臨床樣品中不同蝕斑表型單克隆輪狀病毒VP8基因序列分析
2016-03-29 01:25:53錢雯馬波黃永坤陳敏王麗麗張益麗楊晨徐道俊劉馨
中國醫(yī)藥生物技術 2016年1期
錢雯,馬波,黃永坤,陳敏,王麗麗,張益麗,楊晨,徐道俊,劉馨
?
臨床樣品中不同蝕斑表型單克隆輪狀病毒VP8基因序列分析
錢雯,馬波,黃永坤,陳敏,王麗麗,張益麗,楊晨,徐道俊,劉馨
作者單位:650106 昆明,云南沃森生物技術股份有限公司(錢雯、馬波、陳敏、王麗麗、張益麗、楊晨、徐道俊、劉馨);650032 昆明醫(yī)學院第一附屬醫(yī)院(黃永坤)
輪狀病毒屬于呼腸孤病毒屬,為沒有包膜的雙鏈 RNA病毒,人畜禽共患,其感染每年引起約 50 萬嬰幼兒死亡[1]。
宿主細胞表面的寡糖分子是一個重要的因子,它們與輪狀病毒和細胞的相互識別及吸附有關,同時還影響了病毒的宿主特異性[2]。研究表明,輪狀病毒外殼蛋白 VP4 的糖類結合區(qū)域位于 VP4 的亞基 VP8,其與細胞表面寡糖分子的相互作用使病毒識別并吸附到細胞表面。輪狀病毒對細胞的感染是通過 VP8 與細胞表面多糖,尤其是與內部唾液酸(N-acetyl- and N-glycolylneuraminic acids,Sia)的結合完成的[3]。人株、猴株以及豬輪狀病毒 VP8 基因序列在識別細胞表面唾液酸或其衍生物甲基-α 二氮乙酰神經氨酸的位點有顯著的差異[1],影響病毒在細胞培養(yǎng)上的敏感性和適應性。
研究發(fā)現,分離自豬糞便的輪狀病毒在 157 位呈現出Pro 或者 Ser 的多態(tài)性;牛輪狀病毒株 NCDV 隨細胞培養(yǎng)的適應,VP8 基因 157 位由 Pro 突變?yōu)?Ser,從而產生了NCDV Cody 和 NCDV Lincoln 兩個衍生亞株,這是在VP8 氨基酸 64 ~ 224 區(qū)域內發(fā)生的唯一的氨基酸突變[4]。分離自人糞便樣品輪狀病毒的 VP8 基因是否存在上述位點的多態(tài)性?
輪狀病毒感染細胞后可形成蝕斑,在凝膠的固定下,病毒顆粒不能自由地移動,在一個病毒顆粒進入細胞并產生子代病毒的過程中,細胞死亡并形成蝕斑,一個蝕斑代表了一個單克隆的病毒分子。蝕斑的表型,即蝕斑的大小與病毒感染后產生細胞致病作用(cytopathogenic effect,CPE)的時間和程度有關,蝕斑較大的病毒克隆產生 CPE 的時間較快,也較為劇烈。VP8 基因序列的差異與空斑的大小即蝕斑表型、病毒分子的感染性之間存在怎樣的關系?
為了回答上述問題,本文研究了分離自一份臨床糞便樣品的輪狀病毒單克隆的 VP8 基因序列,并分析了序列差異與病毒蝕斑表型之間可能存在的關系。
1 材料與方法
1.1材料
1.1.1臨床樣品及病毒臨床糞便樣品來自昆明醫(yī)學院第一附屬醫(yī)院兒科秋冬季腹瀉患兒,經病毒核酸電泳或電鏡觀察確證為輪狀病毒感染。
1.1.2細胞及質粒羅猴腎細胞系 MA104 購自中國典型培養(yǎng)物保藏中心,CCTCC 編號為 GDC041;pGEM-T easy載體為美國 Promega 公司產品。
1.1.3試劑與儀器小量液體樣品 RNA 抽提試劑盒、聚丙烯酰胺、硝酸銀、中性紅購自上海華舜生物技術有限公司;RT-PCR 試劑盒、ExTaq 酶購自大連寶生物工程有限公司;小型垂直電泳槽購自美國 Bio-Rad 公司;MEM 培養(yǎng)基、胎牛血清購自美國 Gibco 公司;胰酶購自美國 BBI 公司;二乙氨乙基葡聚糖(DEAE-dextron)購自美國 GE 公司。
1.2方法
1.2.1臨床樣品中輪狀病毒的分離及 PAGE 鑒定具體方法參見文獻[5]。
1.2.2病毒的培養(yǎng)及蝕斑純化過濾后的病毒上清接種24 孔板 MA104 細胞,擴增 6 ~ 7 代后,做病毒蝕斑實驗,并進行 3 輪純化,具體的方法參見文獻[6]。
1.2.3VP8 基因的克隆及測序根據 Genbank 數據庫序列,選取人常見 P 血清型 VP8 的保守區(qū)域,設計引物如下:克隆引物 5' cttctcactaattcatattc 3'(34 ~ 53 bp),5' tatttc agaccatttataacc 3'(859 ~ 879 bp),測序引物 5' tcccaagaatc taaatgtaa 3'(sense)或 5' ttacatttagattcttggga 3'(antisense)(628 ~ –647 bp),使用抽提的輪狀病毒 RNA 為模板,做RT-PCR,將 PCR 產物克隆至 pGEM-T easy 載體后測序,測序由上海生工生物工程有限公司完成。
2 結果
2.1臨床樣品中輪狀病毒的基因型
對 30 份臨床糞便樣品編號 1 ~ 30,并進行輪狀病毒的分離,選取分離結果較好的 5 份樣品分別為 1、4、5、6、7 號進行培養(yǎng)。輪狀病毒基因組為分 11 節(jié)段的雙鏈 RNA,不同血清型的輪狀病毒 RNA 基因組在 PAGE 電泳時呈現不同的電泳特征(圖1),A 群輪狀病毒的 4 個區(qū)段中,各帶的排列位置為 4:2:3:2,根據前 4 條 RNA 的排列特征又可以進一步區(qū)分同屬 A 群但不同血清型的病毒株,Wa株(血清型為 P[8]G1)的前 4 條排列為 1-2-1,因此,初步判定 5 號、7 號樣品為 P[8]G1 型。
2.2分離輪狀病毒培養(yǎng)后的滴度及蝕斑表型
按前述方法在 MA104 細胞上進行連續(xù)擴增,臨床樣品 5 號病毒滴度最高,可達 106TCID50/ml,其 RNA 電泳條帶與 Wa 株一致,血清型對應為 P[8]G1,以 10-5稀釋度做蝕斑實驗,出現了兩種表型,一種是較大的,直徑大于5 mm,一種是較小的,直徑小于 3 mm。選取不同蝕斑表型的輪狀病毒克隆進行 3 輪純化,結果發(fā)現,蝕斑表型即蝕斑的大小與母代病毒保持一致(圖2)。

圖1 臨床糞便樣品中的輪狀病毒 RNA 電泳帶型
2.3不同蝕斑表型病毒克隆 VP8 基因測序結果分析
VP8 的 64 ~ 224 位氨基酸是與唾液酸結合的區(qū)域[7],而兩個主要的位點 157、187 是主要的糖鏈結合點[2]。對大小蝕斑的 VP8 編碼區(qū)(869 bp)進行擴增和測序,結果顯示,兩者 VP8 序列的主要差異集中在 aa 64 ~ 224 區(qū)域,
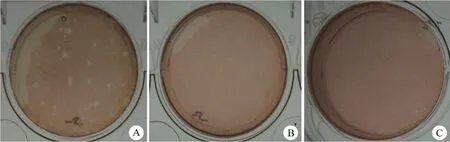
圖2 臨床樣品 5 號分離輪狀病毒經 3 輪蝕斑純化后的表型(A:直徑大于 5 mm 的大蝕斑表型;B:直徑小于 3 mm 的小蝕斑表型;C:陰性對照)
主要差異見表1。在識別多糖受體的關鍵位點 157 位,直徑較大的蝕斑為 Pro,而直徑較小的為 Ser;187 位均為Ser,但 189 位發(fā)生了 Ser 到 Asn 的變化。VP8 基因的同源性比對發(fā)現,該株病毒 VP8 基因與人 Wa株(血清型為P[8]G1)最為相近,相似度為 95%。
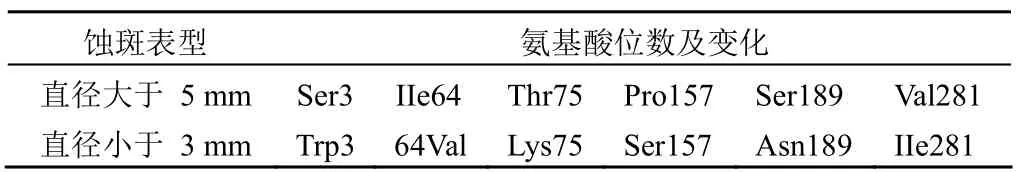
表1 臨床樣品 5 號大小蝕斑 VP8 氨基酸差異
3 討論
感染人的輪狀病毒主要是 A 群,血清組合常見的有P[8]G1、P[4]G2、P[6]G3 等,且病毒 RNA 電泳的核酸帶型有顯著的差異排布。VP4 血清型即 P 血清型特異的區(qū)域位于 VP4 的亞基 VP8,VP4 氨基酸同源性達 89% 或者更高的可判定屬于同一個 VP4 血清型[7-8]。
不同血清型的輪狀病毒感染細胞后,一般可以形成裂解性感染,產生典型的 CPE,但病毒的滴度和產生 CPE 的強度因病毒的來源和血清型不同有較大差異。這與病毒是否可以有效地結合于細胞表面并進入細胞內部完成復制有關,即病毒對細胞表面受體的識別和進入細胞的進程影響了病毒的繁殖周期和滴度水平。
輪狀病毒入侵人體并導致感染的病毒粒子和宿主細胞之間確切的相互作用特異性機制尚未闡明[1]。最近的研究顯示,輪狀病毒的外殼蛋白 VP4 經胰蛋白酶裂解后的產物VP8 對于病毒感染來說是必需的,VP8 介導了病毒與細胞膜的融合[2, 9-10]。
本研究中,發(fā)現分離自臨床糞便樣品的輪狀病毒 5 號,在蝕斑純化時呈現出兩種不同的表型,即大蝕斑和小蝕斑,通過對其 VP8 基因序列的分析發(fā)現,直徑較大的蝕斑在157 位氨基酸為 Pro,而直徑較小的蝕斑在 157 位氨基酸為 Ser,Pro 到 Ser 的突變減弱了輪狀病毒與受體的親和力[11],從而導致病毒的感染性下降,蝕斑表型變小。但這個序列的代表性還需要進一步分析其他臨床樣本,選取多個蝕斑純化病毒進行比較確證。
本文的研究同時提示,作為存在基因重排和變異較高的RNA 雙鏈病毒,輪狀病毒疫苗株的篩選,需要對來自臨床樣品的母代病毒進行多輪的蝕斑純化,得到基因遺傳穩(wěn)定,純凈的單克隆病毒。
參考文獻
[1] Blanchard H, Yu X, Coulson BS, et al. Insight into host cell carbohydrate-recognition by human and porcine rotavirus from crystal structures of the virion spike associated carbohydrate-binding domain (VP8*). J Mol Biol, 2007, 367(4):1215-1226.
[2] Stehle T, Khan ZM. Rules and exceptions: sialic acid variants and their role in determining viral tropism. J Virol, 2014, 88(14):7696-7699.
[3] Fleming FE, B?hm R, Dang VT, et al. Relative roles of GM1 ganglioside, N-acylneuraminic acids, and alpha2beta1 integrin inmediating rotavirus infection. J Virol, 2014, 88(8):4558-4571.
[4] Nishikawa K, Taniguchi K, Torres A, et al. Comparative analysis of the VP3 gene of divergent strains of the rotaviruses simian SA11 and bovine Nebraska calf diarrhea virus. J Virol, 1988, 62(11):4022-4026. [5] Yang ZQ, Huang YK, Liu X. Using optimized PAGE-silver staining technique detecting rotavirus RNA in clinical fecal samples. Chin Med Biotechnol, 2010, 5(4):308-309. (in Chinese)
楊昭慶, 黃永坤, 劉馨. 使用優(yōu)化PAGE電泳及銀染技術檢測臨床糞便樣品中的輪狀病毒RNA. 中國醫(yī)藥生物技術, 2010, 5(4):308-309.
[6] Liu X. Effect of pancreatic concentration on rotavirus plaque formation. Chin Med Biotechnol, 2008, 3(2):146-147. (in Chinese)
劉馨. 胰酶濃度對輪狀病毒蝕斑形成的影響. 中國醫(yī)藥生物技術, 2008, 3(2):146-147.
[7] Rubio RM, Mora SI, Romero P, et al. Rotavirus prevents the expression of host responses by blocking the nucleocytoplasmic transport of polyadenylated mRNAs. J Virol, 2013, 87(11):6336-6345. [8] Gorziglia M, Larralde G, Kapikian AZ, et al. Antigenic relationships among human rotaviruses as determined by outer capsid protein VP4. Proc Natl Acad Sci U S A, 1990, 87(18):7155-7159.
[9] Estes MK, Cohen J. Rotavirus gene structure and function. Microbiol Rev, 1989, 53(4):410-449.
[10] Hu L, Crawford SE, Czako R, et al. Cell attachment protein VP8* of a human rotavirus specifically interacts with A-type histo-blood group antigen. Nature, 2012, 485(7397):256-259.
[11] Yu X, Dang VT, Fleming FE, et al. Structural basis of rotavirus strain preference toward N-acetyl- or N-glycolylneuraminic acid-containing receptors. J Virol, 2012, 86(24):13456-13466.
·讀者·作者·編者·
收稿日期:2015-11-10
通信作者:劉馨,Email:fluidstar@126.com
DOI:10.3969/j.issn.1673-713X.2016.01.015
