Cd2+、Cu2+脅迫對黑藻(Hydrilla verticillata)的生長及光合熒光特性的影響
2016-05-23 05:55:24簡敏菲汪斯琛余厚平李玲玉簡美鋒余冠軍
生態學報 2016年6期
簡敏菲, 汪斯琛, 余厚平, 李玲玉, 簡美鋒, 余冠軍
1 江西師范大學,鄱陽湖濕地與流域研究教育部重點實驗室, 南昌 330022 2 江西師范大學生命科學學院,江西省亞熱帶植物資源保護與利用重點實驗室, 南昌 330022 3 江西鄱陽湖南磯山濕地國家級自然保護區管理局, 南昌 330009
?
Cd2+、Cu2+脅迫對黑藻(Hydrilla verticillata)的生長及光合熒光特性的影響
簡敏菲1,2,*, 汪斯琛2, 余厚平1, 李玲玉2, 簡美鋒2, 余冠軍3
1 江西師范大學,鄱陽湖濕地與流域研究教育部重點實驗室, 南昌3300222 江西師范大學生命科學學院,江西省亞熱帶植物資源保護與利用重點實驗室, 南昌3300223 江西鄱陽湖南磯山濕地國家級自然保護區管理局, 南昌330009
摘要:以黑藻(Hydrilla verticillata )為供試材料,采用Cd2+和Cu2+等兩種重金屬分別在5個濃度梯度水平下的河砂水培方法,研究Cd2+或Cu2+不同濃度脅迫對黑藻株高、生物量、成活率和葉綠素含量的影響,以及對黑藻葉片最小熒光(Fo)、最大熒光(Fm),PSⅡ最大量子產率(QYmax,)、穩態下的PSⅡ反應中心關閉程度(1-Qp_Lss)、穩態下的非光化學淬滅(NPQ_Lss)等光合熒光參數及其熒光成像的影響,探討各個參數分別隨鎘、銅濃度遞增的變化規律。研究結果表明,Cd2+脅迫下黑藻的株高顯著下降,說明Cd2+可能對黑藻葉片的維管束結構產生傷害作用;Cd2+和Cu2+脅迫對黑藻鮮重均無顯著影響,說明與水生植物自由水含量存在一定關系;Cd2+和Cu2+脅迫均使黑藻干重顯著下降,其中Cu2+脅迫對黑藻干重的影響更顯著,說明Cu2+脅迫對黑藻累積生物量的影響遠大于Cd2+;Cd2+和Cu2+脅迫下葉綠素各值均呈下降趨勢,而Cu2+脅迫對葉綠素的影響更大,說明Cu2+對黑藻葉綠體的毒害比Cd2+更大。隨著Cd2+或Cu2+脅迫濃度梯度的增加,黑藻的葉綠素熒光參數(Fo、Fm、QYmax)呈顯著下降趨勢,(1-Qp_Lss)呈上升趨勢,而NPQ_Lss先上升后下降。黑藻在不同重金屬脅迫下的生理指標、熒光參數及成像特征等方面所表現出的變化差異性,反映出同等濃度下黑藻受重金屬脅迫的影響程度為:Cd2+脅迫>Cu2+脅迫;黑藻可以在Cu2+濃度低于1 mg/L的環境下具有正常的光合活性,可推測將黑藻用于低濃度Cu污染水域的修復;在Cu2+濃度達3 mg/L以上環境下黑藻即無法長時間生存,可推測黑藻可以作為Cd污染水環境的指示種。
關鍵詞:黑藻; 重金屬脅迫; 生長活性; 葉綠素含量; 光合熒光參數; 熒光成像
全球工業化引起的重金屬污染,已成為人們關注的環境與社會突出問題[1- 2];水體中的重金屬污染問題也隨著冶煉、電鍍、采礦等各種工業廢水和固體廢物滲濾液的直接排入而日益突出[3]。重金屬污染物因不能自然降解或被微生物分解,易沉積到水域底部或在水中被水生生物吸收,一方面通過食物鏈積累而損害動物和人類健康[4],另一方面可影響植物的生長和植物體內活性氧代謝系統的平衡,導致一系列有害的生理生化變化[5]。鎘、銅均為水體重金屬污染物中最典型的污染元素[6],會對水生植物細胞造成極大破壞[7],極低濃度即可使細胞核仁解體、線粒體和葉綠體空泡化、高爾基體和內質網解體等,甚至導致植物無法正常生長[8]。沉水植物作為水生生態系統中最主要的初級生產者之一,不僅可以去除N、P等營養物質和有機污染負荷,對重金屬也具有一定的吸附和積累能力[9],對Cd和Cu等均具較強的富集作用,可用來去除水中的Cd、Cu[10],沉水植物的這種對重金屬的吸附、轉移或者將其固定在體內的富集特性可被運用于重金屬污染水域的治理[11],沉水植物在湖泊污染治理中的應用已日趨廣泛[12- 14]。近年來,國內外學者對于重金屬污染下沉水植物的生態毒理效應進行過研究,主要集中在植物對重金屬的富集作用[15- 18]、生理生化的傷害反應[19]、細胞內部亞顯微結構變化[20]等方面,但對于重金屬脅迫下黑藻等沉水植物的光合熒光和熒光成像等影響變化的研究尚較缺乏。研究鎘、銅對黑藻生理特性和光合熒光特性,可直接獲得鎘、銅污染脅迫下黑藻活體光合作用的變化,進而探究其耐受程度和脅迫機理,為水體重金屬污染的治理、恢復和重建水生植物群落提供實踐指導,對于研究沉水植物作為重金屬等各種污染物的指示生物等具有重要意義。
葉綠素熒光測定技術是一種簡便、快捷、可靠、無損傷的光合作用研究方法。自Kautsky和Hirsch[21]首次發現光合作用與葉綠素熒光間的聯系后[22],葉綠素熒光在植物的光合作用、脅迫生理學、水生生物學、海洋學和遙感等方面已得到廣泛的應用[23- 24]。脈沖幅度調制方式(PAM, Pulse Amplitude Modulation)下測量Kautsky效應,可以非常詳細地調查植物光合作用從暗適應狀態到光適應狀態,然后又到暗適應狀態的轉變。葉綠素熒光成像技術是1987年由Omasa等人開發[25],PAM技術于1996年由光子系統儀器首次介紹到葉綠素熒光成像[26],當實驗對象受到脅迫后(如受重金屬侵害),其熒光發射具有明顯異質性,事實證明采用葉綠素熒光成像技術已是必不可少[27]。目前,葉綠素熒光成像技術不僅用于光合作用的研究,還用于病毒和其他病原體局部監測(在可見癥狀出現之前)以及重金屬脅迫等方面[28]。葉綠素熒光成像可呈現出植物受到脅迫的熒光狀態,能更全面地了解植物的光合作用狀態,為詮釋其生長和耐受機制提供新的、直觀可見的手段。
黑藻(Hydrillaverticillata)是多年生大型淡水沉水植物,具有生長范圍廣、耐污能力和適應性強等特點,在國內外富營養化水體治理和生態修復中已得到廣泛應用,并對重金屬具有較強的吸收積累能力[29],可作為重金屬污染環境修復植物和水體污染的指示植物[30- 31]。黑藻對Cd、Cu兩種重金屬污染物均具有比較敏感的反應[32],超過一定濃度的Cd、Cu會抑制和傷害黑藻的生長。本文將通過不同濃度鎘、銅脅迫對黑藻株高、生物量、成活率和葉綠素含量的影響,以及對黑藻葉片光合熒光參數及其熒光成像的影響,結合前人的研究探討黑藻受脅迫后的生長和光合生理狀態,以及黑藻耐受和調節光能利用效率的內在機制,為修復和指示重金屬污染的生態環境、重建水生植物群落及修復湖泊生態系統提供科學依據。
1材料與方法
根據徐勤松等學者的研究結果,黑藻對Cd、Cu等兩種重金屬污染物具有比較敏感的反應,且Cd、Cu是重金屬污染水體中極常見的兩種元素,Cd、Cu對黑藻的毒害較大,致死濃度分別為0.5—1 mg/L和1—2 mg/L[7]。分別選取Cd、Cu等兩種重金屬元素作為水體重金屬脅迫污染物,選擇沉水植物黑藻(H.verticillata)作為供試植物材料,材料采自江西師范大學校園內湖水域。選取長勢相似的黑藻植株,截取生長狀況相似的10cm頂尖作為實驗材料[32]。
1.1材料培養與處理
2013年11月初將采回的黑藻放入盛有未經曝氣的自來水盆中(為防止綠藻滋生而嚴重影響黑藻生長,自來水不能曝氣),在室內馴養3—4 d后,采用培養盆(49 cm×36.5 cm×19 cm),在盆底鋪上洗凈的河砂約3.5 cm厚,加入等體積1/40 Hoagland植物培養液,每個培養盆內加培養液30 L,盆內水深15 cm,河沙約3.5 cm,將截取的頂尖10 cm的黑藻扦插于培養盆中,每個盆中扦插35株,2 d后分別于11月12日和28日8:00在培養盆中分別加入不同濃度的CdCl2溶液和CuSO4溶液進行處理,分別設置Cd2+和Cu2+的濃度梯度依次為0.5、1、3 mg/L和5 mg/L 等4個,另外設置0 mg/L Cd2+或Cu2+的對照培養組,相當于每種重金屬濃度梯度共有5個,每個濃度下分別設置3個重復組。培養盆均放置室內,采用空調控溫18—21 ℃,并采用LED燈補光,實際功率紅光∶藍光=3∶1(光照強度約為5000 lx),光周期為14 h∶10 h,水溫為17 ℃左右,第7天8:00 分別取植株進行分析。
1.2光照強度和溫度的測定
采用TES-1339專業照度計測定水面光照強度。自8:00到22:00每隔2h分別采用溫度計和水溫計測定1次空氣溫度和水溫。
1.3株高、生物量和成活率的測定
每組樣品加入不同濃度重金屬溶液培養7 d后,記錄每個培養盆中活體黑藻的株數,計算成活率;用直尺(精確到0.1 cm)測量15個培養盆中活體黑藻的株高;每個培養盆中取出10株黑藻,晾干表面水分,將用來墊著植物的兩層濾紙完全浸濕后,稱量其鮮重,于空氣中自然風干15 d后稱量其干重。
1.4植物葉綠素含量的測定
隨機選取培養盆中黑藻測量,獲得3個以上的平行數據。用100%二甲基亞砜(DMSO)分光光度法[33- 34]測定黑藻葉片的葉綠素含量,用DMSO在研缽中研磨植物葉片萃取,根據所得溶液在645 nm和663 nm的吸光值,運用Arnon公式[35]計算葉片的葉綠素總量(Ct)、葉綠素a (Ca)、葉綠素b (Cb)和Ca/Cb值。
Ca=12.7 A663- 2.69A645
(1)
Cb=22.9A645- 4.68A663
(2)
Ct=Ca+Cb= 8.02A645+ 20.21A663
(3)
1.5植株活體光合熒光參數的測定
每個培養盆中隨機選取黑藻測量,獲得3個以上的平行數據。運用FluorCam葉綠素熒光成像儀測定黑藻的熒光參數和活體熒光成像。取黑藻裝入廣口瓶中,包裹錫箔紙暗處理20 min,設定快門Shutter=0,敏感度Sensitivity=20,光照Act2=100,Act1=100,Super=70,測定熒光參數,包括暗適應狀態下的最小熒光(Fo)、暗適應狀態的最大熒光(Fm)、潛在光合活性(Fv/Fo,Potential photosynthetic activity)、最大PSⅡ量子產率(QYmax)、穩態下PSⅡ反應中心關閉程度(1-Qp_Lss)、穩態下的非光化學淬滅(NPQ_Lss)等指標及對應的植物熒光成像。
1.6數據統計分析方法
實驗結果均為3次以上平行數據的平均值和標準差,實驗數據采用統計軟件SPSS進行單因素方差分析(One-way, ANOVA)和最小顯著差異法(LSD),Pearson相關關系分析, 采用多重方差法分析顯著性,對黑藻在不同鎘、銅濃度下反應數據進行差異性比較;采用Origin 8.0進行作圖。
2結果與分析
2.1Cd2+、Cu2+脅迫對黑藻生長的影響
根據Cd2+或Cu2+污染脅迫中所設置的5個不同濃度梯度,測定黑藻生長中的株高、鮮重、干重以及成活率等數據,分別統計其變化特征及其相關顯著性,結果如圖1所示。

圖1 不同濃度重金屬脅迫下黑藻成活率、生物量和株高的變化及其相關顯著性Fig.1 Varieties and correlations of survival rate, biomass and plant heights of Hydrilla verticillata under the stress of different heavy metal concentrations圖中不同小寫字母表示不同處理之間達到Sig<0. 05水平的顯著差異,不同大寫字母表示不同處理之間達到Sig<0. 01水平的極顯著差異
圖1顯示,采用CdCl2和CuSO4溶液處理過的各培養組,Cd2+脅迫對株高的影響表現出顯著下降趨勢,濃度為3 mg/L 的Cd2+處理后的黑藻株高平均值最低,比對照組的降低6.72%,Cu2+脅迫對黑藻的株高無顯著影響。Cd2+和Cu2+脅迫對鮮重均無顯著影響,這可能與水生植物的自由水含量有關[36]。Cd2+和Cu2+脅迫對干重的影響均表現出顯著下降趨勢,其中Cu2+脅迫的影響更突出;濃度為5 mg/L 的Cu2+或Cd2+處理后的黑藻干重分別達到最低值,分別比對照組的干重降低41.31%和14.9%。0.5 mg/L Cd2+和Cu2+脅迫對成活率的影響均未表現出顯著差異,說明低濃度的Cd2+或Cu2+脅迫對黑藻的成活率影響不明顯,1 mg/L Cd2+脅迫下黑藻的成活率首先表現出顯著下降,不同Cd2+濃度梯度處理后黑藻的成活率分別比對照組的降低了2.02%、5.72%、16.16%和3.70%,而3 mg/L和5 mg/L Cu2+脅迫下黑藻的成活率急劇下降,不同Cu2+濃度梯度處理后黑藻的成活率分別比對照組的降低了0.97%、3.88%、67.96%和26.21%(圖1)。
2.2Cd2+、Cu2+脅迫對黑藻葉綠素的影響
根據Cd2+或Cu2+5個不同濃度梯度脅迫下黑藻的各種葉綠素含量測定數據,分別統計葉綠素總量(Ct)、葉綠素a(Ca)量、葉綠素b(Cb)量、葉綠素a/b(Ca/Cb)及其顯著相關性,結果如圖2所示。
圖2顯示,不同濃度Cu2+處理下的黑藻葉綠素各值均顯著下降,且同濃度的Cu2+處理比Cd2+處理后黑藻的葉綠素各值下降更明顯。Cu2+溶液處理過的培養組中,0.5 mg/L Cu2+處理下的葉綠素總量、葉綠素a、b等值均達到最低值,與對照組相比分別下降了48.7%、52.2%、39.4%;3 mg/L Cu2+處理后的黑藻Ca/Cb達到最低值,與對照組相比下降了45.2%。而不同濃度Cd2+處理過的培養組中,Cd2+濃度為5 mg/L處理下的黑藻葉綠素總量、葉綠素a、b、Ca/Cb等各值指標達最低值,與對照組相比分別下降24.1%、29.9%、5.5%、26.5%。0.5—3 mg/L濃度Cd2+脅迫下的黑藻葉綠素b呈現逐漸上升趨勢,但Cd2+濃度為5 mg/L脅迫下黑藻的葉綠素b呈現下降(圖2)。
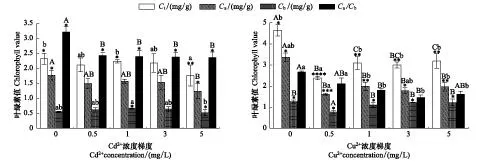
圖2 不同濃度重金屬脅迫下黑藻葉綠素總量(Tc),葉綠素a(Ca),葉綠素b(Cb)和Ca/Cb及其相關性Fig.2 Varieties and correlations of total chlorophyll (Tc), chlorophyll a (Ca), chlorophyll b (Cb) and Ca/Cb of Hydrilla verticillata under the stress of different heavy metal concentrations圖中不同小寫字母表示不同處理之間達到Sig<0. 05水平的顯著差異,不同大寫字母表示不同處理之間達到Sig<0. 01水平的極顯著差異
2.3Cd2+、Cu2+脅迫對黑藻葉綠素熒光參數的影響
分別測定上述5個不同濃度梯度Cd2+或Cu2+脅迫處理一周后黑藻的各種熒光參數:暗適應狀態的最小熒光Fo和最大熒光Fm、潛在光合活性FV/Fo、最大PSⅡ量子產率QYmax、穩態下PSⅡ反應中心關閉程度(1-Qp_Lss)、穩態下的非光化學淬滅NPQ_Lss等指標,分析不同濃度重金屬脅迫對黑藻葉綠素熒光各參數的影響,結果如圖3或表1所示。
圖3結果顯示,Cd2+或Cu2+脅迫處理后,黑藻的Fo、Fm、QYmax和Fv/Fo均呈下降趨勢。Cd2+脅迫對黑藻葉片的Fo有顯著影響,而Cu2+脅迫則無顯著影響。在濃度為0.5 mg/L,Cd2+或Cu2+脅迫處理后黑藻的Fm分別顯著下降43.0%和17.9%,Cd2+脅迫后黑藻的Fm下降更突出(圖3)。Cd2+或Cu2+脅迫下黑藻葉片的Fv/Fo均呈顯著下降,受Cd2+脅迫后的效果相對非常突出,5 mg/L Cd2+脅迫下黑藻葉片的Fv/Fo最低,比對照組下降了90.4%,而5 mg/L Cu2+脅迫下黑藻葉片的Fv/Fo也最低,但比其對照組只下降24.5%。同樣,Cd2+或Cu2+處理后黑藻葉片的QYmax均呈顯著下降,5 mg/L Cd2+脅迫下黑藻葉片的QYmax最低,下降25.2%,而5 mg/L Cu2+脅迫下黑藻葉片的QYmax也最低,下降8.5%(圖3)。黑藻葉片QYmax和Fv/Fo值的下降表明,重金屬離子使PSⅡ反應中心受損,抑制光合作用的原初反應,阻礙光合電子的傳遞過程。圖3中Cd、Cu對黑藻葉片熒光參數的影響均是先顯著降低,之后隨著重金屬濃度的增加有少量回升,最后呈現較平穩趨勢。反映出重金屬對黑藻的脅迫較明顯,且隨著重金屬濃度的上升而上升,而黑藻體內光合機制的改變,使得黑藻能在重金屬環境下仍然保持光合活性。
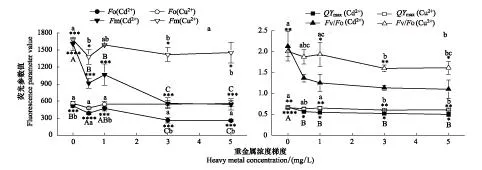
圖3 不同濃度重金屬脅迫對黑藻葉綠素熒光參數的影響Fig.3 Influences on chlorophyll fluorescence parameters of Hydrilla verticillata under the stress of different heavy metal concentrations不同小寫字母表示不同處理之間達到Sig<0.05水平的顯著差異,不同大寫字母表示不同處理之間達到Sig<0.01水平的極顯著差異
表1顯示,與空白對照組相比,Cd2+或Cu2+脅迫后的黑藻葉片穩態下PSⅡ反應中心關閉程度(1-Qp_Lss)、非光化學淬滅系數(NPQ_Lss)上升,表明Cd2+或Cu2+脅迫改變了黑藻葉片的光能利用分配。
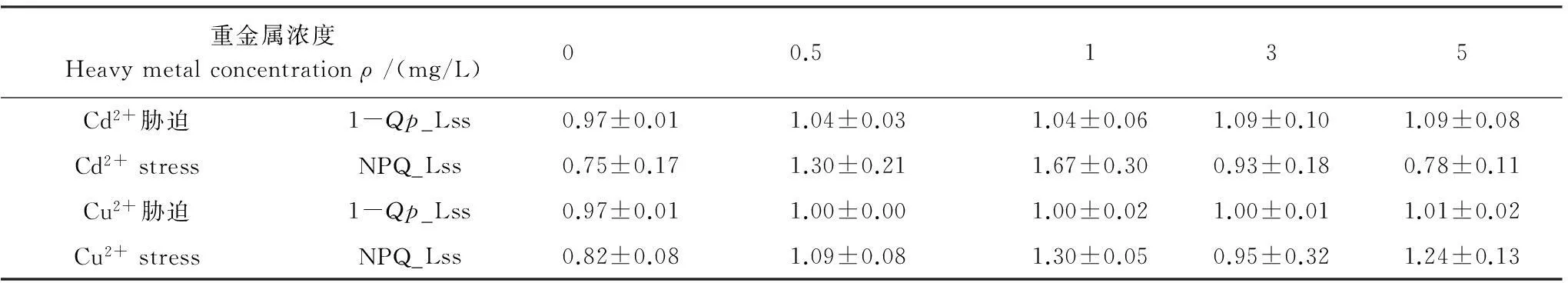
表1 不同濃度重金屬脅迫對黑藻(1-Qp_Lss)和NPQ_Lss的影響
2.4Cd2+、Cu2+脅迫對黑藻熒光成像的影響
Cd2+或Cu2+不同濃度梯度脅迫處理后的黑藻葉片PSⅡ最大量子產率時的熒光成像分別如圖4。結果顯示,Cd2+脅迫下的黑藻葉片熒光成像強度由高變低,濃度在3 mg/L及以上的Cd2+脅迫處理后,黑藻葉片大部分失去光合作用活性,只有莖稈(黑藻葉片中間為少量莖稈橫截面)附近的少量區域有光合活性(圖4)。而濃度為0.5 mg/L和1 mg/L的Cu2+脅迫對黑藻葉片光合活性的影響并不顯著,但隨著Cu濃度的再升高呈顯著下降趨勢(圖4)。另一方面,Cd2+或Cu2+對照組中的黑藻能運用整個葉面進行光合作用,而受Cd2+、Cu2+脅迫后的黑藻葉片光合活性降低,且較高的光合活性位于莖稈處和葉脈處,Cd2+、Cu2+脅迫對黑藻葉片光合活性的傷害從葉片邊緣開始達到最嚴重,而莖稈和葉脈處最輕,這可能與莖和葉脈的維管束結構有一定關聯性。

圖4 不同濃度重金屬脅迫對黑藻葉綠素熒光(QY_max)成像的影響Fig.4 Varieties of chlorophyll fluorescence (QY_max) imagings under the stress of different heavy metal concentrations
3結論與討論
葉綠素是與植物光合作用最密切相關的一種色素,Cd2+、Cu2+脅迫分區均造成黑藻葉綠素含量下降,黑藻在Cu2+脅迫下的葉綠素含量比Cd2+脅迫下的減少更多,說明Cu2+脅迫比Cd2+脅迫對葉綠體的毒害作用更大,可能原因是Cu2+脅迫對內膜、類囊體片層和光合色素等的損害更嚴重[32]。當用0.5—3 mg/L Cd2+濃度脅迫處理黑藻時,葉綠素總量下降并不十分明顯,當Cd2+濃度達到5 mg/L時葉綠素總量下降最多,但是在0.5 mg/L Cd2+脅迫下的葉綠素a/b(Ca/Cb)會立刻顯著下降,后隨著Cd2+濃度的升高并未產生顯著下降。黑藻的葉綠素b在Cd2+濃度為0.5—3 mg/L間的脅迫下有所上升,而Cd2+濃度為5 mg/L時出現下降,有研究認為葉綠素b含量的減少會影響到光系統的穩定性,最終導致整個光合機構的不穩定,具體表現在對逆境的耐受性降低[37- 38]。反之推測,Cd2+脅迫下黑藻葉綠素b含量增加,可能是黑藻對Cd2+脅迫耐受能力的增強,Cu2+脅迫下黑藻葉綠素b也在0.5 mg/L下降到最低后上升,與黑藻對這兩種金屬的耐受機制有關。以上說明在Cd2+脅迫導致黑藻葉綠素含量減少的情況下,黑藻通過調整葉綠素Ca/Cb的比值以適應逆境[39],具體機制尚需進一步探究。
葉綠素a熒光動力學在植物生理領域得到廣泛應用,QYmax反映了 PSⅡ反應中心光能轉化效率,可作為植物對不良環境耐受性等方面的重要指標[40]。Cd2+脅迫的黑藻Fm比Cu脅迫下降得明顯更多,Fo下降的差別較小,因此,Cd2+脅迫下的QYmax值也比Cu2+脅迫下的降低得更多,說明黑藻光合活性能力對重金屬的耐受能力:Cu2+耐受>Cd2+耐受。研究表明在對黑藻細胞的損害中,Cd2+對葉綠體的破壞較小[8],但Cd2+脅迫下黑藻的潛在光合活性(Fv/Fo表示PSⅡ的潛在活性)比受Cu2+脅迫后下降得更多,反映出Cd2+對黑藻內部光合作用結構的破壞比Cu2+更徹底,并且Cd2+、Cu2+脅迫對黑藻光合活性的傷害是不可逆的[8,10]。1-Qp_Lss是 PSⅡ反應中心關閉的程度,在一定程度上反映 QA的氧化還原狀態,1-Qp_Lss越大,PSⅡ反應活性越小。與對照組相比,Cd、Cu脅迫后的黑藻葉片1-Qp_Lss 增加表明電子由PSⅡ的氧化側向PSⅡ反應中心的傳遞受阻,改變了黑藻葉片的光光能利用分配,將原本用來獲得量子產量的部分光能熱耗散等用于增強自我保護,這有可能是造成非光化學淬滅系數(NPQ_Lss)上升和PSⅡ最大量子產率(QYmax)下降的原因,之后非光化學淬滅減小的原因可能在于PSⅡ的部分失活,黑藻的正常生理功能被嚴重破壞。
Govindjee和Nedbal用“百聞不如一見”來表達并強調葉綠素熒光成像如何獲得科學實驗的信念[27]。QYmax表示植物PSⅡ最大量子產率,其葉綠素熒光成像最能反映重金屬脅迫對光系統Ⅱ的損害,熒光成像表明Cd2+、Cu2+重金屬脅迫使黑藻葉片PSⅡ反應中心受到嚴重損害,光合活性減弱。Cd2+、Cu2+對黑藻葉片的損害都是從邊緣開始的,黑藻葉片是由單層細胞組成,Cd2+、Cu2+對黑藻的毒害是對植物細胞的膜結構和非膜結構、生理活動和生化反應的整體傷害[8,10],但是葉脈、莖和葉片結構不同,因此推測維管束結構更難被Cd2+、Cu2+破壞。
從植物形態上可以判斷植物的生長和受脅迫情況,Cd2+脅迫下黑藻株高比Cu2+脅迫下的下降率更高,說明Cd2+脅迫對黑藻株高的影響大于Cu2+脅迫對黑藻株高的影響;3 mg/L 的Cu2+脅迫對黑藻的成活率影響最大,隨著濃度的升高,黑藻的成活率反而有所上升,Cd2+脅迫的下降率更低;Cu2+脅迫下黑藻干重比Cd2+脅迫下降率更高。在3 mg/L Cu2+脅迫下,黑藻成活率、Ca/Cb、QYmax最低;5 mg/L Cu2+脅迫下株高、鮮重、干重最低,表明5 mg/L Cu2+脅迫下黑藻長勢差,有機物和無機物含量減少,葉綠素增加,Ca增加而Cb減少了。圖2中可以看出,Cu2+脅迫下葉綠素b只在0.5 mg/L時下降顯著,其他濃度的脅迫和對照組都沒有顯著差異,說明Cu2+脅迫對黑藻葉綠素a的影響比葉綠素b大,可能的原因有:Cu2+促進葉綠素的生物合成[41],或者破壞葉綠素酶活性,使葉綠素降解減少[42],葉綠素a的主要功能是吸收和傳遞光能,加強黑藻的光合作用能力,因此最大量子產率(QYmax)也增加,光合作用的增強對黑藻的成活率有較大的影響。圖2中5 mg/L Cd2+脅迫下與3 mg/L Cd2+脅迫下相比可以看出,黑藻的葉綠素a和葉綠素b均呈顯著下降,但是光合能力指標Ca/Cb并無顯著差異,說明光合作用能力并未下降,圖3中的量子最大產率QYmax無顯著差異也印證了這一點。光合作用無明顯差異的情況下,有機物和無機物的累積也無明顯變化,干重無顯著差異。而5 mg/L Cd2+脅迫下與3 mg/L Cd2+脅迫下相比,表1中NPQ_Lss的降低說明黑藻的光保護能力下降,圖4可以看出,黑藻葉片的成活面積變小,活體細胞大多集中于中間葉脈部分,光合作用得到的有機物用來維持活體細胞的生長,即很大一部分用于葉脈的生長,葉脈的生長使得株高增加,而由于葉脈比葉片更難被Cd2+破壞,因此成活率得以上升。
Cd2+是黑藻的非必須營養元素,研究表明,Cd2+使黑藻體內活性氧減少、影響細胞壁的形成、破壞細胞器和細胞、損傷類囊體[8],Cd2+脅迫下黑藻光合活性顯著下降,3 mg/L及以上濃度的Cd2+對黑藻的傷害極大,黑藻因此會失去大部分光合能力,無法長時間生存,表明黑藻可作為Cd污染水環境的指示種。Cu2+是黑藻生長必須的微量營養元素,所以一定濃度的Cu2+對黑藻的生長是有利的,但是超過一定范圍就會造成黑藻細胞核消解、線粒體空泡化、類囊體片層排列混亂甚至溶解、葉綠體膨脹成球形然后解體、蛋白質和脂類失活等損害[32]。本研究中,濃度為0.5和1 mg/L 的Cu2+脅迫對黑藻光合活性的抑制不顯著,表明在低濃度的Cu2+環境中黑藻可以進行比較正常的生理活動,可推測將黑藻用于低濃度Cu2+污染水域的修復與水生生態系統的沉水植物重建。本研究得出的結論能否用于實踐,尚需要更多研究來確定其可靠性和實用性。
參考文獻(References):
[1]Khan K S, Huang C Y. Effects of heavy metal pollution on soil microbial biomass. Journal of Environmental Sciences, 1999, 11(1): 40- 47.
[2]Yang X, R?mheld V, Marschner H, Chaney R L. Application of chelator-buffered nutrient solution technique in studies on zinc nutrition in rice plant (OryzasativaL.). Plant and Soil, 1994, 163(1): 85- 94.
[3]刁維萍, 倪吾鐘, 倪天華, 楊肖娥. 水體重金屬污染的生態效應與防治對策. 廣東微量元素科學, 2003, 10(3): 1- 5.
[4]黃永杰, 劉登義, 王友保, 王興明, 李晶. 八種水生植物對重金屬富集能力的比較研究. 生態學雜志, 2006, 25(5): 541- 545.
[5]顏昌宙, 曾阿妍. 沉水植物對重金屬Cu2+的生物吸附及其生理反應. 農業環境科學學報, 2009, 28(2): 366- 370.
[6]楊艷華, 陳國祥, 劉少華, 王娜, 吳國榮, 施國新. 鎘對黑藻葉光化學及硝酸還原酶特性的影響. 南京師大學報: 自然科學版, 2002, 25(1): 28- 32.
[7]徐勤松, 施國新, 周耀明, 吳國榮, 王學. 鎘在黑藻葉細胞中的亞顯微定位分布及毒害效應分析. 實驗生物學報, 2004, 37(6): 461- 468.
[8]施國新, 杜開和, 解凱彬, 丁小余, 常福辰, 陳國祥. 汞、鎘污染對黑藻葉細胞傷害的超微結構研究. 植物學報, 2000, 42(4): 373- 378.
[9]馬劍敏, 靳萍, 吳振斌. 沉水植物對重金屬的吸收凈化和受害機理研究進展. 植物學通報, 2007, 24(2): 232- 239.
[10]徐勤松, 施國新, 許丙軍, 吳國榮, 胡金朝. Cu、Zn在黑藻葉片中的富集及其毒理學分析. 水生生物學報, 2007, 31(1): 1- 8.
[11]賀鋒, 吳振斌. 水生植物在污水處理和水質改善中的應用. 植物學通報, 2003, 20(6): 641- 647.
[12]Qiu D R, Wu Z B, Liu B Y, Deng J Q, Fu G P, He F. The restoration of aquatic macrophytes for improving water quality in a hypertrophic shallow lake in Hubei Province, China. Ecological Engineering, 2001, 18(2): 147- 156.
[13]Wang G X, Zhang L M, Chua H, Li X D, Xia M F, Pu P M. A mosaic community of macrophytes for the ecological remediation of eutrophic shallow lakes. Ecological Engineering, 2009, 35(4): 582- 590.
[14]Chojnacka K, Chojnacki A, Górecka H. Biosorption of Cr3+, Cd2+and Cu2+ions by blue-green algaeSpirulinasp.: kinetics, equilibrium and the mechanism of the process. Chemosphere, 2005, 59(1): 75- 84.
[15]潘義宏, 王宏鑌, 谷兆萍, 熊國煥, 易鋒. 大型水生植物對重金屬的富集與轉移. 生態學報, 2010, 30(23): 6430- 6441.
[16]Visoottiviseth P, Francesconi K, Sridokchan W. The potential of Thai indigenous plant species for the phytoremediation of arsenic contaminated land. Environmental Pollution, 2002, 118(3): 453- 461.
[17]Zhao F J, Ma J F, Meharg A A, McGrath S P. Arsenic uptake and metabolism in plants. New Phytologist, 2008, 181(4): 777- 794.
[18]周守標, 王春景, 楊海軍, 畢德, 李金花, 王影. 菰和菖蒲對重金屬的脅迫反應及其富集能力. 生態學報, 2007, 27(1): 281- 287.
[19]閔海麗, 蔡三娟, 徐勤松, 施國新. 外源鈣對黑藻抗鎘脅迫能力的影響. 生態學報, 2012, 32(1): 256- 264.
[20]Aravind P, Prasad M N V. Modulation of cadmium-induced oxidative stress inCeratophyllumdemersumby zinc involves ascorbate -glutathione cycle and glutathione metabolism. Plant Physiology and Biochemistry, 2005, 43(2): 107- 116.
[21]Kautsky H, Hirsch A. Neue versuche zur Kohlens?ureassimilation. Naturwissenschaften, 1931, 19(48): 964- 964.
[22]Lin S Q, Xu C H, Zhang Q D, Xu L, Mao D Z, Kuang T Y. Some application of chlorophyll fluorescence kinetics to plants plant stress physiologyphy to ecology and agricultural modernization. Chinese Bulletin of Botany, 1992, 9(1): 1- 16.
[23]Krause G H, Wei E. Chlorophyll fluorescence and photosynthesis: The basics. Annual Review of Plant Physiology and Plant Molecular Biology, 1991, 42(1): 313- 349.
[24]Fracheboud Y, Haldimann P, Leipner J, Stamp P. Chlorophyll fluorescence as a selection tool for cold tolerance of photosynthesis in maize (ZeamaysL.). Journal of Experimental Botany, 1999, 50(338): 1533- 1540.
[25]Omasa K, Shimazaki K I, Aiga I, Larcher W, Onoe M. Image analysis of chlorophyll fluorescence transients for diagnosing the photosynthetic system of attached leaves. Plant Physiology, 1987, 84(3): 748- 752.
[26]Nedbal L, Soukupova J, Kaftan D, Whitmarsh J, Trtilek M. Kinetic imaging of chlorophyll fluorescence using modulated light. Photosynthesis Research, 2000, 66(1/2): 3- 12.
[27]Govindjee N L. Seeing is believing. Photosynthetica, 2000, 38(4): 481- 482.
[28]Balachandran S, Osmond C B, Daley P F. Diagnosis of the earliest strain-specific interactions between tobacco mosaic virus and chloroplasts of tobacco leaves in vivo by means of chlorophyll fluorescence imaging. Plant Physiology, 1994, 104(3): 1059- 1065.
[29]黃亮, 李偉, 吳瑩, 張經, 周菊珍. 長江中游若干湖泊中水生植物體內重金屬分布. 環境科學研究, 2002, 15(6): 1- 4.
[30]Wang C, Zhang S H, Wang P F, Li W, Lu J. Effects of ammonium on the antioxidative response inHydrillaverticillata(L. f.) Royle plants. Ecotoxicology and Environmental Safety, 2010, 73(2): 189- 195.
[31]Yan S, Zhou Q X. Toxic effects ofHydrillaverticillataexposed to toluene, ethylbenzene and xylene and safety assessment for protecting aquatic macrophytes. Chemosphere, 2011, 85(6): 1088- 1094.
[32]徐勤松, 施國新, 王學, 吳國榮. 鎘、銅和鋅脅迫下黑藻活性氧的產生及抗氧化酶活性的變化研究. 水生生物學報, 2006, 30(1): 107- 112.
[33]陳振德, 傅以彬, 鄒琦, 程炳嵩, 梁作勤. 二甲亞砜和丙酮混合法測定葉綠素含量. 山東農業大學學報, 1989, (2): 31- 35.
[34]陳元, 趙洋甬, 潘雙葉, 徐運, 蔣蕾蕾. Phyto-PAM對浮游植物中葉綠素的分類測定. 現代科學儀器, 2009, (4): 100- 103.
[35]葉濟宇. 關于葉綠素含量測定中的Arnon計算公式. 植物生理學通訊, 1985, (6): 69- 69.
[36]肖志華, 張義賢, 張喜文, 李萍. 外源鉛、銅脅迫對不同基因型谷子幼苗生理生態特性的影響. 生態學報, 2012, 32(3): 889- 897.
[37]Chu H A, Nguyen A P, Debus R J. Site-directed photosystem Ⅱ mutants with perturbed oxygen-evolving properties. 1. instability or inefficient assembly of the manganese clusters in vivo. Biochemistry, 1994, 33(20): 6137- 6149.
[38]Havaux M, Tardy F. Thermostability and photostability of photosystem Ⅱ in leaves of the Chlorina-f2 barley mutant deficient in light-harvesting chlorophyll a/b protein complexes. Plant Physiology, 1997, 113(3): 913- 923.
[39]Rüdiger W. Biosynthesis of chlorophyllband the chlorophyll cycle. Photosynthesis Research, 2002, 74(2): 187- 193.
[40]張守仁. 葉綠素熒光動力學參數的意義及討論. 植物學通報, 1999, 16(4): 444- 448.
[41]孫賽初, 王煥校, 李啟任. 水生維管束植物受鎘污染后的生理變化及受害機制初探. 植物生理學報, 1985, 11(2): 113- 121.
[42]楊丹慧. 重金屬離子對高等植物光合膜結構與功能的影響. 植物學通報, 1991, 8(3): 26- 29.
InfluenceofCd2+orCu2+stressonthegrowthandphotosyntheticfluorescencecharacteristicsofHydrilla verticillata
JIANMinfei1,2,*,WANGSichen2,YUHouping1,LILingyu2,JIANMeifeng2,YUGuanjun3
1 Key Laboratory of Poyang Lake Wetland and Watershed Research ,Ministry of Education, Jiangxi Normal University, Nanchang 330022, China 2 College of Life Sciences, Jiangxi Normal University, Jiangxi Provincial Key Laboratory of Protection and Utilization of Subtropical Plant Resources, Nanchang 330022, China 3JiangxiNanjiWetlandNationalNatureReserveBureau,Nanchang330009,China
Abstract:In this study, we selected the aquatic plant Hydrilla verticillata as our specimen and cultured it in clean river sand and water in order to study the influence of five separate concentration levels of Cd2+or Cu2+on the plant. Effects of the stress on the plant were identified, and we recorded parameters such as height, biomass, chlorophyll content, and survival ratios of H. verticillata, as well as the effects of fluorescence parameters such as minimal fluorescence (Fo), maximal fluorescence (Fm), PS II maximum quantum yield (QYmax), degree of PSⅡ reaction centers closed under stable state (1-Qp_Lss), non-photochemical quenching under stable state (NPQ_Lss), and fluorescence images of H. verticillata leaves. We also separately studied the variation characteristics of each parameter of H. verticillata under the stress of increasing Cd2+or Cu2+concentrations. We found that the height of H. verticillata decreased significantly under the different levels of pollution stress by the heavy metal Cd2+. This indicated that Cd2+pollution may damage the bundle structure of H. verticillata. There were no significant effects on the fresh weight of H. verticillata under the stress of either Cd2+or Cu2+pollution, and this may be related to the high content of free water in aquatic plants. Dry weight of H. verticillata decreased significantly when the plants were exposed to Cd2+or Cu2+pollution. Effect of Cu2+pollution on the dry weight of H. verticillata was far more prominent than that of Cd2+pollution. All chlorophyll values decreased significantly when the plants were subjected to Cd2+or Cu2+pollution, and we found that the effect of Cu2+pollution was greater than that of Cd2+pollution. This suggested that the toxicity of Cu2+against H. verticillata chloroplasts is greater than that of Cd2+. The chlorophyll fluorescence parameters (Fo, Fm, and QYmax) of H. verticillata under Cd2+or Cd2+stress were significantly lower than those of the control groups; however, the values of (1-Qp_Lss) increased, and the values of NPQ_Lss first increased and then decreased as the concentrations of the heavy metals increased. The different values for the physiological parameters, fluorescence parameters, and imaging characteristics indicated that the influence of Cd2+on H. verticillata was greater than that of Cu2+when the same concentrations of the two different heavy metals were used. When Cu2+concentration was lower than 1 mg/L in the water environment, photosynthetic activities of H. verticillata under Cu2+stress were almost normal. This means that H. verticillata could be used to restore a water environment with a low Cu2+concentration. H. verticillata cannot survive for a long time in a water environment contaminated with Cd2+or if Cu2+concentrations reach or exceed 3 mg/L. This means that H. verticillata could be used as an indicator species for water contaminated with Cd2+.
Key Words:Hydrilla verticillata; heavy metal stress; growth activity; chlorophyll content; photosynthetic fluorescence parameters; fluorescence imaging
DOI:10.5846/stxb201408081588
*通訊作者
Corresponding author.E-mail: jianminfei0914@163.com
收稿日期:2014- 08- 08; 網絡出版日期:2015- 05- 20
基金項目:國家自然科學基金資助項目(41161035, 41461042);國家科技部“十二五”科技支撐計劃資助項目(2011BAC13B02); 江西省重大生態安全問題監控協同創新中心資助項目(JXS-EW-03); 江西省科技支撐計劃項目(20133BBG70005); 江西省亞熱帶植物資源保護與利用重點實驗室開放基金資助項目(YRD201314)
簡敏菲, 汪斯琛, 余厚平, 李玲玉, 簡美鋒, 余冠軍.Cd2+、Cu2+脅迫對黑藻(Hydrillaverticillata)的生長及光合熒光特性的影響.生態學報,2016,36(6):1719- 1727.
Jian M F, Wang S C, Yu H P, Li L Y, Jian M F, Yu G J.Influence of Cd2+or Cu2+stress on the growth and photosynthetic fluorescence characteristics ofHydrillaverticillata.ActaEcologicaSinica,2016,36(6):1719- 1727.
