發育,進入EGSI。雌、雄花序的生長曲線基本相似,整體上表現為雄花序的長度大于雌花序。苞片長度和質量的增長在花序生長中占據重要位置。胡樹平(2011)研究了向日葵產量形成及農藝調控機理,發現向日葵單株及群體干物質積累符合典型的S形生長曲線,出苗后54 d內為指數增長階段,55~75 d為直線增長階段,76~110 d為緩慢增長階段。裴艷梅(2015)研究發現,金絲小棗與無核小棗縱橫徑、單果重和果形指數發育趨勢基本相似,從花后5~25 d果實處于緩慢增長期,從花后30 d左右開始果實縱橫徑迅速增長,至花后45 d左右增長趨于平緩;單果重從花后20 d增長明顯,尤其從花后30 d開始呈指數增長趨勢。可見,植物整體或某些器官在其發育過程中存在EGSI。
本研究結果表明,油棕雌花在性別分化前期有一個兩性期階段,由一個位于中央且有功能的雌花和分別在兩側的2個伴隨雄花組成,隨著花序的發育,伴隨雄花選擇性敗育,在開花之前脫落;而雄花在雌雄顯性期初期其雌蕊就不再生長,僅保留了一個退化雌蕊,不存在兩性期階段,直接進入單性期。這種雌、雄花間的差異與麻瘋樹的發育情況相似(王秀榮等,2011),即一旦進入花序性別分化期后,很難對雌、雄花的性別進行轉換,因此要提高雌花比例,應在性別分化前采取相應措施。通過細胞組織學觀察研究,發現油棕雌、雄花序每個小穗軸上花原基分生組織數目明顯不同,雄花花原基數量遠多于雌花花原基,具有數量級差別,依據每個小穗軸上花原基的數量即可斷定雌雄未顯性期的花序性別。
油棕雌、雄花的發育方式均為外始式。成熟的雌花只有一個具有功能的雌花,絕大多數有3片花萼、3片花瓣,個別雌花具有4片花萼、4片花瓣,其形態結構基本相同,顏色呈淡黃色,排列方式為覆瓦狀。雌蕊由3個心皮組成,子房位于雌蕊的基部,具有3室,每室內都有一個胚珠。在心皮的基部與末端間有一個通道,為花柱道,可能是花粉管在受精前的通道,與棕櫚科的蠟油棕(Balhara et al.,2013)、尖山椰子(Casta o et al.,2014)的子房發育情況相同。成熟的雄花具有3片萼片、3片花瓣,與雌花類似,其形態結構也相同,排列方式亦為覆瓦狀。雄花具有6個雄蕊,每個雄蕊均由一個花絲和一個二裂片花藥組成。每個花藥有4個花粉囊,最終形成的花粉形狀為類似三角形。
4 結論
油棕雌雄未顯性期與雌雄顯性期間存在花序長度和質量顯著增長的EGSI,油棕花性別分化之初花序開始迅速發育,進入EGSI,即油棕花序的發育過程中存在指數增長期。幼嫩雌、雄花序的花原基分生組織數目顯著不同,根據花原基分生組織的數量可判斷尚未從葉片基部露出的幼嫩花序性別。
參考文獻:
高尚士. 1994. 高產油料樹——油棕[J]. 林業科技通訊,(11):41.
Gao S S. 1994. High-oil tree——oil palm[J]. Forest Science and Technology,(11):41.
高英. 2011. 核桃花芽分化組織學和激素原位分析研究[D]. 北京:中國林業科學研究院.
Gao Y. 2011. Histology and in situ hormones of flower-bud differentiation in walnut[D]. Beijing:Chinese Academy of Forestry.
胡青,李鳳蘭,杜藝連,郭惠紅. 2009. 平基槭花性別分化的細胞形態研究[J]. 植物研究,29(2):136-140.
Hu Q,Li F L,Du Y L,Guo H H. 2009. Morphological and cytological study on floral sex differentiation of Acer truncatum Bge.[J]. Bulletin of Botanical Research,29(2):136-140.
胡樹平. 2011. 向日葵產量形成及農藝調控機理[D]. 呼和浩特:內蒙古農業大學.
Hu S P. 2011. Yield formation and related agronomical control mechanism in sunflower[D]. Hohhot:Inner Mongolia Agricultural University.
李靜,王永,雷新濤,楊耀東,肖勇,夏薇. 2014. 油棕育種現狀及關聯分析在油棕分子輔助育種中的應用展望[J]. 江西農業學報,26(11):16-20.
Li J,Wang Y,Lei X T,Yang Y D,Xiao Y,Xia W. 2014. Current situation of oil palm(Elaeis guineensis) breeding and applicationprospects of association analysis in molecular-assisted breeding of oil palm[J]. Acta Agriculturae Jiangxi,26(11):16-20.
林秀香,陳振東. 2007. 我國棕櫚科植物的研究進展[J]. 熱帶作物學報,28(3):115-119.
Lin X X,Chen Z D. 2007. Advances on studies of palmae plant in China[J]. Chinese Journal of Tropical Crops,28(3):115-119.
裴艷梅. 2015. 金絲小棗和無核小棗4CL基因的克隆和表達研究[D]. 保定:河北農業大學.
Pei Y M. 2015. Molecular cloning and expression analysis of 4CL gene in Ziziphus jujuba Mill. ‘Jinsixiaozao and Z. jujuba Mill. ‘Wuhexiaozao[D]. Baoding:Agricultural University of Hebei.
王秀榮,丁貴杰,李平,劉烈志. 2011. 麻瘋樹花的形態和解剖結構[J]. 林業科學,47(9):57-61.
Wang X R,Ding G J,Li P,Liu L Z. 2011. Morphological and anatomical structure of the barbadosnut[J]. Scientia Silvae Sinicae,47(9):57-61.
趙密珍,龐夫花,袁華招,于紅梅,蔡偉建. 2016. 不同栽培條件下草莓品種寧玉花序分化進程[J]. 江蘇農業學報,32(1):196-200.
Zhao M Z,Pang F H,Yuan H Z,Yu H M,Cai W J. 2016. Floral bud differentiation of strawberry Ningyu under conditions of forcingcultivation and open field cultivation[J]. Jiangsu Journal of Agricultural Sciences,32(1):196-200.
趙志虎. 2007. 荔枝花性分化雄蕊發育過程的細胞化學和超微結構研究[D]. 福州:福建農林大學.
Zhao Z H. 2007. Studies on cytochemistry and ultrastructure of stamen development in litchi(Litchi chinensis Sonn.) sex differentiation[D]. Fuzhou:Fujian Agriculture and Forestry University.
Adam H, Jouannic S, Escoute J, Duval Y, Verdeil J L, Tregear J W. 2005. Reproductive developmental complexity in the African oil palm(Elaeis Guineensis,Arecaceae)[J]. American Journal of Botany,92(11):1836-1852.
Balhara M,Stauffer F W,Balslev H,Barfod A S. 2013. Floral structure and organogenesis of the wax palm Ceroxylon ce-
riferum(Arecaceae;Ceroxyloideae)[J]. American Journal of Botany,100(11):2132-2140.
Casta o F,Stauffer F,Marquinez X,Crèvecoeur M,Collin M,Pintaud J C,Tregear J. 2014. Floral structure and development in the monoecious palm Gaussia attenuate(Arecaceae;Arecoideae)[J]. Annals of Botany,114(7):1483-1495.
Jacquemard J C. 1995. Le palmier à huile. Series le technician dagriculture tropicale[M]. Paris: Editions Maisonneuve & Larose.
(責任編輯 蘭宗寶)
