DHAP脅迫對天山云杉幼苗生長過程中抗氧化酶活性的影響
2016-04-20 11:20:11劉潤楊麗阮曉潘存德王強
天津農業(yè)科學 2016年4期
劉潤 楊麗 阮曉 潘存德 王強


摘 要:天山云杉(Picea schrenkiana)是新疆森林生態(tài)系統(tǒng)中的特有種,自然更新過程出現(xiàn)了阻礙,3,4-二羥基苯乙酮(3,4-dihydroxy-acetophenone,DHAP )已被確證是引起這一生態(tài)過程的主要自毒物質。為了揭示天山云杉天然更新障礙的內在生理機制,本文選擇5個濃度梯度(0,1.0,2.0,3.0,5.0 mmol·L-1)DHAP處理天山云杉幼苗,以胚根長、胚芽長和鮮質量作為幼苗生長參數(shù),測定天山云杉幼苗根尖細胞活力和6種抗氧化酶活力(SOD、POD、CAT、APX、GPX、GR)。結果顯示,2.0 mmol·L-1 DHAP能顯著促進天山云杉幼苗的胚根、胚芽和鮮質量生長,5.0 mmol·L-1 DHAP 顯著抑制天山云杉幼苗的生長;天山云杉在DHAP濃度(c≤2.0 mmol·L-1)處理下,抗氧化酶SOD、CAT、APX、GPX、GR的活性隨著DHAP濃度的升高而升高,POD的活性隨著濃度的升高而下降,6種抗氧化酶協(xié)同作用以維持抗氧化酶防御系統(tǒng)的穩(wěn)定; DHAP濃度(2.0 mmol·L-1 < c ≤5.0 mmol·L-1)處理后的前3 d,抗氧化酶SOD、CAT、APX、GPX、GR的活性隨著DHAP濃度的升高而降低,POD的活性隨著濃度的升高而升高,處理3 d后6種抗氧化酶活性與對照相比差異性顯著但無規(guī)律,在此濃度范圍內DHAP脅迫導致天山云杉生理機制紊亂,打破了正常的生長發(fā)育機制致使細胞加速死亡。DHAP通過影響天山云杉幼苗根尖細胞活力和抗氧酶活性,進而干擾天山云杉幼苗的正常生長,成為引發(fā)天山云杉天然更新障礙的一種因素。
關鍵詞:3,4-二羥基苯乙酮;幼苗生長;抗氧化酶;根尖細胞活性
中圖分類號:S718 文獻標識碼:A DOI 編碼:10.3969/j.issn.1006-6500.2016.04.001
Abstract: Schrenk spruce (Picea schrenkiana) is a major tree species in the forest ecosystems of Xinjiang , China. But the natural regeneration of P. Schrenkiana has been disturbed. 3,4-dihydroxy-acetophenone (DHAP) , a major autotoxic substance which separated from P. Schrenkiana needles, has effect on the growth. This experiment was to study the physiological mechanism of antioxidant enzyme defense system and to elucidate the underling regulation on natural regeneration barrier of P. Schrenkiana. The growth radical, plumule and fresh weight of the seedings that treated with DHAP were measured, the activity of antioxidant enzyme measured too. The results showed that 2.0 mmol·L-1 DHAP treatment had significant effect on the growth radicle, plumule, fresh weight and root cell viability, which significantly promote the seeding growth. But 5.0 mmol·L-1 DHAP treatment had significant inhibition on growth of P. Schrenkiana. 1.0 mmol·L-1 and 3.0 mmol·L-1 DHAP treatment could promote or inhibit the seedings growth, but there had no significant. The other result showed that DHAP could change the activity of antioxidant enzyme, such as super oxidedismutase (SOD), guaiacol peroxidase (POD), glutathione reductase (GR) and so on. It seems that at higher concentration, DHAP showed significantly effects on the activity of antioxidant enzyme except for POD, however the effects were not significantly at lower concentration.
Key words: 3,4-dihydroxy acetophenone (DHAP); seeding growth; antioxidant enzymes; root cell activity
植物化感作用是指植物通過向環(huán)境中釋放一種或多種具有生物活性的化學物質,對周圍植物(包括微生物)產生直接或間接的有害或有利的作用[1]。化感作用通常發(fā)生在種間[2-3],但同種植物間也存在化感作用,這種現(xiàn)象被稱為自毒[4]。自毒作用廣泛存在于植物森林生態(tài)系統(tǒng)中[5],已有研究發(fā)現(xiàn)赤松(P. densiflora)、白松(P. halepensis)、馬尾松(P. massoniana)、紅松(P. koraiensis)等分泌或分解的自身物質有干擾自身種子萌發(fā)、幼苗生長以及幼苗建成的現(xiàn)象[6],同時自毒作用也是引起森林天然更新障礙的重要原因[7]。
天山云杉(Picea schrenkiana)是亞洲中部針葉林中特有的云杉屬植物種,主要分布在天山及昆侖山西北坡,是構成天山以及新疆森林生態(tài)系統(tǒng)的物質主體,對新疆山地水源涵養(yǎng)、水土保持,以及林區(qū)生態(tài)系統(tǒng)的形成與維護有著至關重要的作用[8]。全球氣候變化和經濟建設對森林物質產品需求的增長,以及人類活動對森林生態(tài)系統(tǒng)干擾強度的增加,導致天山云杉生產力下降、地力衰減及天然更新問題日益突出[9]。在無火干擾的有限時間森林自然恢復中,自毒作用可能是導致天山云杉更新障礙的主要因素[10]。Ruan等曾提出“天山云杉天然更新障礙的自毒作用”假說,并且證實了從天山云杉針葉和凋落物中分離純化出的3,4-二羥基苯乙酮(DHAP)是引起天山云杉自毒作用的主要化學物質之一。那么,DHAP是如何從生理途徑上影響天山云杉自然更新過程的呢?對DHAP是如何從生理學的途徑影響天山云杉更新過程的探討,將有助于進一步揭示自毒作用發(fā)生的內在規(guī)律。
越來越多的研究已經把化感物質看作一種生物脅迫,植物中存在的抗氧化酶系統(tǒng)對環(huán)境脅迫下植物體內活性氧清除起到極其重要的作用[11]。目前有關化感物質脅迫條件下對抗氧化酶活性變化的報道較少。從云杉凋落物水浸提取液有機層中分離出來的以云杉特定代謝物命名的對羥基苯乙酮(p-hydroxyacetophenone)能夠影響其幼苗根系的生長[12],且表明自毒物質能夠抑制幼苗的生長或者延遲其發(fā)芽和限制后代的繁殖[13]。 此外,從黃瓜根系中分離提取的化感物質(苯甲酸、肉桂酸)能顯著激活POD、SOD、CAT等的活性[14],從而影響植株生長。有研究者對苜蓿幼苗不同器官的SOD、APX、CAT等抗氧化酶在干旱、鹽漬等逆境脅迫下進行研究,結果表明,在逆境脅迫中抗氧化酶系統(tǒng)在苜蓿成苗過程中起著重要的保護作用。上述研究表明,化感物質能夠響應自毒物質對植物幼苗的脅迫作用。但化感物質對植物幼苗生長過程中抗氧化酶調控作用的研究尚未見報道。本研究以胚芽長、胚根長和鮮質量的變化為參數(shù),用4個濃度處理天山云杉幼苗,分時間段測定6種酶的活性變化,探討天山云杉發(fā)生自毒作用的生理機制。
1材料和方法
1.1 材 料
天山云杉種子于2014年10月采自于新疆農業(yè)大學實習林場2林班(樺樹溝N43°23′19″~N43°23′25″、E86°49′14″~E86°49′19″,海拔1 730~1 745 m)。3,4-二羥基苯乙酮(DHAP)購于上海百靈威科技有限公司,其他試劑均購于寧波杭景生物科技有限公司。
1.2 方 法
1.2.1 DHAP處理對種子萌發(fā)和幼苗生長的影響 選擇籽粒飽滿的天山云杉種子,用去離子水培養(yǎng)至胚根突破種皮,取100粒胚根突破的種子均勻擺放在鋪有濾紙的25 cm×25 cm的培養(yǎng)盒內,加入10 mL不同濃度的DHAP處理液(對照組加入去離子水),每個處理設置3個重復。將培養(yǎng)盒放入人工氣候培養(yǎng)箱,培養(yǎng)條件:光周期20 ℃,16 h;暗周期15 ℃,8 h;光強40 nmol·m-2·s-1。每3 d統(tǒng)計天山云杉幼苗的胚根長、胚芽長和鮮質量及測定抗氧化酶的活性,第12天結束試驗。
1.2.2 根細胞活力測定 伊文斯蘭染色法是測定根細胞活性的一種常用的方法[14],隨機選擇不同濃度DHAP處理的天山云杉幼苗,截取根尖處1.0 cm,用0.25% (w/v) 的伊文斯蘭水溶液在30 ℃的暗環(huán)境下浸泡1 h;蒸餾水沖洗干凈后,用N,N-二甲基酰胺(N,N-dimethylformamide)在30 ℃的暗環(huán)境下浸泡24 h,然后利用紫外線分光光度計測其在600 nm處的吸光度(Beckman DUO 640; Beckman Coulter Inc., Fullerton, CA, USA)。
1.2.3 酶活性的測定 參照Bai等[15]和Soares等[16]的提取方法并加以調整:每次從每個處理中隨機稱取0.1 g種子,加入1.0 mL含有EDTA的磷酸緩沖液研磨,用磷酸緩沖液沖洗研缽定容至10 mL,靜置、取上清液于10 000 r·min-1離心20 min,4 ℃保存用于抗氧化酶活性的測定,測定方法參照張玉竹等[17]的抗氧化酶活性測定方法。
1.3 數(shù)據(jù)分析與處理
相對抑制率=(試驗組-對照組)×100%。
采用Excel、SPSS、Sigmaplot軟件對數(shù)據(jù)進行處理分析。利用單因素方差分析和最小顯著差數(shù)法(Least signnificant difference, LSD)多重比較分析幼苗胚根長、胚芽長和鮮質量的差異。
2 結果與分析
2.1 DHAP處理對天山云杉幼苗胚根長、胚芽長和鮮質量的影響
試驗結果如圖1所示,DHAP處理對天山云杉胚根、胚芽和鮮質量的影響表現(xiàn)為促進和抑制兩種:1.0 mmol·L-1和2.0 mmol·L-1的DHAP處理對天山云杉生長發(fā)育具有促進作用,且后者促進作用最為顯著;3.0 mmol·L-1和5.0 mmol·L-1 DHAP處理對天山云杉生長發(fā)育有抑制作用,且后者抑制作用最顯著。試驗結果表明:天山云杉在低濃度的DHAP作用下能夠更好地生長,但高濃度的DHAP脅迫下其正常生長受到了嚴重的抑制作用。
2.2 DHAP處理對天山云杉幼苗生長過程中根細胞活力的影響
伊文斯蘭是一種細胞活性染料,常用于檢測細胞膜的完整性和壞死的細胞[14]。如表1所示,與對照組相比,在3.0 mmol·L-1和5.0 mmol·L-1 DHAP 處理組伊文斯蘭的吸收率遠遠高于其他處理組,且后者更為顯著。1.0 mmol·L-1和2.0 mmol·L-1 DHAP處理組伊文斯蘭的吸收率低于對照組,且后者更為顯著。試驗結果表明:5.0 mmol·L-1 DHAP處理組比3.0 mmol·L-1 DHAP處理組更容易誘導天山云杉根細胞死亡。1.0,2.0 mmol·L-1 DHAP處理組能夠延緩天山云杉根細胞的死亡,尤其是2.0 mmol·L-1 DHAP處理組最為顯著。
2.3 DHAP處理對種子萌發(fā)過程中抗氧化酶活性的影響
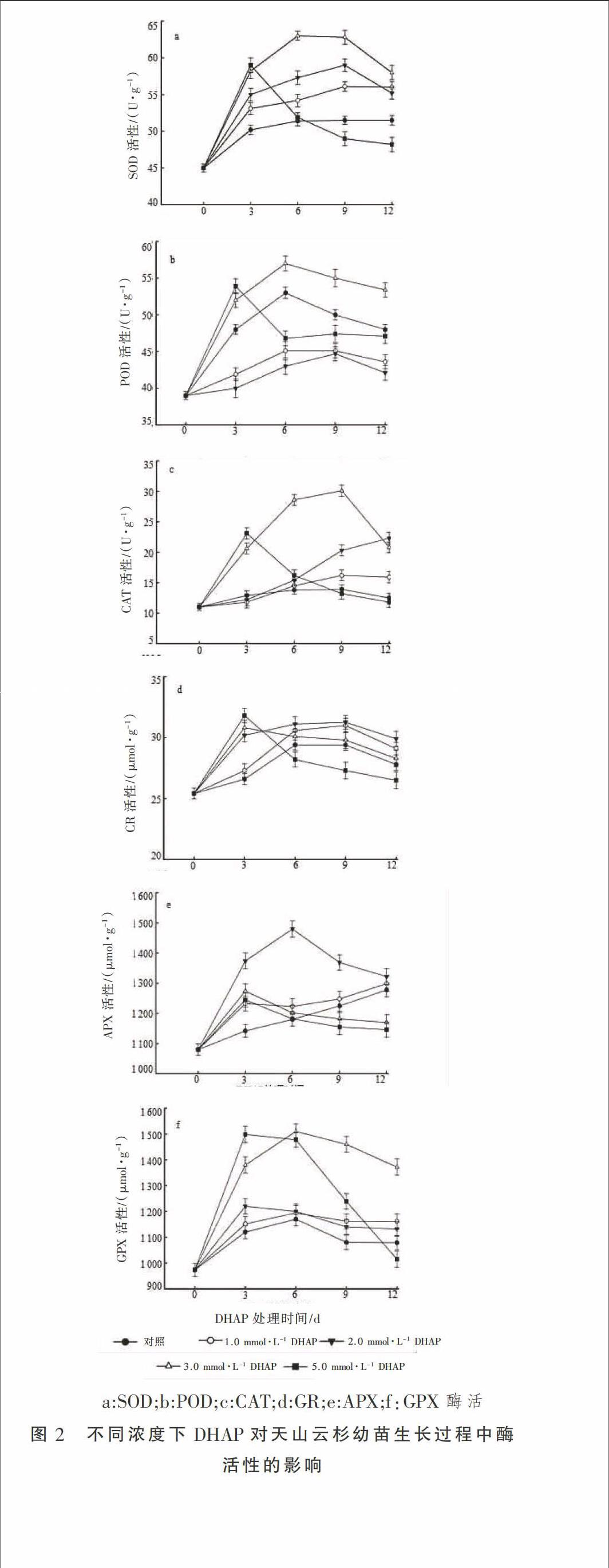
2.3.1 DHAP對天山云杉幼苗生長過程中SOD和CAT活性的影響 SOD在植物抗氧化防御系統(tǒng)中起主導地位,它的主要作用是將細胞內過量的 O2-轉化為H2O2,H2O2 再通過CAT等協(xié)同作用來降低ROS對其細胞造成的氧化損傷,維持植物細胞正常的生理活動。由圖2-a和2-c可以看出,天山云杉幼苗生長過程中對照組SOD和CAT的活性基本不變。與對照相比,在處理后的第3天 SOD和CAT活性隨著DHAP濃度的增加而升高。5.0 mmol·L-1 DHAP 處理組SOD、CAT活性在3 d后開始大幅度下降,至第9 天比對照分別降低了4.85%和3.45%,1.0,2.0,3.0 mmol·L-1 DHAP處理組SOD、CAT活性變化如圖2顯示。由試驗結果分析,天山云杉幼苗受到DHAP的脅迫時,植物細胞就會調節(jié)自身的代謝機制,升高SOD、CAT的活性消除過量的 O2-、H2O2以抵抗脅迫適應環(huán)境。5.0 mmol·L-1 DHAP 處理組的原因可能是高濃度的DHAP促使其代謝產生大量的ROS而得不到及時清除,使細胞發(fā)生毒害作用,導致抗氧化系統(tǒng)破壞,致使SOD、CAT活性迅速下降。因此,可以得出一定濃度的DHAP(2.0 mmol·L-1)能夠促進SOD、CAT活性的升高,從而能更有效地清除過量的ROS,維持植物細胞正常的生理活動,但是高濃度的DHAP(≥3.0 mmol·L-1)則會引起植物產生大量的ROS,發(fā)生氧化損傷。
2.3.2 DHAP對天山云杉幼苗生長過程中POD活性的影響 POD在活性氧的代謝過程中也起著重要作用。由圖2-b 可以看出,第3天1.0 mmol·L-1和2.0 mmol·L-1 DHAP處理組POD活性比對照分別降低了12.71%和16.67%,3.0,5.0 mmol·L-1 DHAP處理組POD活性比對照分別增加了8.33% 和12.29%。而5.0 mmol·L-1 DHAP處理組于第6天后POD的活性基本保持不變且低于對照組,原因同5.0 mmol·L-1 DHAP處理組SOD活性變化的原因一致。POD在細胞壁中可催化NADH或NADPH產生O2-,POD與CAT都具有代謝H2O2的功能,但在愈創(chuàng)木酚存在下其主要參與植物細胞的木質化和木栓化過程。POD的活性越高,其抗逆性就會越弱。由試驗結果得出:2.0 mmol·L-1 DHAP處理組POD活性最低,最有利于植物細胞的生長發(fā)育,而5.0 mmol·L-1 DHAP處理組POD活性最高,說明該濃度能夠加快細胞老化,3.0 mmol·L-1次之。
2.3.3 DHAP對天山云杉幼苗生長過程中APX、GPX和GR活性的影響 在植物體中,抗壞血酸-谷胱甘肽循環(huán)(AsA-GSH)是一個主要的H2O2清除系統(tǒng)。它由APX、GPX和GR相互調節(jié)代謝機制完成其生理功能,其中APX是最主要的清除酶且與H2O2有更好的親和力。植物細胞中GPX的主要作用不是清除H2O2,而是參與新陳代謝的反應,如木栓化、木質化和IAA的分解。由圖2-c可以看出,隨著處理時間的增加,1.0 mmol·L-1 DHAP處理組變化趨勢與對照相似,2.0 mmol·L-1 DHAP處理組APX的活性均高于其他試驗組,3.0 mmol·L-1 和5.0 mmol·L-1 DHAP處理組都是在處理第3天達到最大值后開始下降,最終低于對照組。對此試驗現(xiàn)象的解釋是,2.0 mmol·L-1 DHAP 處理能夠更好地促進APX代謝清除H2O2。3.0 mmol·L-1 DHAP和5.0 mmol·L-1 DHAP處理組為適應脅迫響應,APX活性升高,當植物細胞產生大量的ROS時,其細胞受到氧化損傷致使APX的活性開始大幅度的下降。
GPX和GR主要是完成AsA-GSH循環(huán)中GSSG和GSH的相互轉化,GPX和GSH是由GR催化NADPH再生的,進而促進APX和CAT對H2O2的清除。由圖2-e和2-f可以看出,在DHAP處理第3天,隨著濃度的增大,GR和GPX活性變化趨勢一致。在圖2-e中,1.0 mmol·L-1 和2.0 mmol·L-1 DHAP處理組隨著處理時間的延長,其變化趨勢與對照基本保持一致且始終是2.0 mmol·L-1 >1.0 mmol·L-1 >對照。而3.0 mmol·L-1 和5.0 mmol·L-1 DHAP處理組隨著處理時間的延長出現(xiàn)下降的趨勢,可能原因是前期的抗氧化循環(huán)中植物細胞發(fā)生氧化損傷,致使GR活性下降。由此分析得出,在2.0 mmol·L-1 DHAP處理下,能夠有效地調節(jié)GR活性,促使植物細胞正常的生長發(fā)育。而5.0 mmol·L-1 DHAP處理組能破壞其正常的抗氧化防御系統(tǒng),致使植物細胞死亡。在圖2-f中可以看出,1.0 mmol·L-1 和2.0 mmol·L-1 DHAP處理組GPX的變化趨勢和GR一致,由于GPX是通過GR等一系列的生理調控機制生成,所以3.0 mmol·L-1和5.0 mmol·L-1 DHAP處理組的變化趨勢與GR相比出現(xiàn)滯后現(xiàn)象,但是最終的結果是一致的。因此可以得出,2.0 mmol·L-1的DHAP處理組能夠促進GPX更好地完成其生理活動,促使植物正常生長發(fā)育,而5.0 mmol·L-1 DHAP 則會引起植物細胞發(fā)生氧化損傷。
3 討 論
以往研究植物的化感作用時,幼苗的胚根長、胚芽長以及鮮質量都會被看做實生苗的生長參數(shù)[18]。不同濃度DHAP作為一種生物活性化合物可以促進或抑制植物的生長,然而,它對濃度的確定取決于植物對化感物質劑量的敏感性[19]。本試驗的研究結果與對人參皂苷(ginsenosides)的研究結果一致。低濃度的人生皂苷(12.5 mg·L-1;25 mg·L-1)能夠顯著促進西洋參的生長,而高濃度(25 mg·L-1;100 mg·L-1)的人生皂苷能夠抑制西洋參的生長[20]。由此可以說明,化感物質能夠影響植物,即適宜濃度的化感物質可以維持植物正常的生理代謝延緩植物的衰老;但是當濃度超過一定的范圍時,就會破壞正常的生理代謝,致使細胞死亡。
在正常條件下,抗氧化酶可以調控細胞內H2O2的水平,但在逆境脅迫中,當ROS積累過量,它的產生與消除的動態(tài)平衡就會被破壞,從而引起細胞損傷[21]。為了適應脅迫所造成的氧化損傷,植物會做出一系列的反應來保護其正常的生理代謝。抗氧化酶防御系統(tǒng)是抗氧化保護中一個極其重要的組成部分,且其防御能力大小取決于多種酶的活性和彼此間協(xié)調的綜合結果[22]。一般研究認為,在逆境脅迫下植物生長發(fā)育好的植物有更高的抗氧化酶活性[23]。本試驗中2.0 mmol·L-1 DHAP處理組SOD活性變化與鹽脅迫下對豌豆、甘蔗、玉米[24]的研究結果一致,表明此條件下SOD對O2- 有更強的清除能力,減少了O2- 對細胞的損害作用。5.0 mmol·L-1時,SOD的活性與人參皂苷對西洋參的研究結果一致[20],可能的原因是在高濃度的DHAP作用下產生大量的H2O2,當其不能被迅速清除時細胞就會受到氧化損傷死亡。
SOD在歧化過程中產生H2O2可以氧化損傷細胞膜,因此需要快速清除H2O2。植物中APX和CAT活動途徑可以將H2O2轉化為H2O分子調節(jié)H2O2平衡。已有研究表明,GR和GPX通過ASA-GSH循環(huán)途徑協(xié)助完成APX、POD和CAT的生理作用機制。在鹽脅迫下對番茄[25]、甘蔗和玉米等的研究也表明,在逆脅迫下作物APX、POD和CAT相互協(xié)調完成其生理機制。由試驗結果可看出,CAT、APX的活性在2.0 mmol·L-1 DHAP處理下均顯著高于其他試驗組,但在5.0 mmol·L-1 處理下結果與之相反。這表明,低濃度下CAT和APX的活性強于高濃度下的活性,因此增強它們對H2O2的清除,使植物細胞免受更嚴重的氧化損傷。該試驗結果與伊文斯蘭染色檢測細胞活性的結果一致。
POD 是植物在逆境條件下酶促防御系統(tǒng)的關鍵酶之一。它不僅參與了清除H2O2的過程而且還參與了植物木質化和木栓化[26]。已有研究表明,在逆境中車前草[27]、向日葵[28]等抗逆性較弱植物的POD的活性較高,本試驗結果與上述研究結果相同。引起此試驗結果的原因是:POD的活性增強致使木質素積累加速了細胞的衰老;而適宜濃度的DHAP能夠合理調控其生理機制,阻礙了天山云杉在生長發(fā)育過程中受到的氧化損傷。
植物的抗氧化酶是由植物自身代謝產生的一類調節(jié)植物生長的物質,在極低的濃度下也能產生生理效應,與植物的生長發(fā)育以及抗逆性密切相關[29]。不同濃度的DAHP對天山云杉幼苗生長過程中抗氧化酶活性的影響不同;抗氧化酶活性的變化可能會干擾活性氧生產和清除的平衡:因此,濃度的化感物質可能是引起天山云杉自然更新的原因之一。
4 結 論
通過對幼苗生長發(fā)育過程中根長、胚芽長和鮮質量的分析得出,低濃度的DHAP(c ≤ 2.0 mmol·L-1)能夠促進幼苗的生長而高濃度的DHAP(2.0 mmol·L-1 < c ≤ 5.0 mmol·L-1)抑制天山云杉幼苗的生長。通過對天山云杉幼苗根細胞抗氧化酶活性的綜合分析得出,DHAP作為化感物質能夠誘導天山云杉生長過程中6種抗氧化酶的含量發(fā)生變化,DHAP處理濃度在( c ≤ 2.0 mmol·L-1)時,抗氧化酶SOD、CAT、APX、GPX、GR的活性隨著DHAP濃度的升高而升高,POD的活性隨著濃度的升高而下降;DHAP處理濃度在(2.0 mmol·L-1 < c ≤ 5.0 mmol·L-1)時,抗氧化酶SOD、CAT、APX、GPX、GR的活性在前3 d隨著DHAP濃度的升高而降低,POD的活性隨著濃度的升高而降低,3 d后6種抗氧化酶活性隨著處理時間的延長變化規(guī)律紊亂。試驗結果表明,天山云杉幼苗生長發(fā)育狀況與6種抗氧化酶活性強弱密切相關,同時這6種抗氧化酶活性強弱與其受到脅迫因子的濃度有直接關系。
參考文獻:
[1] CALLAWAY, R M, VIVANCO J M. Can plant biochemistry contribute to understanding of invasion ecology?[J].Trends Plant Science, 2006, 11(12): 574-580.
[2] CALLAWAY R M, ASCHEHOUG E T. Invasive plants versus their new and old neighbors: a mechanism for exotic invasion[J]. Science, 2000, 290(5491): 521-523.
[3] WEIDENHAMER J D, HARTNETT D C, ROMEO J T. Density-dependent phytotoxicity: distinguishing resource competition and allelopathic interference in plants[J]. Journal of Applied Ecology, 1989, 26(2): 613-624.
[4] YU J Q, SHOU S Y, QIAN Y R, et al. Autotoxic potential of cucurbit crops[J]. Plant Soil, 2000, 223(1/2): 149-153.
[5] ZENG R S, MALLIK A U, LUO S M. Allelopathy in Sustainable Agriculture and Forestry[M]. New York:Springer, 2008: 363-386.
[6] FERNANDEZ C,VOIRIOT S,MEVY J P, et al. Regeneration failure of Pinus halepensis Mill. The role of autotoxicity and some abiotic environmental parameters[J]. Forest Ecology and Management, 2008, 255(7) : 2928-2936.
[7] HALLIWELL B, FOYER C H. Properties and Physiological function of a glutathione reductase purified from spinach leaves by affinity chromatography[J]. Planta ,1978, 139(1):9-17.
[8] WANG Q, RUAN X, PAN C D, et al. Need for sustainability policy-A case study of the National Forest Conservation Program (NFCP)in the western region of Tianshan Mountain,China[J].The Forestry Chronicle, 2006, 82(1) : 31-39.
[9] 張新時,張瑛山.烏蘇林區(qū)天山云杉天然更新的初步研究[J].新疆農業(yè)科學,1963(1): 8-12.
[10] LI Z H, WANG Q, RUAN X, et al. Phenolics and plant Allelopathy[J]. Molecules, 2010, 15( 12): 8933-8952.
[11] BROZ A K, BROECKLING C D, DELAPEA C. Plant neighbor identity influences plant biochemistry and physiology related to defense[J]. BMC Plant Biology, 2010, 10(1):115.
[12] FALIK O, REIDES P, GERSANI M, et al. Self/non-self discrimination in roots[J]. Journal of Ecology, 2003, 91(4):525-531.
[13] WEIR T L, PARK S W, VIVANCO J M, Biochemical and physiological mechanisms mediated by allelochemicals[J]. Current Opinion Plant Biology, 2004, 7(4):472-479.
[14] TAM S L, IMONOVIOV M, HUTTOV J, et al. Aluminium stimulated hydrogen peroxide production of germinating barley seeds[J]. Environment and Experimental Botany, 2004, 51(3): 281-288.
[15] BAI R , MA F , LIANG D, et al. Phthalic aicd induces oxidative stress and alters the activity of some antioxidant enzymes in roots of Malus prunifolia [J]. Journal of Chemical Ecology, 2009,35(4):488-494.
[16] SOARES A R, FERRARESE M L,DE CASSIA SIQUEIRA-SOARES R, et al. The allelochemical L-DOPA increases melanin production and reduces reactive oxygen species in soybean roots[J]. Journal of Chemical Ecology, 2011,37(8):891-898.
[17] 張玉竹,劉潤,潘存德,等. DHAP脅迫對天山云杉種子萌發(fā)過程中抗氧化酶活性影響的研究[J].新疆農業(yè)大學學報,2015,38(1):1-11.
[18] LEATHER G R, EINHELLING F A. Bioassays in the study of allelopathy II[M]//PUTNAM A R,TANG C S. The Science of allelopathy. New York: John Wiley,Sons,1986:133-145.
[19] LI X X, YU M F, RUAN X, et al. Phytotoxicity of 4, 8-dihydroxy-1-tetralone Isolated from Carya cathayensis Sarg. to various plant species[J]. Molecules, 2014,19(10): 15452-15467.
[20] ZHANG A H, LEI F J, FANG S W. Effects of ginsenosides on the growth and activity of antioxidant enzymes in American ginseng seedlings[J]. Journal of Medicinal Plants Research, 2011,5(14) :3217-3223.
[21] ZHOU Z S,WANG S J, YANG Z M. Biological detection and analysis of mercury toxicity to alfalfa plants[J]. Chemosphere, 2008,70(8):1500-1509.
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
