不同碳源條件下PQQ對植物促生菌Rahnella aquatilis HX2溶解無機磷影響的研究
2016-03-17 05:03:21焦子偉張相鋒吾爾恩郭巖彬
新疆農業科學 2016年2期
焦子偉,張相鋒,張 娜,吾爾恩,郭巖彬
(1. 伊犁師范學院化學與生物科學學院,新疆伊寧 835000;2. 中國農業大學資源與環境學院,北京 100193)
?
不同碳源條件下PQQ對植物促生菌RahnellaaquatilisHX2溶解無機磷影響的研究
焦子偉1,張相鋒1,張 娜1,吾爾恩1,郭巖彬2
(1. 伊犁師范學院化學與生物科學學院,新疆伊寧835000;2. 中國農業大學資源與環境學院,北京100193)
摘要:【目的】水生拉恩氏菌(Rahnella aquatilis)HX2能合成葡萄糖脫氫酶(glucose dehydrogenase, GDH)和吡咯喹啉醌(pyrroloquinoline quinone,PQQ),PQQ為GDH輔酶,與其共同參與葡萄糖溶磷代謝。揭示不同碳源條件下PQQ影響HX2菌株溶解無機磷作用的機理。【方法】以野生菌株HX2、突變體MH15及其互補菌株CMH15(pqq)為供試材料,在不同碳源條件下采用平板溶磷、鉬銻抗比色法等方法對其進行溶解無機磷定性、定量以及pH相關分析。【結果】除D-山犁醇、D-果糖外,PQQ參與HX2菌株對木糖、葡萄糖、D-甘露糖、D-甘露醇、蔗糖、乳糖6種碳源的溶磷代謝,但其溶磷代謝因HX2菌株利用不同碳源的能力而不同,以乳糖利用最低,木糖利用最高。【結論】PQQ作為GDH的輔酶,兩者共同參與HX2菌株的木糖、葡萄糖、D-甘露糖、D-甘露醇、蔗糖和乳糖溶解無機磷代謝,并起到重要調控作用。
關鍵詞:吡咯喹啉醌;水生拉恩氏菌HX2;不同碳源;無機磷
0 引 言
【研究意義】RahnellaaquatilisHX2菌株已作為優良的溶磷促生菌應用于伊犁河谷綠色有機農業生產中[1],進一步明確其溶磷機理可對其促進植物生長提供理論依據和技術支持。【前人研究進展】 HX2菌株具有較強的溶磷能力,可利用不同碳源的培養基生長,產生PQQ和GDH,PQQ作為GDH的輔酶,兩者共同參與其葡萄糖溶磷代謝[2]。焦子偉等[1]已明確在不同碳源溶磷培養基條件下,GDH參與了HX2菌株對木糖、萄萄糖、D-甘露糖、D-甘露醇、乳糖、蔗糖、D-果糖D-山梨醇溶磷代謝。【本研究切入點】Guo等[3]已通過Tn5插入突變pqq基因獲得了突變菌株MH15,以及其互補菌株CMH15(pqq)。研究在已有基礎上,采用平板溶磷、鉬銻抗比色法等實驗方法,分析研究不同碳源下PQQ對HX2菌株溶解無機磷能力的影響。【擬解決的關鍵問題】明確PQQ在不同碳源下對HX2菌株溶磷的作用與代謝,揭示PQQ作為GDH的輔酶共同參與對HX2菌株溶解無機磷作用影響機理。
1材料與方法
1.1 材 料
1.1.1 菌 株
RahnellaaquatilisHX2,Tn5插入突變pqq基因的突變體MH15,MH15的互補菌株CMH15(pqq),列出具體特性。表1
表 1 菌株及質粒特征
Table 1 Characteristis of bacteria strains and plasmids

名稱Names特征Characteristics文獻來源Literatureresources水生拉恩氏菌RahnellaaquatilisHX2Apr,WildtypeChen[2]MH15Apr,Kmr,HX2derivativecontainingaTn5insertioninpqqEgeneGuo[3]CMH15(pqq)Apr,Kmr,Tcr,MH15containingplasmidpCH15withthepqqgenesGuo[3]質粒PlasmidsCP465Tcr,pLAFR-5containingpqqgenes,cosmidGuo[3]pCH15Gmr,Tcr,pRK415Gcontainingapproximately8.0kbBamHIfragmentincludingpqqgenesfromcosmidCP465Guo[3]
注:Apr, Kmr, Gmrand Tcr分別表明抗氨芐青霉素、卡那霉素、慶大霉素和四環素
Note: Apr, Kmr, Gmrand Tcrindicate resistance to ampicillin, kanamycin, gentamicin and tetracycline, respectively
1.1.2 培養基
采用國際植物研究所磷酸鹽生長培養基(NBRIP)、不同碳源NBRIP培養基和LB液體培養基參照焦子偉等[1]配方。
1.2 方 法
1.2.1碳源選擇
根據前期已有研究結果[1],選用蔗糖、乳糖、D-甘露糖、D-甘露醇、D-山梨醇、木糖、D-果糖和葡萄糖(作為對照)8種碳源進行溶解無機磷的定性、定量分析。
1.2.2 溶磷定性檢測
將HX2、MH15、CMH15(pqq)菌株活化、搖培后,將菌懸液接種于含有直徑為0.5 cm的濾紙片的不同碳源NBRIP固體培養基平板上,并將各處理樣品進行培養,具體操作與檢測分析參照焦子偉等[1]溶磷平板定性檢測方法。
1.2.3 有效磷定量、pH檢測
將HX2、MH15、CMH15(pqq)菌株活化、搖培后,將菌懸液接種到不同碳源NBRIP液體培養基中進行培養,將所有處理樣品于不同搖培時間分別抽取搖培液進行離心取上清液,進行有效磷定量和pH的檢測,具體操作與檢測參照焦子偉等[1]有效磷定量和pH值檢測方法。
1.3 數據統計
采用EXCEL軟件進行數據分析處理和SPSS軟件方差分析(ANOVA)。
2結果與分析
2.1 不同碳源對HX2、MH15、CMH15菌株溶磷定性檢測
在8種不同碳源作為NBRIP固體培養基的碳源,經7 d培養,HX2、MH15、CMH15(pqq)菌株都有溶磷現象,溶磷圈直徑也各不相同。同一碳源溶磷培養基培養情況下,各菌株在D-果糖、D-山犁醇作為碳源情況下,其溶磷圈直徑無顯著差異;而在其它不同6種(木糖、蔗糖、乳糖、D-甘露糖、D-甘露醇和對照)碳源培養基情況下,其溶磷圈直徑差異顯著(P<0.05)。以D-山犁醇為例,其溶磷圈直徑最小,HX2 、MH15、CMH15(pqq)菌株溶磷直徑在1.35~1.43 cm;木糖、D-甘露糖和對照作為碳源,其溶磷圈直徑較大,如在D-甘露糖作為碳源情況下,HX2、CMH15(pqq)溶磷圈直徑分別為2.21、2.15 cm;突變體MH15溶磷圈直徑1.10 cm,缺失pqq基因的菌株溶磷圈直徑明顯減小。圖1,表2

注:不同碳源分別為A,木糖; B, D-甘露糖; C, D-甘露醇; D, D-山犁醇; E, 乳糖; F, 蔗糖; G, D-果糖;H,對照(葡萄糖);a, HX2; b, MH15; c, CMH15(pqq) ;下同
Note: Different carbon resources: A, Xylose; B, D-Mannose; C, D-Mannitol; D, D-Sorbitol; E, lactose; F, Surcose; G, D-fructose; H, control (glucose). a, HX2; b, MH15; c, CMH15(pqq);the same as bolow
圖1 不同處理和培養基下菌株溶磷圈
Fig.1 Clear halo of solubilization of tricalcium phosphate by strains under different treatment and medium
表2不同處理和培養基下菌株溶磷圈直徑 (cm)
Table 2 Phosphate-solubilizing halo diameter (cm) produced by strains under different treatment and medium
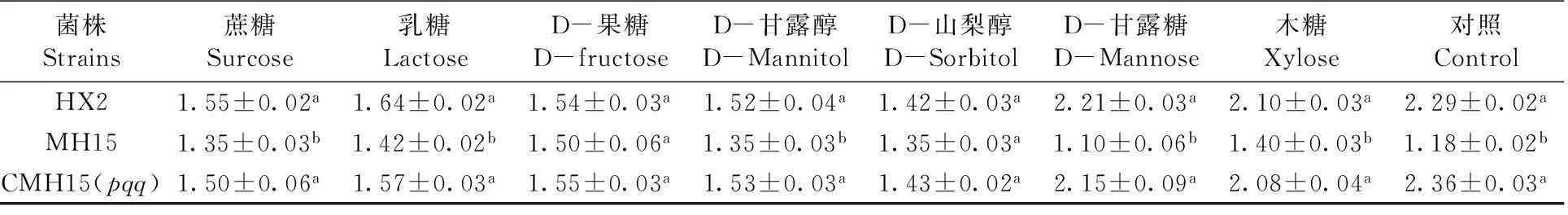
菌株Strains蔗糖Surcose乳糖LactoseD-果糖D-fructoseD-甘露醇D-MannitolD-山梨醇D-SorbitolD-甘露糖D-Mannose木糖Xylose對照ControlHX21.55±0.02a1.64±0.02a1.54±0.03a1.52±0.04a1.42±0.03a2.21±0.03a2.10±0.03a2.29±0.02aMH151.35±0.03b1.42±0.02b1.50±0.06a1.35±0.03b1.35±0.03a1.10±0.06b1.40±0.03b1.18±0.02bCMH15(pqq)1.50±0.06a1.57±0.03a1.55±0.03a1.53±0.03a1.43±0.02a2.15±0.09a2.08±0.04a2.36±0.03a
數據為平均值和標準誤差。(a-b),不同字母表示為顯著性差異(P<0.05)
Data was shown by Mean (± standard error) values. (a-b), different letters indicated statistically significant (P< 0.05)
2.2 不同碳源對HX2、MH15、CMH15菌株的pH檢測
8種碳源分別作為不同碳源NBRIP培養基,經7 d培養,HX2、MH15、CMH15(pqq)菌株在不同碳源NBRIP溶磷培養基上均能生長,但隨著培養時間延長,pH均逐漸下降,但pH各不相同,最終相對趨于穩定。除D-果糖、D-山犁醇作為碳源溶磷培養基外,HX2、CMH15(pqq)與突變體MH15處理的pH值都有顯著性差異(P<0.05);以D-甘露糖、對照(葡萄糖)和木糖作為碳源培養基,HX2及其衍生菌株pH值變化較大。如木糖作為碳源的NBRIP培養基,HX2、CMH15(pqq)的pH分別為3.28、3.26,MH15突變體的pH 4.53,明顯增高。表3,圖2
表3 不同處理和培養基下菌株pH值
Table 3 pH observed by strains under different treatment and medium
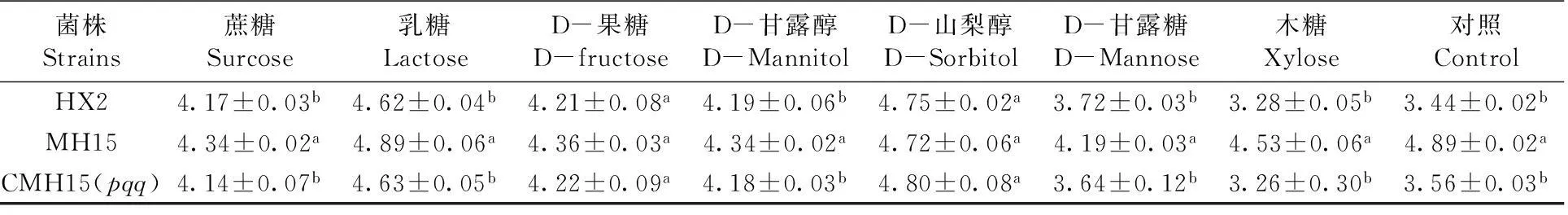
菌株Strains蔗糖Surcose乳糖LactoseD-果糖D-fructoseD-甘露醇D-MannitolD-山梨醇D-SorbitolD-甘露糖D-Mannose木糖Xylose對照ControlHX24.17±0.03b4.62±0.04b4.21±0.08a4.19±0.06b4.75±0.02a3.72±0.03b3.28±0.05b3.44±0.02bMH154.34±0.02a4.89±0.06a4.36±0.03a4.34±0.02a4.72±0.06a4.19±0.03a4.53±0.06a4.89±0.02aCMH15(pqq)4.14±0.07b4.63±0.05b4.22±0.09a4.18±0.03b4.80±0.08a3.64±0.12b3.26±0.30b3.56±0.03b
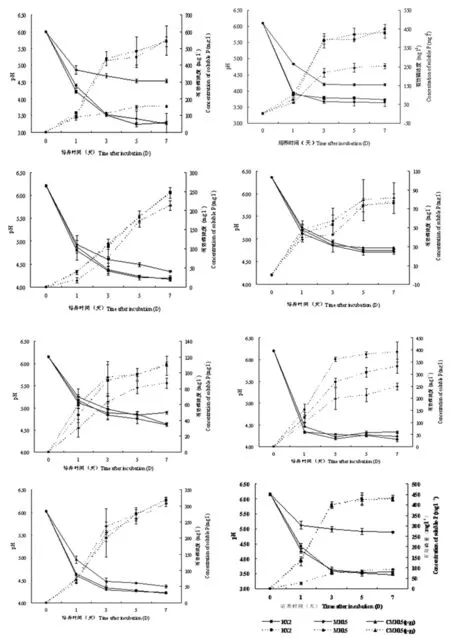
注:虛線部分代表有效磷濃度變化圖;實線部分代表pH值變化
Note:pH of NBRIP after incubation was signed with the real line. Concentration of soluble-P was indicated by broken line
圖2不同處理和培養基下菌株pH和產有效磷變化曲線
Fig. 2Change curve of pH and concentration of soluble-P produced by strains under different treatment and medium
2.3 不同碳源對HX2、MH15、CMH15菌株溶磷定量檢測
HX2、MH15、CMH15(pqq)菌株在8種不同碳源溶磷培養基上,經7 d培養,隨著培養時間延長,有效磷濃度逐漸上升,最終趨于相對穩定;各菌株均都能產生有效磷,但產生有效磷的濃度各不相同。同一碳源NBRIP培養情況下,D-果糖和D-山犁醇作為碳源下,各菌株之間產生有效磷濃度差異不顯著;其余的如蔗糖、乳糖、D-甘露醇、D-甘露糖、木糖和對照作為碳源NBRIP培養基,野生菌株HX2、CMH15(pqq)與插入突變pqq基因的菌株MH15之間產生的有效磷濃度差異顯著(P<0.05)。各菌株在D-山犁醇作為碳源溶磷培養基中,產生的有效磷濃度最小;在木糖中產生的最大,其次是葡萄糖和D-果糖。以木糖為例,HX2、CMH15(pqq)菌株產生的有效磷濃度分別為541.26和547.32 mg/L;MH15突變體產生的有效磷濃度為154.29 mg/L,產生的有效磷濃度最低。表4,圖2
表4不同處理和培養基下菌株產有效磷量
Table 4 Concentration of soluble-P produced by strains under different treatment and medium(mg/L)
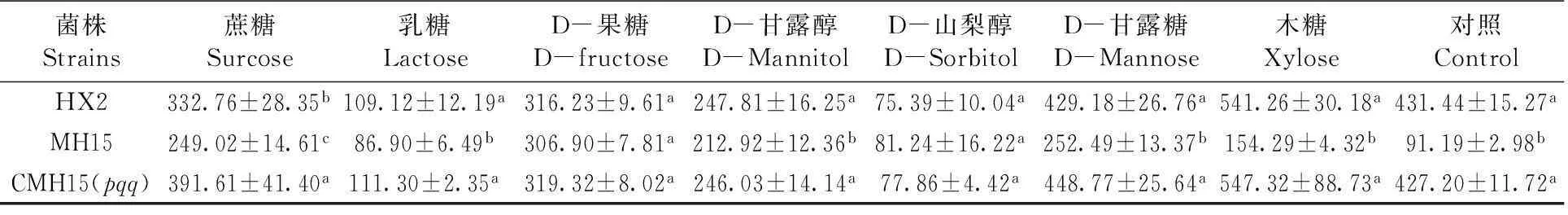
菌株Strains蔗糖Surcose乳糖LactoseD-果糖D-fructoseD-甘露醇D-MannitolD-山梨醇D-SorbitolD-甘露糖D-Mannose木糖Xylose對照ControlHX2332.76±28.35b109.12±12.19a316.23±9.61a247.81±16.25a75.39±10.04a429.18±26.76a541.26±30.18a431.44±15.27aMH15249.02±14.61c86.90±6.49b306.90±7.81a212.92±12.36b81.24±16.22a252.49±13.37b154.29±4.32b91.19±2.98bCMH15(pqq)391.61±41.40a111.30±2.35a319.32±8.02a246.03±14.14a77.86±4.42a448.77±25.64a547.32±88.73a427.20±11.72a
3討 論
許多研究表明一些革蘭氏陰性菌如Pseudomonascepacia[4]、Rahnellaaquatilis[5]和Enterobacterintermedium60-2G[6]的PQQ 合成酶的pqq基因決定其溶解無機磷的能力[7]。野生菌株HX2在以葡萄糖、D-甘露醇、乳糖、D-甘露糖等為碳源的培養基中均可正常生長[2]。研究以野生菌株HX2為供試研究對象,除葡萄糖作為NBRIP的碳源之外,把乳糖、蔗糖、木糖、D-甘露醇、D-甘露糖、D-山梨醇、D-果糖來作為NBRIP不同的7種碳源,進行其溶磷定性、定量分析,HX2、CMH15(pqq)與MH15菌株之間在蔗糖、乳糖、D-甘露糖、D-甘露醇、木糖和葡萄糖6種碳源NBRIP培養基上產生的有效磷濃度差異顯著,這說明插入突變pqq的突變體很難利用不同碳源溶解難溶性的無機磷,也得出了類似的結果。
PQQ作為GDH等脫氫酶的輔酶,對GDH的功能發揮起關鍵控制作用,溶磷細菌通過GDH-PQQ 全酶作用能溶解土壤無機或有機磷酸鹽,促進植物對營養的攝入和植物生長[4,6,8,9]。Rodríguez等[10]研究報道導入pqq基因的BurkholderiacepaciaIS-16 和假單孢菌屬(Pseudomonassp)兩個菌株能增強礦質磷酸鹽溶解表型,促進對磷的溶解。野生菌株HX2既能合成GDH和PQQ,在HX2菌株中,GDH都參與木糖、萄萄糖、D-甘露醇、D-甘露糖、乳糖、蔗糖、D-山梨醇、D-果糖溶磷代謝[1];PQQ參與木糖、葡萄糖、D-甘露醇、D-甘露糖、蔗糖、乳糖溶磷代謝,不參與D-山梨醇、D-果糖溶磷代謝;這說明PQQ作為GDH的輔酶參與了HX2菌株的木糖、葡萄糖、D-甘露糖、D-甘露醇、蔗糖、乳糖的溶磷代謝。這也可能說明了GDH有更好的底物特性,PQQ作為GDH重要的輔酶,但不是唯一的,PQQ的結構類似物也可作為GDH氧化還原酶的輔酶或輔助因子參與溶磷代謝[11]。
MH16、MH15突變體在不同碳源溶磷培養基條件下也能產生有效磷,說明除葡萄糖溶磷主要機制外,可能還有其它的溶磷機制也參與HX2菌株的溶磷代謝。關于PQQ作為GDH的輔酶參與其溶磷產酸的調控機理,以及該菌株涉及的其它相關溶磷機理還有待進一步深入研究。
4結 論
從PQQ參與不同碳源條件下對HX2菌株溶解無機磷作用影響結果分析,HX2菌株在8種不同碳源上具有不同的溶磷效果,PQQ除D-山梨醇、D-果糖外均參與該菌株對木糖、葡萄糖、D-甘露糖、D-甘露醇、蔗糖和乳糖溶磷代謝,以乳糖利用最低,木糖利用最高。PQQ作為GDH的輔酶,共同參與HX2菌株的木糖、葡萄糖、D-甘露糖、D-甘露醇、蔗糖和乳糖溶磷代謝,并起到重要調控作用。
參考文獻(References)
[1]焦子偉, 吳文良, 郭巖彬. 不同碳源條件下GDH對植物促生菌HX2溶解無機磷影響的研究[J]. 新疆農業科學, 2015,52(2):268-274.
JIAO Zi-wei, WU Wen-liang, GUO Yan-bin. (2015). Effect of glucose dehydrogenase on mineral phosphate solubilization with different carbon sources in Rahnella aquatilis HX2 [J].XinjiangAgriculturalSciences, 52(2):268-274. (in Chinese)
[2]陳凡. 水生拉恩氏菌HX2菌株防治葡萄根癌病的初步研究[D]. 北京:中國農業大學博士論文,2007.
CHEN Fan. (2007).PrimarystudiesonbiologicalcontrolofgrapevinecrowngallbyRahnellaaquatilisHX2 [D]. PhD Dissertation. China Agriuclture University, Beijing. (in Chinese)
[3]Yan Bin, G., Jinyun, L., Lei, L., Fan, C., Wenliang, W., & Jianhui, W., et al. (2009). Mutations that disrupt either the pqq or the gdh gene of rahnella aquatilis abolish the production of an antibacterial substance and result in reduced biological control of grapevine crown gall.Applied&EnvironmentalMicrobiology, 75(21):6,792-6,803.
[4]Babukhan, S., Yeo TCMartin, W. L., Duron, M. R., Rogers, R. D., & Goldstein, A. H. (1995). Cloning of a mineral phosphate-solubilizing gene from pseudomonas cepacia.Applied&EnvironmentalMicrobiology, 61(3):61--972.
[5]Kim, K. Y., Nald, G. A., & Jordan, D. (1997). Solubilization of hydroxyapatite by enterobacter agglomerans and cloned escherichia coli in culture medium.Biology&FertilityofSoils, 24(4):347-352.
[6]Kim, C. H., Song, H. H., Kim, K. Y., Cho, B. H., Yong, H. K., & Koo, B. S., et al. (2003). Cloning and expression of pyrroloquinoline quinone (pqq) genes from a phosphate-solubilizing bacterium enterobacter intermedium.CurrentMicrobiology, 47(6):457-461.
[7]Vikram, A., Alagawadi, A. R., Krishnaraj, P. U., & Kumar, K. S. M. (2007). Transconjugation studies in azospirillum sp. negative to mineral phosphate solubilization.WorldJournalofMicrobiology&Biotechnology, 23(9):1,333-1,337.
[8]Han, S. H., Kim, C. H., Lee, J. H., Ju, Y. P., Song, M. C., & Park, S. K., et al. (2008). Inactivation of pqq genes of enterobacter intermedium 60-2g reduces antifungal activity and induction of systemic resistance.FemsMicrobiologyLetters, 282(1):140-146.
[9]Liu, S. T., Lee, L. Y., Tai, C. Y., Hung, C. H., Chang, Y. S., & Wolfram, J. H., et al. (1992). Cloning of an erwinia herbicola gene necessary for gluconic acid production and enhanced mineral phosphate solubilization in escherichia coli hb101.JournalofBacteriology, 174(18):5,814-5,819.
[10]Rodríguez, H., Gonzalez, T., & Selman, G. (2000). Expression of a mineral phosphate solubilizing gene from erwinia herbicola in two rhizobacterial strains.JournalofBiotechnology, 84(2):155-161.
[11]周怡雯,陳建華. 新輔酶吡咯喹啉醌研究進展. 中國生化藥物雜志,2008,29(4):279-282.
ZHOU Yi-wen, CHEN Jian-hua. (2008).ProgressintheresearchofpyrroloquinolinequinoneChineseJournalofBiochemicalPharmaceutics, 29(4):279-282. (in Chinese).
Fund project:Supported by University Scientific Research Projects of Xinjiang Uygur Autonomous Region (XJEDU2014I041) and NSFC (31200386)
Influences of Pyrroloquinoline Quinone on Inorganic Phosphate
Solubilization under Different Carbon Sources in
RahnellaaquatilisHX2
JIAO Zi - wei1, ZHANG Xiang - feng1, ZHANG Na1, Wueren1, GUO Yan- bin2
(1.CollegeofChemistryandBiologicalSciences,YiliNormalUniversity,YiningXinjiang835000,China; 2.DepartmentofEcologyandEcologicalEngineering,CollegeofResourcesandEnvironmentalSciences,ChinaAgriculturalUniversity,Beijing100193,China)
Abstract:【Objective】 Rahnella aquatilis strain HX2 can produce glucose dehydrogenase (GDH) and pyrroloquinoline quinone (PQQ). PQQ as coenzyme of GDH jointly take part in metabolism of phosphate solubilization under glucose source in HX2 strain. Based on prior researches, this paper further revealed effects of PQQ on inorganic phosphate solubilization under different carbon source conditions in HX2 strain. 【Method】 HX2 wild strains, mutant MH15 and its complementary strains were used as test material. The methods of solubilzing phospate on plate, molybdenum-blue method and so on were used to analyze the qualitative and quantitative ability of phosphate solubilization under different carbon sources. 【Result】Except D-sorbitol and D-fructose, PQQ played a key role in HX2 phosphate solubilization with glucose, xylose, D-mannose, D-mannitol, sucrose and lactose as a sole carbon sources. However, its ability of phosphate solubilization was different with different carbon sources, which was the lowest for lactose and the highest for xylose as carbon sources. 【Conclusion】 It has been clear that PQQ and GDH as holoenzyme were involved in metabolism of phospate solubilization under different carbon sources such as glucose, xylose, D-mannose, D-mannitol, sucrose and lactose, and played an important regulatory role in HX2 strain.
Key words:PQQ; Rahnella aquatilis HX2; Carbon sources; Inorganic phosphate
通訊作者:郭巖彬(1978-),男,博士,副教授,研究方向為植生物生態學、土壤微生物、有機農業,(E-mail)guoyb@cau.edu.cn
作者簡介:焦子偉(1973-),男,博士,副教授,研究方向為微生物生態及綠色有機農業有害生物綜合防控,(E-mail)741285332@qq.com
基金項目:自治區高校科研計劃項目(XJEDU2014I041);國家自然科學基金項目(31200386)
收稿日期:2015-09-09
中圖分類號:S188+.4
文獻標識碼:A
文章編號:1001-4330(2016)02-0295-07
doi:10.6048/j.issn.1001-4330.2016.02.015
