蘿卜隱性核雄性不育系136A的選育及其育性研究
2016-05-14 17:56:29白靜段乃彬謝坤王俊峰王效睦
山東農業科學 2016年7期
白靜 段乃彬 謝坤 王俊峰 王效睦

摘要:本研究從自然群體中發現蘿卜核雄性不育材料136A,通過創制178個雜交組合,經多代雜交,選育出具有50%保持能力的保持系136B。田間遺傳試驗結果表明,136A不育性狀表現穩定,呈現出由隱性單基因控制的核不育遺傳特點。細胞學觀察顯示該材料花粉敗育徹底,花粉發育在四分體后開始發生異常,花粉敗育主要由花粉壁破裂造成。PCR結果顯示,該材料中不含有Ogura 雄性不育類型的主效基因orf138;線粒體和葉綠體測序結果也顯示該材料具有正常的可育細胞質。
關鍵詞:蘿卜;核雄性不育;136A;保持系
中圖分類號:S631.101文獻標識號:A文章編號:1001-4942(2016)07-0026-06
蘿卜是典型的異花授粉作物,雜種優勢非常明顯,利用雄性不育系進行雜交育種可以得到純度高、性狀好的雜交種,且制種過程簡單,成本低[1]。雄性不育根據不育基因的遺傳方式和在細胞中的定位分為細胞核雄性不育和細胞質雄性不育。目前生產上利用最多的蘿卜不育系為Ogura類型的細胞質雄性不育系,它不僅在蘿卜上使用,還被轉育到大白菜、小白菜、甘藍、油菜、菜花等其他十字花科作物中。單一化的雄性不育種質導致雜交種綜合性狀難以得到突破性的提高,對外界有害生物與環境的抵抗力非常低,存在潛在的生產風險,如1970年美國大面積種植T型細胞質的玉米雜交種,因玉米小斑病T小種的爆發流行,導致玉米產業遭受嚴重的經濟損失。
細胞核雄性不育 (GMS)由細胞核不育基因控制,不受細胞質影響,沒有正、反交遺傳效應[2,3]。相對于細胞質雄性不育,核不育系具有育性穩定、恢復源廣的優點。核不育現象十分普遍,已在294 種植物和21個種屬間雜交組合中發現由細胞核基因控制的雄性不育類型[2,4,5]。目前,在十字花科植物中,細胞核雄性不育已經被應用在油菜、白菜、甘藍等作物中。
相對于細胞質雄性不育,核雄性不育的保持系只有50%的保持率,在生產上很難直接利用,但通過不斷發現新的核不育基因同樣得到了保持率100%的保持系。陳鳳祥等在甘藍型油菜中發現一類新的細胞核雄性不育材料9012A受三對核基因控制,分別為兩對隱性重疊不育基因和一對隱性上位可育基因[6],并選育出具有100%保持能力的保持系。張書芳等在大白菜中利用核隱性基因和顯性上位基因育出了能夠100%保持的核雄性不育保持系[7]。通過類似的方法,科研工作者先后育成一些品種應用于生產[8~12]。隨著生物技術的發展,核雄性不育材料具有更廣闊的應用前景。美國杜邦先鋒公司發明了一項雜交種子生產技術(Seed production technology,SPT),這種新的基因工程手段能夠充分利用隱性核不育突變材料,將花粉育性恢復基因、花粉致死基因和標記篩選基因緊密連鎖并導入核雄性不育突變體中,從而獲得相應的保持系,有效解決了隱性核雄性不育系的繁殖難題,同時該方法還解決了產品的轉基因問題,為隱性核不育材料的利用提供了新的模式[13]。
然而,在蘿卜雄性不育系培育中,國內尚未有細胞核雄性不育材料的報道。本研究利用從自然群體中發現的蘿卜雄性不育株為供試材料,通過田間育性遺傳統計試驗證明新發現的雄性不育材料為由一對隱性基因控制的核不育材料,填補了蘿卜雄性不育資源中缺乏核不育材料的空白,為蘿卜雄性不育系的培育與研究提供了重要的不育源。通過細胞學觀察初步確定花粉敗育時期及不同類型不育系花粉敗育的差異。
1材料與方法
1.1試驗材料
蘿卜Ogura雄性不育系RS43,蘿卜Ogura雄性不育保持系RS45及50余份國內外蘿卜種質材料。
1.2試驗方法
1.2.1蘿卜核不育系的選育與遺傳學驗證2012年春于自然群體中發現1株不育株,命名為136A,以同一自然群體中的可育株作父本進行雜交,雜交組合31個;同時與5個不同品種的可育株進行雜交,雜交組合147個。每個組合后代F1及父本自交種子于當年秋季種于田間,調查育性。從中選出可以對不育材料136A保持的可育株為保持系136B。雄性不育株的標準為,雌蕊發育正常,雄蕊完全退化或不能形成有效花粉。
將不育材料與保持系的后代多代連續姐妹交,統計育性分離情況;將可育株自交,統計后代育性分離情況。
1.2.2花藥發育細胞學觀察以136A、RS43、136B的50個花蕾為試驗材料。從現蕾期開始選取同一花序上不同發育時期的花蕾(從1 mm大小開始,直至開放前的花蕾),每個花蕾取一枚花藥放于載玻片上,用鑷子將花藥研碎,用濃度為1%的醋酸洋紅染液浸泡花藥進行染色,蓋上蓋玻片,于顯微鏡(Leica DM5000 B)下分別觀察3個材料不同時期的花藥發育情況,并拍照。
1.2.3PCR驗證利用CTAB法[14]分別提取RS44(即136A)、RS43(具有Ogura細胞質,陽性對照)、RS45(具有正常可育細胞質,陰性對照)的全基因組DNA,以Ogura類型主效不育基因orf138的特異引物(ORF-F:5′-TTCAAATCCTGTCCCCGCACC-3′和ORF-R:5′-GCCTTACACCATTGGGATACTTC-3′)對3個材料進行PCR擴增,凝膠電泳檢測136A中是否含有orf138基因。
1.2.4葉綠體和線粒體測序分別以RS43和RS45為陽性和陰性對照,對RS43、RS45和136A 3個材料進行線粒體和葉綠體基因組DNA測序、組裝。
為保證樣品純度,防止出現混雜現象,在提取線粒體和葉綠體DNA過程中,分別從某一單株提取樣品。根據線粒體和葉綠體DNA的提取方法[15],提取含有線粒體和葉綠體的混合DNA樣品,進行高通量測序。葉綠體測序深度在600倍以上,線粒體測序深度在100倍以上。根據測序深度的不同,經過De novo程序組裝及PCR查補間隙,先組裝葉綠體。De novo組裝的線粒體Contigs先與葉綠體DNA序列進行比對,剔除相同部分,再進行線粒體組裝,組裝拼接方法與葉綠體相同,最后再用PCR的方法,將葉綠體與線粒體相同的DNA序列進行甄別。
2結果與分析
2.1蘿卜核不育系與保持系的選育
與136A進行雜交的178個組合中,2012年秋調查發現有一個組合的雜種F1出現1∶1的育性分離,即對136A不育系具有50%的保持能力,將其命名為136B。136B與136A來自同一自然群體,其他所有組合的雜種一代均表現為可育。將136B自交獲得的50粒種子種于田間觀察育性,結果為自交后代中有37株可育株,13株不育株,分離比例約為3∶1。將136A與136B雜交一代中的可育株與不育株雜交,收獲種子FA2于來年種植觀察育性分離情況。2013年春 136A與136B雜交后代中的不育株與可育株雜交后代FA2的育性分離比約為1∶1。
將136A與136B的雜交后代連續6代姐妹交,最終基本形成了純合、穩定的蘿卜核不育AB系,選育過程如圖1。
2.2136A基因型的遺傳學驗證
以具有正常細胞質的可育株RS45為母本,以雜交后代中的可育株(FB1)為父本進行雜交,其后代BF1全部可育。取20粒BF1種子種植并自交得到BF2,對20份BF2每份取50粒種子進行春化處理后種于溫室中,調查育性分離情況。其中,11份BF2代種子表現為全部可育,9份BF2代種子出現育性分離,兩者比例約為1∶1。9份出現育性分離的BF2分離情況如表1,合計可育株330株,不育株108株,分離比例符合3∶1比率,即符合隱性單基因的遺傳規律。與136B自交試驗相比,該試驗中BF2的細胞質由136B的細胞質更換為RS45的細胞質,但育性分離結果一致,表明該材料育性分離情況不受細胞質的影響,同時驗證了該雄性不育為核基因控制的隱性單基因性狀。
2.3136A花粉敗育的生物學觀察
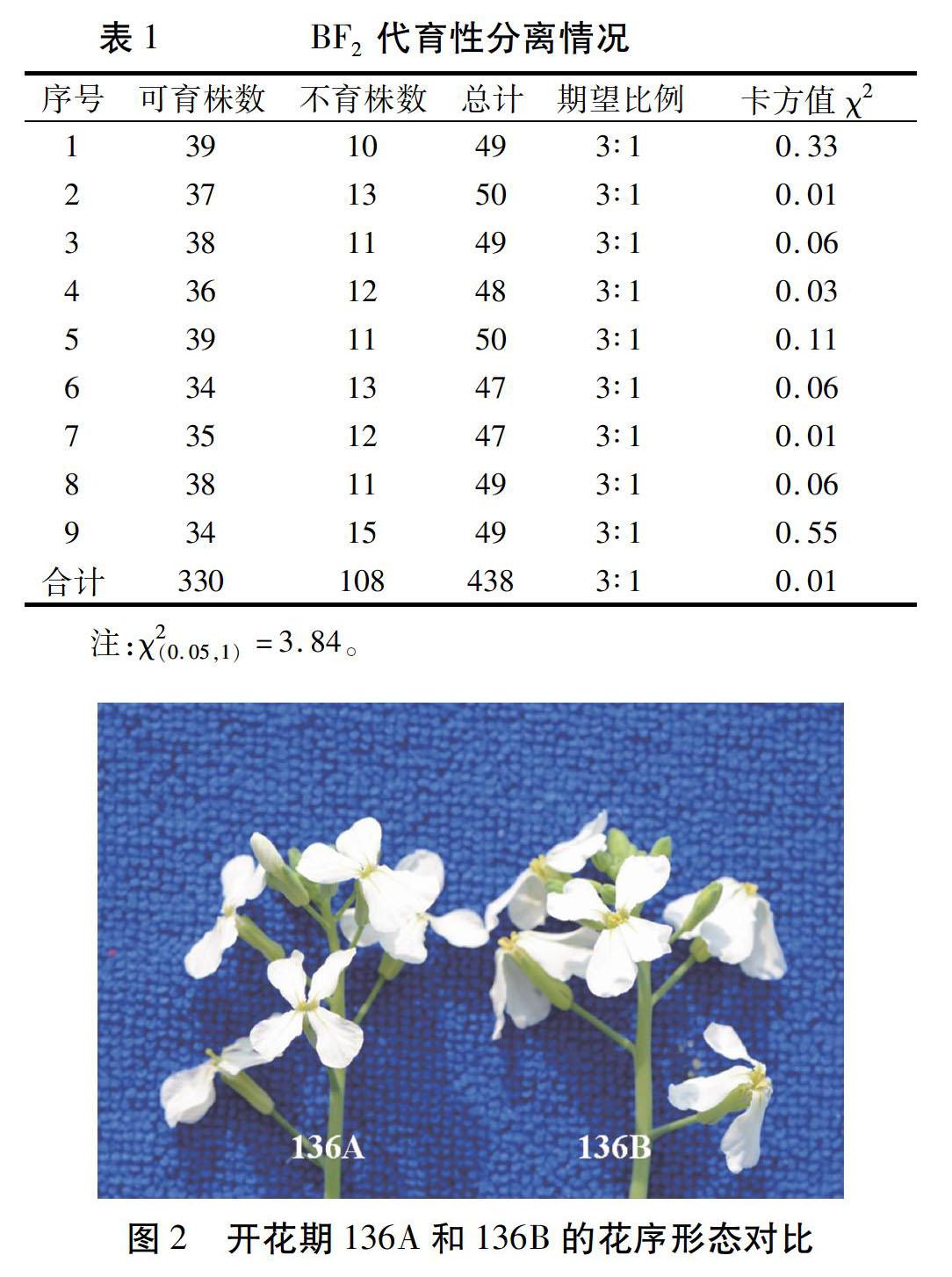
2.3.1花器官發育觀察通過觀察開花期不育系136A和保持系136B的50個花序,選取兩者的典型花序進行對比,如圖2所示。不育系136A與保持系136B的盛花期基本同步,在花器官的大小上,前者比后者明顯要小。
對136A核不育系的花藥與保持系136B進行比較(圖3)發現,剛開放的花花藥就已萎縮的很小,且花藥干癟,摘取花藥擠壓,無花粉,表明該不育系的花藥退化較為徹底。
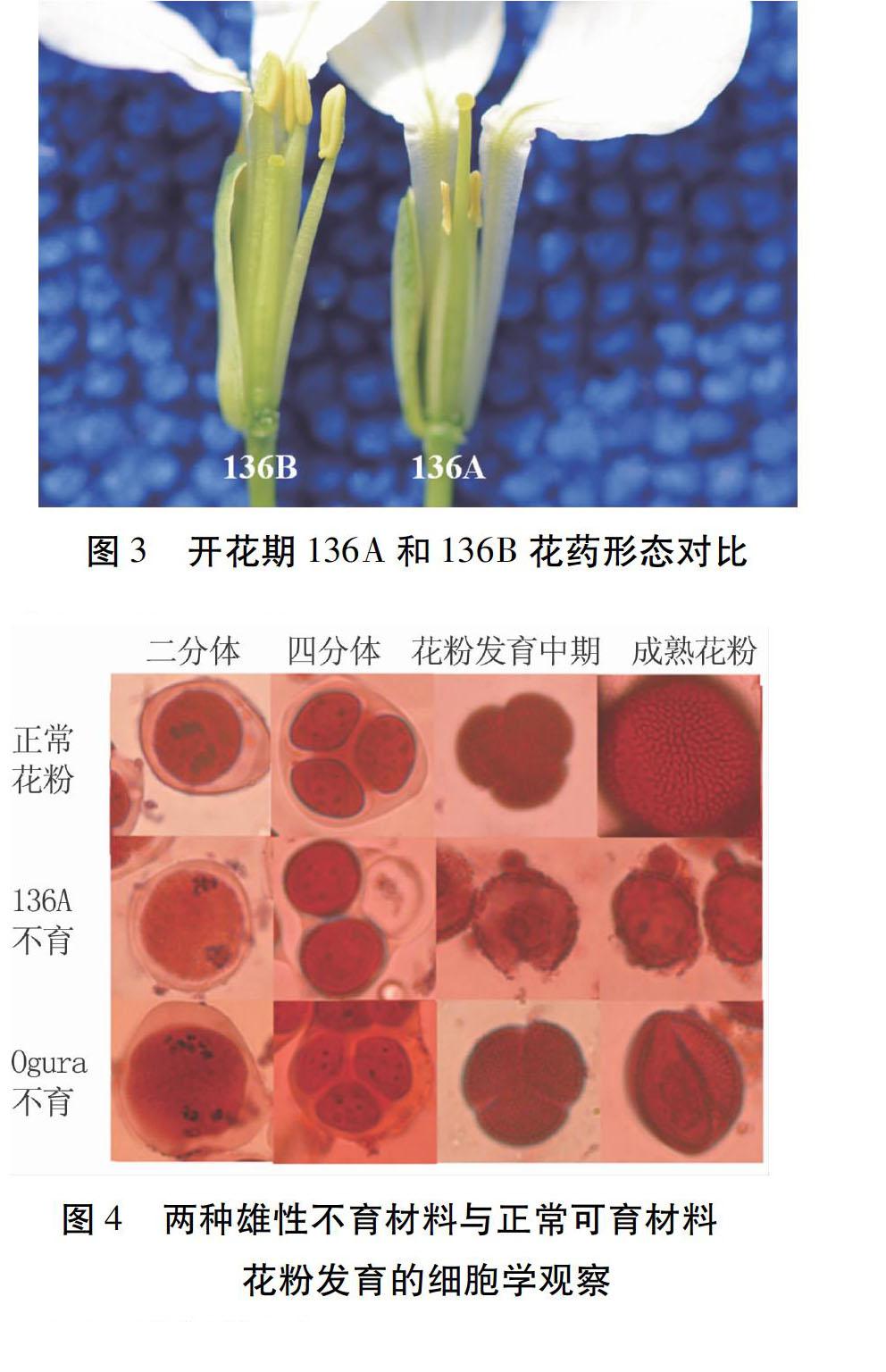
2.3.2花粉粒敗育觀察對136A、136B和RS43的花粉粒發育情況進行觀察(圖4)發現,花粉敗育的轉折點是四分體時期。在四分體以前,無論是核不育、細胞質不育和正常可育材料,花粉發育都是正常的,三者之間沒有明顯的區別。在四分體之后,花粉粒的前體游離成單個個體,正常植株的花粉開始出現花粉壁不均勻增厚,并逐漸增大,最后形成球狀,且外壁有突起;細胞質雄性不育系的花粉粒前體在四分體后,很快就停止發育,并逐漸萎縮,最后形成干癟的花粉粒;核雄性不育系在四分體后,游離出的花粉粒前體也有一個短暫的花粉壁增長過程,但隨著花粉的發育,花粉壁表面越來越粗糙,局部會逐漸變薄,最后破裂,在花蕾開放前,會剩很少一部分花粉粒,花粉壁粗糙,花粉干癟不飽滿。
花粉發育的細胞學觀察
2.4PCR驗證結果
orf138基因特異引物的目標帶型大小為400~500 bp之間,凝膠電泳檢測結果為僅陽性對照RS43中擴增到orf138基因特異條帶;核不育系136A與陰性對照RS45中均未出現特異條帶(圖5)。該結果初步確定核不育材料136A中不含有orf138基因。
1、2、3、4號樣品為136A;5號樣品為RS43;
6號樣品為RS45;M為DL2000。
2.5葉綠體和線粒體測序
以NCBI 公布的蘿卜基因組組裝序列(GCA_000715565.1)及蘿卜的葉綠體、線粒體基因組(KJ716483、AB694744.1)為參考基因組,完成線粒體和葉綠體的測序組裝工作。通過本地Blast,136A(RS44)與RS43及RS45的線粒體基因結構對比如圖6所示,從上至下順序為RS44(即136A)、RS43、RS45。RS44與RS45的線粒體基因組序列完全一致,具有Ogura細胞質的RS43與前兩者差別較大,存在很大的結構變異,出現大量的基因組重排現象。
通過本地Blast,136A(RS44)與RS43及RS45的葉綠體基因結構對比結果如圖7所示,從上至下順序為RS44(即136A)、RS43、RS45。三個材料的葉綠體基因組序列基本一致,其中RS44比RS45僅在其鏡像重復區內多出30個堿基的重復,其它完全一致。對RS44發生變異的區域進行基因序列和開放閱讀框(ORF)比對、尋找,結果顯示,此30個堿基不在已明確的葉綠體基因或ORF內,表明該區域不存在有功能的基因序列。由此推斷,136A不育材料的不育基因不在細胞質中。
3討論與結論
本研究通過連續6代雜交選育與育性調查、細胞學觀察等,明確了136A核雄性不育系的特點。
136A雄性不育材料為核基因控制的隱性單基因雄性不育材料,保持系136B具有50%的保持能力。136A不育系的花器官略小于可育株,雄性育性退化徹底,經過多代觀察,其育性不受環境等因素的影響,表現為雌蕊發育正常,花藥退化徹底,不能形成具有功能的花粉。
蘿卜核雄性不育與細胞質雄性不育花粉發育有著明顯的差異。在四分體之后,花粉粒的前體游離成單個個體,細胞質雄性不育系的花粉粒很快就停止發育,并逐漸萎縮,最后形成干癟的花粉粒;核雄性不育系在四分體后,游離出的花粉粒前體有一個短暫的花粉壁增長過程,但隨著花粉的發育,花粉壁表面越來越粗糙,局部會逐漸變薄,最后破裂,在花蕾開放前,只有很少一部分花粉粒,但花粉壁粗糙,花粉干癟不飽滿。
通過對136A(RS44)、Ogura不育材料(RS43)、正常可育材料(RS45)進行線粒體和葉綠體DNA的測序、組裝,結果表明136A與RS45的線粒體DNA完全一致,具有正常的可育細胞質,不同于Ogura類型的不育細胞質。
新發現的136A雄性不育材料填補了蘿卜雄性不育資源中缺乏核不育材料的空白,為蘿卜雄性不育系的培育與研究提供了重要的不育源。為加速136A雄性不育系的利用,在下一步工作中我們將利用136AB系多代選育過程中建立的近等基因系,采用圖位克隆與測序相結合的方法定位和克隆出核雄性不育基因與恢復基因。針對核雄性不育系不能獲得100%不育株的問題,我們將利用標記基因或對某種藥物敏感基因(如熒光蛋白、水稻苯達松敏感基因[16]等)與恢復基因建立連鎖,將連鎖基因轉化到136A雄性不育系中建立新的保持系,該保持系雖然也只有50%的保持能力,但在不育系與保持系的混合群體中,可通過標記基因或藥物敏感基因表達,在苗期將具有指示性狀的單株拔除或通過噴撒藥物殺死可育株獲得100%的不育株,使得136A隱性核雄性不育系真正應用于生產中。
參考文獻:
[1]馬二磊.蘿卜雄性不育胞質類型鑒定與分析[D].南京:南京農業大學,2009.
[2]Kaul M L H. Male sterility in higher plants. Monographs on theoretical and applied genetics 10[M]. New York:Springer-Verlag,1988.
[3]Li Z F, Xia J F, Tang G Y. Types and genetic mechanisms of plant male sterility[J].Journal of Anhui Agricultural Sciences,2008,28(6):742-746.
[4]Johns C W, Delannay X, Palmer R G, et al. Structural sterility controlled by nuclear mutations in angiosperms[J].The Nucleus,1981,24:97-105.
[5]Okamuro J K, Denb G W, Jofuku K D, et al. Regulation of Arabidopsis flower development[J]. Plant Cell,1993,5:1183-1193.
[6]陳鳳祥,胡寶成,李強生,等. 甘藍型油菜細胞核雄性不育材料9012A的發現與初步研究[J].北京農業大學學報,1993,19(增刊):57-61.
[7]張書芳,宋兆華,趙雪云.大白菜細胞核基因互作雄性不育系選育及應用模式[J].園藝學報, 1990, 17(2):117-125.
[8]陳鳳祥,胡寶成,李強生,等.甘藍型油菜隱性上位互作核不育雙低雜交種皖油14的選育[J].中國油料作物學報,2003,25(1):63-65.
[9]陳鳳祥,胡寶成,李強生,等.甘藍型油菜隱性上位互作核不育雙低雜交種“皖油18”的選育[J]. 安徽農業科學,2002,30(4):535-537.
[10]孫超才,趙華,王偉榮,等.甘藍型雙低隱性核不育雜交種滬油雜1號的選育[J].中國油料作物學報,2004,26(1):63-65.
[11]陳大倫,張瑞茂.甘藍型油菜隱性細胞核雄性不育系118A的發現與選育[J].貴州農業科學,2006,34(6):5-7.
[12]李慧,胡勝武,李瑋,等.甘藍型油菜隱性上位互作核不育系的選育及其細胞學研究[J].西北農林科技大學學報,2010,38(1):111-124.
[13]Hondred D, Young J K, Brink K, et al. Plant genomic DNA flanking SPT event and methods for identifying SPT event:US20090210970A1[P]. 2009.
[14]Paterson A H, Brubaker C, Wendel J F, et al. A rapid method for extraction of cotton (Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis[J]. Plant Mol. Biol. Rep., 1993,11:122-127.
[15]段乃彬,王俊峰,白靜,等.蘿卜葉綠體及線粒體基因組測序與組裝[J].分子植物育種, 2015,13(11):2429-2436.
[16]Wang S M, Zhu Q S, Wang W L, et al. The application of bentazon susceptible gene on seed production of hybrid rice[J]. Agricultural Science & Technology, 2008,9(1):99-103,145.
