秘魯鳀資源變動及與海洋環境要素的關系研究進展
2016-03-09 09:13:46汪金濤陳新軍
海洋漁業 2016年2期
關鍵詞:資源
陳 芃,汪金濤,陳新軍
(1.上海海洋大學海洋科學學院,上海 201306;2.國家遠洋漁業工程技術研究中心,上海 201306;3.大洋漁業資源可持續開發省部共建教育部重點實驗室,上海 201306;4.遠洋漁業協同創新中心,上海 201306)
·綜述·
秘魯鳀資源變動及與海洋環境要素的
關系研究進展
陳 芃1,4,汪金濤1,4,陳新軍1,2,3,4
(1.上海海洋大學海洋科學學院,上海 201306;2.國家遠洋漁業工程技術研究中心,上海 201306;3.大洋漁業資源可持續開發省部共建教育部重點實驗室,上海 201306;4.遠洋漁業協同創新中心,上海 201306)
秘魯鳀(Engraulis ringens)是一種小型中上層魚類。作為重要的商業性魚類之一,秘魯鳀的捕撈產業曾形成了世界上最大的單魚種漁業,但其產量的年間差異非常大,上升流流場結構變化是其產量變化的重要影響因素。上升流對秘魯鳀捕撈量的作用機制可歸納為低緯度的地理位置、適宜的水溫結構、低溶解氧、高能量傳遞效率的食物網以及復雜的海洋環境要素變化五個方面。秘魯鳀漁業生物學的多個方面都顯著地受到了海洋環境變化的影響。此外,與氣候相關的大尺度海洋生態系統周期性變化(regime shift)也影響到了秘魯鳀的資源變動。本研究認為,今后在加強對長時間尺度生態系統周期性變化的研究同時,也應注重結合海洋環境變化、捕撈因素及種群的內部動力過程這三者之間的關系,結合基于個體的海洋動力學,建立秘魯鳀資源評估及預測的模型,為合理開發和管理提供基礎。
秘魯鳀;海洋環境;資源變動;秘魯上升流;氣候變化
秘魯鳀(Engraulis ringens)是一種小型中上層魚類[1-2],作為重要的商業性魚類之一,秘魯鳀的捕撈業曾是世界上最大的單魚種漁業[3]。但其產量的年間差異也非常大,產量最高的年份(1970年)與最低的年份(1984年)之間相差了1 305×104t(圖1),2000年以后的產量維持在500×104t的水平[4]。研究發現,秘魯鳀的資源變動與海洋環境甚至氣候因素都有非常大的關系[5]。為此,本文從秘魯鳀棲息環境的特點及其對秘魯鳀捕撈量的作用機制出發,闡述秘魯鳀與海洋環境及氣候變化之間的關系,為合理開發和利用該資源提供依據。
1 秘魯鳀的棲息環境
1.1 秘魯上升流系統
秘魯鳀棲息于東南太平洋南美洲4°30'S~42°30'S的西部沿岸30 nmile內海域[2,6]。該海域存在著世界上著名的東部上升流——秘魯上升流(該區域又被稱為洪堡海流系統,Humboldt current system,HCS)[7]。
上升流是一種典型的海洋現象,指水體從次表層向表層的上升運動,并被水平流帶到其它流域[8]。秘魯上升流的形成與海域表面吹向赤道方向的信風(trade wind)、海水水柱的層化、沿岸地形和地球自轉有關,其中信風是最主要的原因,信風能夠產生離岸的水體埃克曼輸送(Ekman transport)和由下層至表層的水體埃克曼抽吸(Ekman pumping),由此產生了上升流[7-9]。根據上升流的強弱及海域生產力的大小,秘魯上升流在緯度方向上可以分成以下三個主要區域:(1)南部智利季節性(夏季)上升流(30°S~40°S);(2)南部秘魯和北部智利較弱上升流(18°S~26°S);(3)北部秘魯的終年上升流(4°S~16°S)[7]。
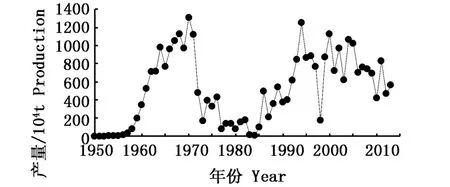
圖1 1950~2013年世界秘魯鳀產量變化[4]Fig.1 Global production of anchoveta(Engraulis ringens)from 1950 to 2013
秘魯上升流海域的海洋要素組成十分復雜(圖2),在南部(45°S),洪堡海流(Humboldt current,又稱為秘魯海流,Peru current)沿著東部岸界向赤道方向輸送冷的亞極地表層水(sub-Antarctic surface water,SASW);SASW從18°S開始與高溫高鹽的亞熱帶表層水(subtropical surface water,SSW)混合;在北部,高營養鹽的上升流冷水與熱帶表層水(tropical surface water,TSW)延伸的赤道表層水(equatorial surface water,ESW)也形成了混合區域;與秘魯海流方向相反,在表層循環或水團之下還存在著由赤道潛流(equatorial undercurrent)產生的極地方向的秘魯-智利潛流(Peru-Chile undercurrent,PUC)[10]。渦等中小尺度海洋現象也常在該海域中發生:CHAIGNEAU等[11]發現,秘魯欽博特(Chimbote,9°S)及15°S~19°S的沿岸附近經常有中尺度渦(約為 3個月的時間周期)的存在;HORMáZABAL等[12]發現,在中部智利(29°S~39°S)沿岸至離岸600~800 km的海域存在著與渦和水舌相關的高渦動能(eddy kinetic energy)的沿岸轉換帶(coast transition zone,CTZ)。而在秘魯沿岸(10°S~15°S)和北部智利沿岸(19°S~ 29°S)也都有CTZ的發現[12-13]。
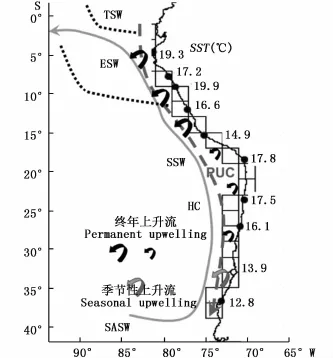
圖2 秘魯上升流區域表層海洋要素狀況[10]Fig.2 Oceanic conditions of the Peru upwelling surface region
1.2 秘魯上升流對秘魯鳀捕撈量的作用機制
早在1906年,NATHANSOHN就提出“上升流流域,一般生產力高,因而形成了優良的漁場”的論斷[14]。然而研究發現(表1)[7,15],對比非洲西北上升流及本格拉上升流,秘魯上升流的潛在新生產力與這兩個海域相差不大,而衛星監測的非洲西北上升流的初級生產力比秘魯上升流要高許多,但是在這兩個海域,不管是鳀魚類還是其它商業性開發魚類的漁業,卻沒有形成像秘魯鳀這樣在世界范圍內單一魚種產量最大的漁業。因此本研究針對秘魯上升流對秘魯鳀捕撈量的作用機制歸納出以下五點。
1.2.1 低緯度的地理位置
秘魯上升流低緯度的地理位置有益于營養鹽被生物利用。上升流到達海水表面的同時也伴隨著海水的離岸埃克曼輸送,若是輸送的速度過大或者由于海水表面風速過強導致海水湍流(turbulent)過強將不利于營養物質的聚集和生物的利用。CHAVEZ等[7]比較了世界四大東部邊界上升流的湍流強度(表1),發現在四個上升流系統中,秘魯上升流的湍流強度是最弱的。BAKUN等[16]研究發現,海水粒子在沿岸的平均滯留時間(residence times of water parcels)與埃克曼層的深度和羅士貝變形半徑(Rossby radius of deformation,可以看作沿岸棲息地的范圍)成正比,與埃克曼的水流輸送成反比。而在低緯度的秘魯上升流海域,埃克曼層的深度和羅士貝變形半徑比高緯度要大,赤道附近風速較小,由風產生埃克曼的水流輸送速度也比高緯度小,較長的海水粒子滯留時間意味著營養物質能夠富集,從而有于秘魯鳀及其餌料生物的聚集和利用。
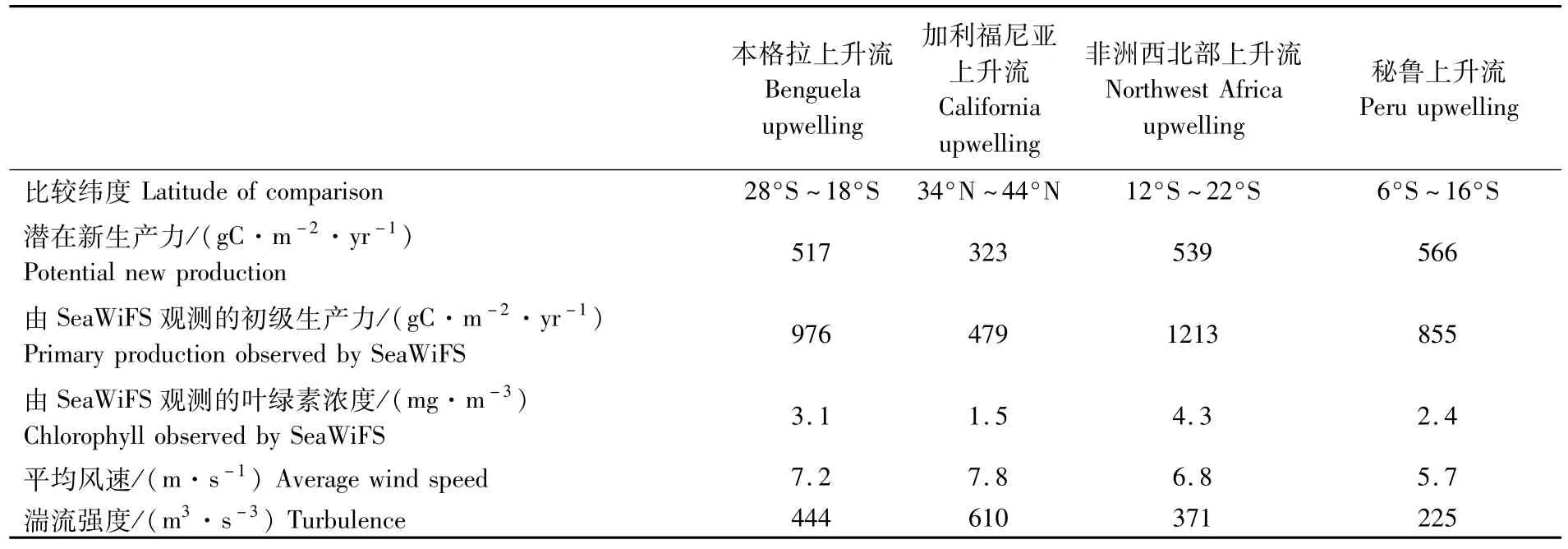
表1 沿岸0~150 km 10°緯度范圍世界四大東部上升流平均理化性質比較[7]Tab.1 Comparison of annual mean properties in a 10°latitude coastal(0-150 km)band for the four large Eastern boundary upwelling
1.2.2 適宜的水溫結構
水溫結構對秘魯鳀的資源變動有著重要的影響。研究表明[17],作為一種冷水性種類,秘魯鳀更喜歡棲息于冷的沿岸上升流水(upwelled cold coastal water,CCW)和沿岸亞熱帶表層水(mixed coastal-subtropical water,MCS)中,上升流形成的冷水區域為秘魯鳀提供了廣泛的棲息環境。從氣候變化的角度也可以看出水平方向上的水溫結構變化對秘魯鳀的資源變動有影響:秘魯上升流海域也是厄爾尼諾(El Ni?o)事件直接作用的區域[18]。El Ni?o發生時,海水表面溫度(sea surface temperature,SST)的異常偏高使得喜歡棲息于冷水區域的秘魯鳀更靠近南部及近岸偏移,棲息地的減少加劇了種群內部的競爭,同時也便于天敵的捕食和人類捕撈[19-20]。此外,垂直方向上的水溫結構變化也會影響到秘魯鳀的資源變動:上升流使得秘魯海域的溫躍層較淺[7];而研究發現[21],東南太平洋秘魯沿岸的海水營養鹽躍層與溫躍層的深度基本一致,El Ni?o現象會導致海域上升流變弱或消失,加深了海水溫躍層和營養躍層的深度,從而也減少了深海向海水表面營養鹽的供給,這對秘魯鳀的生長、成活和繁殖成功率是不利的。
1.2.3 低溶解氧
海水中溶解氧的含量會對熱帶東南太平洋沿岸區域的生物群落有著重要影響[22]。上升流導致的海水涌升使得下層未飽和的貧氧水到達表層。BERTRAND等[23]發現,上升流導致的低溶解氧濃度及氧氣最小區域(oxygen minimum zone,OMZ)的水深變淺對秘魯鳀的資源豐度起到了有益的作用。原因有以下兩點:第一,秘魯鳀對OMZ水深變淺的反應不是太敏感[24],氧氣對魚類的供給量與其形體大小有關,秘魯鳀作為一種小型魚類對氧氣的需求沒有其它大型魚類大,同時在OMZ附近生存有利于其對天敵的躲避[21];第二,秘魯鳀主要依靠視覺攝食大型浮游動物[25]。研究發現,在秘魯沿岸作為其餌料的大型浮游動物中,有79%的種類都有著與OMZ相關的晝夜垂直移動行為,白天分布在氧躍層(oxycline)下方的OMZ地帶,夜晚移動至表層進行攝食,因此淺的OMZ水深減少了餌料生物的棲息范圍從而加大了秘魯鳀的攝食機會[23,26]。
1.2.4 具有高能量傳遞效率的食物網
ESPINOZA等[25]對秘魯鳀的胃含物進行分析,發現其中99.52%的成分都是浮游植物,但是從食物的碳含量組成(即其主要的能量來源)來看,浮游動物提供了其能量來源的98%,在這之中,大型浮游動物磷蝦類(euphausiids,67.5%)和橈足類(copepods,26.3%)占了絕大部分,這表明直接利用初級生產力的浮游植物并不是支配秘魯鳀能量供應的主要來源,大型浮游動物的變化對其資源變動有著重要的影響。AYóN等[27]分析了1961~2005年浮游動物生物量數據與秘魯鳀上岸量之間的關系發現,浮游動物的資源變化與秘魯鳀的資源波動基本保持同樣的趨勢;而ALHEIT等[19]發現,1970年秘魯鳀漁業崩潰的前兩年,海域內浮游動物生物量就已經發生下降,而1985年開始秘魯鳀的資源恢復也與大型橈足類資源恢復有關。因此,從食物網結構上看,秘魯鳀與初級生產者之間相隔著浮游動物一個營養級卻形成較高的資源豐度,這與秘魯上升流生態系統食物網較高的能量傳遞效率有著密切的聯系[25,28]。TAM等[29]使用Ecopath模型對比了1997~1998年El Ni?o現象(上升流變弱)和1995~1996年拉尼娜(La Ni?a)現象(上升流增強)發生時北部秘魯上升流生態系統的食物網能量流動情況,發現上升流變弱時,生態系統的食物網結構縮小并且其內部間的能量流動也減弱,上升流增強時則情況相反。綜上所述,秘魯上升流的增強能夠提升海域內食物網的能量傳遞效率。
1.2.5 復雜的海洋環境要素變化
秘魯上升流海域受到了多種尺度的海洋環境要素變化的控制[10],如開爾文波(Kelvin wave)[30]、羅士貝波(Rossby waves)[16]、ENSO現象[18]和數十年尺度的太平洋年代際振蕩(Pacific Decadal Oscillation,PDO)[5],此外還有空間上中小尺度的海洋要素變化[31]。秘魯鳀作為一種快速生長和成熟的魚類,被認為是一種r選擇型種類[10,15],其資源能在環境利好的情況下快速恢復。同時復雜的海洋環境要素變化也使得需要穩定環境、生存時間較長、高營養級的秘魯鳀天敵們對該海域保持“遠離”[16]。
2 海洋環境與秘魯鳀的漁業生物學
海洋環境的變化不僅影響了秘魯鳀的分布及其與其它種類間的關系,同時也影響到了秘魯鳀漁業生物學的其它部分(包括種群結構、繁殖和早期生活史等),進而影響到其資源變動。
2.1 種群結構
根據秘魯鳀主要的產卵區域及商業性開發的地理位置(圖3)[19,32-34],可以將秘魯鳀分成三個主要的種群,分別是中北秘魯種群(north central Peru stock,NCP,4°S~15°S)、南秘魯北智利種群(southern Peru-northern Chile stock,SPNC,16°S~24°S)和中南智利種群(central-southern Chile stock,CSC,又叫做塔爾卡瓦諾種群Talcahuano stock,33°S~42°S)。CAHUIN等[33]分析了1963~2004年NCP和SPNC的上岸量、產卵群體生物量(spawning stock biomass,SSB)和補充量的時間序列發現,除了在El Ni?o現象發生的時間內,兩個種群的波動變化顯著相關。
El Ni?o現象的發生對秘魯鳀的種群結構有著重要影響。?IQUEN等[20]比較了1997~1998年El Ni?o現象發生前后NCP捕撈群體的體長組成發現,El Ni?o發生后捕撈群體體長組成較發生前偏小,未成熟的稚魚比例增多,性腺成熟度也較往年偏低。CANALES等[35]對北部智利采集的捕撈群體的體長組成進行分析,也發現了同樣的結果(圖4),同時結合方差分析發現,捕撈群體的平均體長與SST呈顯著的負相關關系(29.870-0.763SST,P=0.041,R2=0.243)。
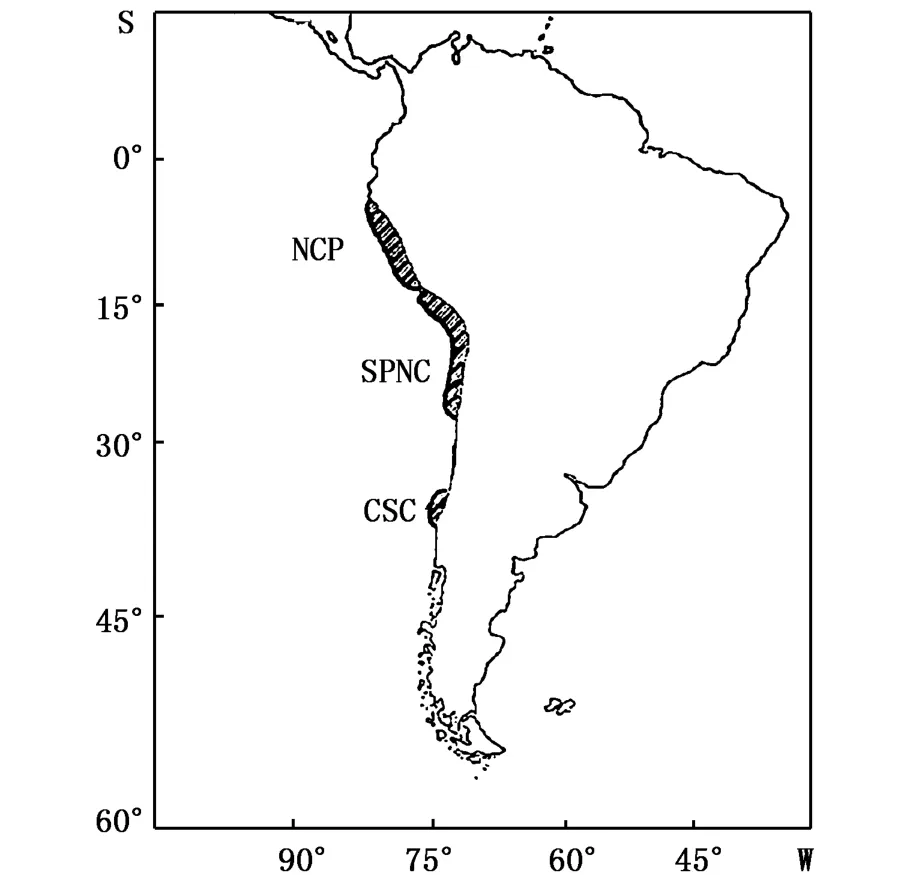
圖3 秘魯鳀的種群分布[19]Fig.3 Stock distribution of anchoveta(E.ringens)[19]

圖4 1990~2009年北部智利捕撈群體的平均體長和最大體長的年間變化[35]Fig.4 Themean and maximum body length of anchoveta(E.ringens)fished in northern Chile from 1990 to 2009[35]
2.2 繁殖
秘魯鳀在產卵時間上的繁殖策略會對環境進行權衡。在上升流強的夏季產卵,后代能夠得益于高的食物密度,但是強的埃克曼輸送會將暫時活動能力較弱的幼魚輸送至離岸食物條件不利的區域;同時在春夏季,幼魚捕食者的生物量也會增加。而冬季產卵雖能夠得益于近岸較弱的上升流(北部)或下降流(南部)減少遭遇捕食者的機會并靠近岸地,但是仔稚魚的餌料條件并不好[36]。CUBILLOS等[37]指出,秘魯鳀的產卵最高峰常常出現在季節轉換的時期,這個時期海表面風向經常發生南北轉變:北風能夠使海水向近岸輸送讓幼魚能夠停留在沿岸地帶,而不強的南風可以帶來溫和的上升流給予幼魚餌料[38]。這對南部的CSC種群最為有利:例如對1995年CSC的個體產卵行為的觀察就發現[39],8月份海表面的風向由北轉向南,而當年8月正是其主要的產卵時期。如表2所示,總體上,南部CSC的個體一般在4月開始產卵,6月達到頂峰,9月前后產卵結束,而北部的NCP和SPNC的個體在全年內都會產卵,但是從季節上看,這三個種群都在冬季有著產卵高峰期[33,38]。此外,MORI等[40]對1986~2008年北部的種群的性腺成熟度和產卵率的研究表明,NCP的產卵個體會根據環境和種群密度選擇一年產卵高峰次數:NCP種群的年齡組成存在著不同大小的個體,大的個體全年都會產卵,而小的個體(一般是初次達到性成熟)在到達性成熟時,若是海洋環境良好(適合產卵),在夏季可能會產生該種群的第二個產卵高峰期。CLARAMUNT等[38]認為,秘魯鳀的產卵有著自己的“能量儲存策略(energy storage strategy)”:在夏季上升流強,高生產力的情況下會為自己在冬季的產卵儲備能量。
在產卵地域的選擇上,研究發現,秘魯鳀的產卵并不是在穩定的水柱之內,而是在近岸海水湍流擾動較大的眾多水團之間和溫躍層附近,以使得幼體獲得更多的餌料[34,39,41]。CLARAMUNT等[41]使用廣義可加模型研究了調查海域的產卵數量(egg number)與產卵的空間位置(經度longitude和緯度latitude)、SST和葉綠素a濃度(chlorophyll-a,chl-a)之間的關系,最適模型表明〔egg number=s(latitude)+s(longitude)+s[log(chl-a))]〕chl-a和產卵的空間位置決定了產卵的數量,SST并不是影響其產卵數量的因素,這也進一步地表明其產卵地點的選擇是為了使幼體獲得更多的餌料;溫度對其產卵的影響主要還是在產卵時間和產卵頻率上。
但是也有研究表明,溫度影響到了秘魯鳀胚胎的尺寸[41-43]。LEAL等[42]發現,產卵前60~90 d環境的溫度對秘魯鳀卵母細胞大小有著重要的影響。有報道發現,在溫度較高的低緯度(20°S)發現的胚胎(平均體積0.2 mm3),其大小要比溫度較低的高緯度(36°S)小(平均體積0.31 mm3)[43]。大的胚胎孵化出的個體體形較大,生長也較快,能夠加大逃避捕食者和尋找食物的機率,因此孵化出大胚胎也是秘魯鳀親體對低溫不利幼體環境的一種響應[40,43]。

表2 秘魯鳀產卵概況[33,38,45]Tab.2 Review of the spawning of anchoveta(E.ringens)
2.3 早期生活史
秘魯鳀的胚胎發育、幼魚的成活及生長受到了產卵地點和時間、溫度及氣候變化等多種因素的影響。
SOTO-MENDOZA等[46]利用了基于個體的模型(individual-based model,IBM)模擬了8~10月中南部智利秘魯鳀胚胎和幼魚的傳送及成活情況,結果發現,平流輸送(advection,AD,指水流將秘魯鳀胚胎或幼魚輸送至對其不利的環境,并且一定會對其造成死亡的情況)造成的幼魚死亡率要比溫度變化導致的死亡率要高(圖5),利用廣義線性模型對這兩個因子進行分析發現[46],產卵及孵化的地點和時間對這兩個因子均有顯著的影響:產卵地南部的胚胎或幼魚更容易受到洪堡海流的影響向北部傳送,而模型的結果表明,從南部智利雷布-科拉爾(Lebu-Corral,38°S~40°S)沿岸最終輸送到北部35°S附近的胚胎或幼魚成活率非常低(分別為3.69%和0.01%);南部區域親魚的產卵若發生在早春9~10月,風向的轉變(由極向轉為赤道方向)使得研究區域開始存在強勁上升流,并伴隨著離岸埃克曼輸送,因此增加了平流輸送導致的死亡率。PARADA等[45]同樣利用了IBM模型模擬了中南部智利秘魯鳀的幼魚輸送,并且與已有的資料進行對比發現,在冬季產卵生長的幼魚其活動區域與該地區歷史上的幼魚發現區域相匹配,即使有上升流的存在,幼魚的離岸平流輸送也會被上升流在外部所產生的水舌所阻擋。
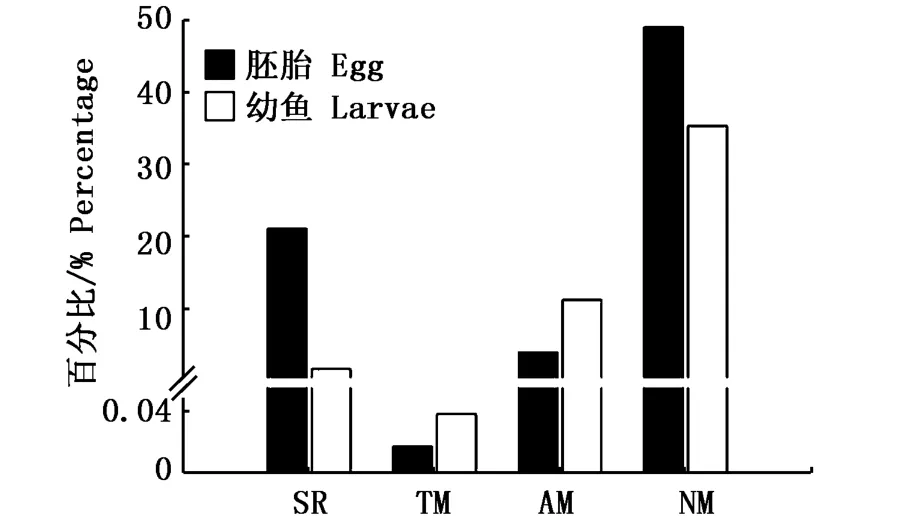
圖5 溫度和平流輸送對秘魯鳀胚胎及幼魚存活的影響[45]Fig.5 Influence of temperature and advection on the survival rate of anchoveta(E.ringens)eggs and larvae[45]
溫度對秘魯鳀胚胎或幼魚的成活影響較小,BROCHIER等[47]對北部秘魯種群的模擬結果也發現,除非產卵的密度在一定空間范圍內增加到某個限度,溫度不是制約胚胎或幼魚成活的限制因素。但也有研究發現,胚胎的孵化時間及幼魚帶卵黃(yolk-sac)期的持續時間都隨著溫度的升高而降低[7,43]。
另外,El Ni?o事件會對秘魯鳀胚胎和幼魚的成活具有明顯影響。ROJAS等[48]調查發現,在1997~1998年的El Ni?o事件發生時,秘魯鳀的胚胎和幼魚水平方向上朝南偏移,垂直方向上往深海移動,同時近岸的小型浮游動物增多,棲息地及餌料組成的變化導致了胚胎和幼魚較高的死亡率。
3 秘魯鳀資源變動與氣候變化的關系
秘魯鳀的資源變動和與氣候相關的海洋生態系統周期性變化(regime shift)有著密切的聯系。海洋生態系統的周期性變化指一定區域內的海洋生態系統狀況尤其是在營養級層面上的生物組成發生的低頻率周期性的高震蕩轉變[49]。ALHEIT等[19]通過觀察1950年以后的整個秘魯上升流生態系統的生物組成變化,發現海域內存在著冷暖兩個不同的時期,冷時期(1950~1970年和1985年以后)海域中秘魯鳀占主導地位,因此又叫做秘魯鳀時期(anchovy regime);暖時期(1970~1985年)海域中遠東擬沙丁魚(Saedinops sagax)占主導地位,因此又叫做沙丁魚時期(sardine regime)。而CHAVEZ等[5]觀察發現,太平洋在20世紀70年代中期轉變為El Ni?o事件多發的暖水時代,直至1990年以后才重新轉化為冷水時代,不同的時代大氣和海洋的要素都明顯不同,同樣以東南太平洋秘魯鳀和遠東擬沙丁魚的產量為標志,將太平洋分成秘魯鳀時期和沙丁魚時期兩個不同的階段。與ALHEIT等[19]研究結果不同的是,CHAVEZ等[5]定義的太平洋重新恢復到秘魯鳀時期的時間為20世紀90年代中期。此外,OLIVEROS-RAMOS等[50]通過計算1961~2009年秘魯鳀魚的補充量,根據補充量的變化定義了三個補充量時期(圖6):1961~1971年、1971~1991年和1991~2009年,其中第一和第三個時期之間差異不顯著(P=0.227 1),而中間一段時期與其前后兩個時期差異極顯著(P<0.01),他們認為,ALHEIT等[19]的結果未考慮到1985年前后秘魯鳀補充量的變化和捕撈努力量的增加。因此海洋生態系統周期性變化的時期劃分應與CHAVEZ等[5]提出的結果一致。關于整個秘魯鳀與海洋生態系統周期性變化的響應機制還不是很清楚,但是以上作者都指出[5,19,51],人類活動(對秘魯鳀以及海域其它資源的過度捕撈)對秘魯鳀資源量的變化及生態系統的擾動是不可忽視的。
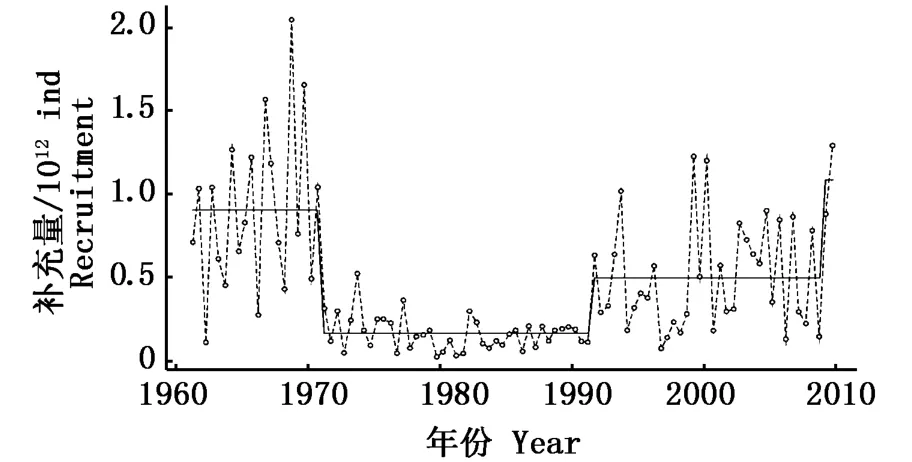
圖6 1960~2010年秘魯鳀的補充量[50]Fig.6 Recruitment of anchoveta(E.ringens)from 1996 to 2010[50]
更長的時間范圍上看,GUI?EZ等[51]利用北部智利的梅希約內斯灣(Mejillones Bay,23°S)的海底沉積物,分析了歷史上的秘魯鳀資源動態與環境的關系,有以下兩點發現:第一,在1500~1850年的小冰川時期(Little Ice Age)內,El Ni?o事件發生頻率的較高,海域內有著高溶解氧和低生產力的特征同時伴隨著長時間較低的秘魯鳀資源豐度;在這個時間段后,海水溫度降低,上升流活動變強及生產力的豐富,秘魯鳀的資源豐度也隨之變高;第二,秘魯鳀的資源變動與25~30年一個相位的PDO有著負相關的關系。
4 分析與展望
秘魯鳀是r選擇型種類,有著短生命周期(4年)、生長快速(孵化到成熟只需要一年)、高死亡率和對環境變化極其敏感的特點[31,52]。海洋環境變化對其資源變動的影響受到了研究者的關注,雖然在一定程度上揭示了其中的作用機制,但是目前僅僅利用這種關系來預測秘魯鳀的資源變動還是十分困難。準確地進行秘魯鳀資源量預報不僅有助于資源的合理利用和科學管理,同時也有助于節省漁船的成本,安排合適的捕撈努力量,制定合適的漁業政策,這也是適應負責任捕撈和漁業可持續發展的要求[53]。現有的研究發現,捕撈因素的加入十分必要。Yá?EZ等[54]分析了北部智利秘魯鳀CPUE與SST、氣候因子以及秘魯鳀和遠東擬沙丁魚的產量的關系,結果發現,CPUE只與捕撈當月算起7個月之前的智利安托法加斯塔港的水溫(SST of Antofagasta,SSTA)以及前幾個月內的各月秘魯鳀和遠東擬沙丁魚的產量有關,其中上一個月的秘魯鳀產量是最重要的一個因子,并基于這些因子利用神經網絡模型進行了 CPUE的預報。GUTIéRREZ-ESTRADA等[55]結合了自相關移動平均(Auto-Regressive Integrated Moving Average,ARIMA)和神經網絡的方法,只用所需預測月份前六個月內各月秘魯鳀的產量作為輸入層預測當月的產量。這些研究很好地考慮到了產量月間變化的時間自相關關系,但是只能應用在短的時間范圍之內。分析表明,秘魯鳀的資源變動不僅僅只是簡單地受年際ENSO事件的影響,同時多種因素也共同作用使得其資源發生變動。由于不了解秘魯鳀與整個生態系統周期性變化的響應機制,因此對于中長期年際的資源量的預測還是很困難。秘魯生態系統的冷暖時期交替之前,是否有具體的現象發生進而能預測秘魯鳀資源的走向?1970年秘魯鳀達到歷史上最高的產量之后幾年迅速下降(圖1),這剛好與交替時期重疊,那么交替時期的海洋環境變化和捕撈因素是如何綜合作用于種群的內部動力過程?這些問題都有待于今后進一步的研究。
本研究認為,今后在加強對長時間尺度冷暖世代周期性變化的研究同時,也應注重結合環境變化、捕撈因素及種群的內部動力過程這三者之間的關系,基于個體的海洋動力學模型,建立適合秘魯鳀的資源評估及預測的模型,為合理開發和管理該資源提供基礎。
[1]方 海,張 衡,劉 峰,等.氣候變化對世界主要漁業資源波動影響的研究進展[J].海洋漁業,2008,30(4):363-370.
FANG H,ZHANG H,LIU F,et al.A summary of research progress related with the fluctuation of the worldwide main fishery resources influenced byclimate changes[J].Marine Fisheries,2008,30(4):363-370.
[2]XU Y,CHAIF,ROSE K A,et al.Environmental influences on the interannual variation and spatial distribution of Peruvian anchovy(Engraulis ringens)population dynamics from 1991 to 2007:A threedimensional modeling study[J].Ecological Modelling,2013,264(16):64-82.
[3]FRéON P,BOUCHON M,MULLON C,et al.Interdecadal variability of anchoveta abundance and overcapacity of the fishery in Peru[J].Progress in Oceanography,2008,79(2):401-412.
[4]Food and Agriculture Organization of the United Nations.Food and Agriculture Organization of the United Nations′fisheries database Global capture production from 1950 to 2013[DB/OL].http://www.fao.org/fishery/statistics/global-captureproduction/query/zh.
[5]CHAVEZ FP,RYAN J,LLUCH-COTA SE,etal.From anchovies to sardines and back:Multidecadal change in the Pacific Ocean[J].Science,2003,299(5604):217-221.
[6]TARIFE?O E,CARMONA M,LLANOS-RIVERA A,et al.Temperature effects on the anchoveta Engraulis ringens egg development:Do latitudinal differences occur?[J].Environmental Biology of Fishes,2008,81(4):387-395.
[7]CHAVEZ F P,MESSIéM.A comparison of eastern boundary upwelling ecosystems[J].Progress in Oceanography,2009,83(1):80-96.
[8]吳日升,李 立.南海上升流研究概述[J].臺灣海峽,2003,22(2):269-277.WU R S,LIL.Summarization of study on upwelling system in the South China Sea[J].Journal of Oceanography in Taiwan Strait,2003,22(2):269-277.
[9]HALPERN D.Offshore Ekman transport and Ekman pumping off Peru during the 1997-1998 El Ni?o[J].Geophysical Research Letters,2002,29(5):1075.
[10]MONTECINO V,LANGE C B.The Humboldt Current System:Ecosystem components and processes,fisheries,and sediment studies[J].Progress in Oceanography,2009,83(1):65-79.
[11]CHAIGNEAU A,GIZOLME A,GRADOSC.Mesoscale eddies off Peru in altimeter records:Identification algorithms and eddy spatio-temporal patterns[J].Progress in Oceanography,2008,79(2):106- 119.
[12]HORMáZABAL S,SHAFFER G,LETH O.Coastal transition zone off Chile[J].Journal of Geophysical Research,2004,109(C1):C01021.
[13]MONTECINO V,QUIROZ D.Specific primary production and phytoplankton cell size structure in an upwelling area off the coast of Chile(30°S)[J].Aquatic Sciences,2000,62(4):364-380.
[14]陳新軍.漁業資源與漁場學[M].北京:海洋出版社.2004:153.
CHEN X J.Fisheries biology and oceanography[M].Beijing:China Ocean Press,2004:153.
[15]MESSIéM,LEDESMA J,KOLBER D D,et al.Potential new production estimates in four eastern boundary upwelling ecosystems[J].Progress in Oceanography,2009,83(1):151-158.
[16]BAKUN A,WEEKS S J.The marine ecosystem off Peru:What are the secrets of its fishery productivity and what might its future hold?[J].Progress in Oceanography,2008,79(2):290-299.
[17]SWARTZMAN G,BERTRAND A,Gutiérrez M,et al.The relationship of anchovy and sardine to water masses in the Peruvian Humboldt Current System from 1983 to 2005[J].Progress in Oceanography,2008,79(2):228-237.
[18]PHILANDER S G.A review of tropical oceanatmosphere interactions[J].Tellus A,1999,51(1):71-90.
[19]ALHEIT J,NIQUEN M.Regime shifts in the Humboldt Current ecosystem[J].Progress in Oceanography,2004,60(2):201-222.
[20]?IQUEN M,BOUCHON M.Impact of El Ni?o events on pelagic fisheries in Peruvian waters[J].Deep Sea Research PartⅡ:Topical Studies in Oceanography,2004,51(6):563-574.
[21]SANDWEISSD H,MAASCH K A,CHAIF,et al.Geoarchaeological evidence for multidecadal natural climatic variability and ancient Peruvian fisheries[J].Quaternary Research,2004,61(3):330-334.
[22]GIBSON R N,ATKINSON R JA.Oxygen minimum zone benthos:Adaptation and community response to hypoxia[J].Oceanography and Marine Biology:An Annual Review,2003,20(41):1-45.
[23]BERTRAND A,CHAIGNEAU A,PERALTILLA S,et al.Oxygen:A fundamental property regulating pelagic ecosystem structure in the coastal southeastern tropical Pacific[J].PLoSOne,2011,6(12):e29558.
[24]BERTRAND A,BALLóN M,CHAIGNEAU A.Acoustic observation of living organisms reveals the upper limit of the oxygen minimum zone[J].PLoS One,2010,5(4):e10330.
[25]ESPINOZA P,BERTRAND A.Revisiting Peruvian anchovy(Engraulis ringens)trophodynamics provides a new vision of the Humboldt Current system[J].Progress in Oceanography,2008,79(2):215-227.
[26]BALLóN M,BERTRAND A,LEBOURGESDHAUSSY A,etal.Is there enough zooplankton to feed forage fish populations off Peru?An acoustic(positive)answer[J].Progress in Oceanography,2011,91(4):360-381.
[27]AYóN P,SWARTZMAN G,BERTRAND A,et al.Zooplankton and forage fish species off Peru:Largescale bottom-up forcing and local-scale depletion[J].Progress in Oceanography,2008,79(2):208-214.
[28]CHAVEZ F P,BERTRAND A,GUEVARA-CARRASCO R,et al.The northern Humboldt Current System:Brief history,present status and a view towards the future[J].Progress in Oceanography,2008,79(2):95-105.
[29]TAM J,TAYLOR M H,BLASKOVIC V,et al.Trophic modeling of the Northern Humboldt current ecosystem,PartⅠ:Comparing trophic linkages under La Ni?a and El Ni?o conditions[J].Progress in Oceanography,2008,79(2):352-365.
[30]BERTRAND S,DEWITTE B,TAM J,et al.Impacts of Kelvin wave forcing in the Peru Humboldt Current system:Scenarios of spatial reorganizations from physics to fishers[J].Progress in Oceanography,2008,79(2):278-289.
[31]BERTRAND A,GERLOTTO F,BERTRAND S,et al.Schooling behaviour and environmental forcing in relation to anchoveta distribution:An analysis across multiple spatial scales[J].Progress in Oceanography,2008,79(2):264-277.
[32]BARANGE M,COETZEE J,TAKASUKA A,et al.Habitat expansion and contraction in anchovy and sardine populations[J].Progress in Oceanography,2009,83(1):251-260.
[33]CAHUIN S M,CUBILLOS L A,ESCRIBANO R.Synchronous patterns of fluctuations in two stocks of anchovyEngraulis ringensJenyns,1842 in the Humboldt Current System[J].Journal of Applied Ichthyology,2015,31(1):45-50.
[34]CASTRO L R,CLARAMUNT G,KRAUTZM C,etal.Egg trait variation in anchovetaEngraulis ringens:A maternal response to changing environmental conditions in contrasting spawning habitats[J].Marine Ecology Progress Series,2009(5),381:237-248.
[35]CANALES TM,LAW R,WIFF R,et al.Changes in the size-structure of amultispecies pelagic fishery off Northern Chile[J].Fisheries Research,2015(161):261-268.
[36]PARRISH R H,NELSON C S,BAKUN A.Transport mechanisms and reproductive success of fishes in the California Current[J].Biological Oceanography,1981,1(2):175-203.
[37]CUBILLOS L,ARCOS D,BUCAREY D,et al.Seasonal growth of small pelagicfish off Talcahuano,Chile(37°S,73°W):A consequence of their reproductive strategy to seasonal upwelling?[J].Aquatic Living Resources,2001,2(14):115-124.
[38]CLARAMUNT G,CUBILLOS L A,CASTRO L,etal.Variation in the spawning periods ofEngraulis ringensandStrangomera bentinckioff the coasts of Chile:A quantitative analysis[J].Fisheries Research,2014,160(12):96-102.
[39]LEONARDO R C,CUSTAVO R.S,EDUARDO,H H.Environmental influences on winter spawning of the anchovetaEngraulis ringens,off Talcahuanv,central Chile[J].Marine Ecology Progress,2000,197(1):247-258.
[40]MORI J,BUITRóN B,PEREA A,et al.Interannual variability of the reproductive strategy of the Peruvian anchovy off northern-central Peru[J].Ciencias Marinas,2011(37):513-525.
[41]CLARAMUNTG,CASTRO LR,CUBILLOSL A,et al.Inter-annual reproductive trait variation and spawning habitat preferences ofEngraulis ringensoff northern Chile[J].Revista De Biología Marina Y Oceanografía,2012,47(2):227-243.
[42]LEAL E M,CASTRO L R,CLARAMUNT G.Variability in oocyte size and batch fecundity in anchoveta(Engraulis ringens,Jenyns 1842)from two spawning areas off the Chilean coast[J].Scientia Marina,2009,73(1):59-66.
[43]LLANOS-RIVERA A,CASTRO L R.Latitudinal and seasonal egg-size variation of the anchoveta(Engraulis ringens)off the Chilean coast[J].Fishery Bulletin-National Oceanic and Atmospheric Administration,2004,102(1):207-212.
[44]LLANOS-RIVERA A,CASTRO L R.Inter-population differences in temperature effects onEngraulis ringensyolk-sac larvae[J].Marine Ecology Progress,2006,312(8):245-253.
[45]PARADA C,COLASF,SOTO-MENDOZA S,etal.Effects of seasonal variability in across-and alongshore transport of anchoveta(Engraulisringens)larvae on model-based pre-recruitment indices off central Chile[J].Progress in Oceanography,2012,92(5):192-205.
[46]SOTO-MENDOZA S,PARADA C,CASTRO L,et al.Modeling transport and survival of anchoveta eggs and yolk-sac larvae in the coastal zone off centralsouthern Chile:Assessing spatial and temporal spawning parameters[J].Progress in Oceanography,2012,92(1):178-191.
[47]BROCHIER T,LETT C,TAM J,et al.An individual-based model study of anchovy early life history in the northern Humboldt Current system[J].Progress in Oceanography,2008,79(2):313-325.
[48]ROJAS P M,LANDEAETA M F,ULLOA R.Eggs and larvae of anchoveta Engraulis ringens off northern Chile during the 1997-1998 El Ni?o event[J].Revista De Biología Marina Y Oceanografía,2011,46(3):405-419.
[49]COLLIE J S,RICHARDSON K,STEELE J H.Regime shifts:Can ecological theory illuminate the mechanisms?[J].Progress in Oceanography,2004,60(2):281-302.
[50]OLIVEROS-RAMOS R,PE?A C.Modeling and analysis of the recruitment of Peruvian anchovy(Engraulis ringens)between 1961 and 2009[J].Ciencias Marinas,2011,37(4B):659-674.
[51]GUI?EZ M,VALDS J,SIFEDDINE A,et al.Anchovy population and ocean-climatic fluctuations in the Humboldt Current System during the last 700 years and their implications[J].Palaeogeography Palaeoclimatology Palaeoecology,2014(415):210-224.
[52]CUBILLOS L A,BUCAREY D A,CANALES M.Monthly abundance estimation for common sardineStrangomera bentinckiand anchovyEngraulis ringensin the central-southern area off Chile(34-40°S)[J].Fisheries Research,2002,57(2):117-130.
[53]陳新軍,高 峰,官文江,等.漁情預報技術及模型研究進展[J].水產學報,2013,37(8):1270-1280.
CHEN X J,GAO F,GUANW J,et al.Review of fishery forecasting technology and its models[J].Journal of Fisheries of China,2013,37(8):1270-1280.
[54]Yá?EZE,PLAZA F,GUTIéRREZ-ESTRADA JC,et al.Anchovy(Engraulis ringens)and sardine(Sardinops sagax)abundance forecast off northern Chile:A multivariate ecosystemic neural network approach[J].Progress in Oceanography,2010,87(1):242-250.
[55]GUTIéRREZ-ESTRADA JC,SILVA C,Yá?EZ E,etal.Monthly catch forecasting of anchovyEngraulis ringensin the north area of Chile:Non-linear univariate approach[J].Fisheries Research,2007,86(2):188-200.
Review on relationship between oceanic environment factors and population dynamics of anchoveta Engraulis ringens
CHEN Peng1,4,WANG Jin-tao1,4,CHEN Xin-jun1,2,3,4
(1.College of Marine Sciences of Shanghai Ocean University,Shanghai201306,China;2.National Distant-waterFisheries Engineering Research Center,Shanghai Ocean University,Shanghai 201306,China;3.Key Laboratory ofSustainable Exploitation of Oceanic Fisheries Resources,Ministry of Education,Shanghai Ocean University,Shanghai 201306,China;4.Collaborative Innovation Center for Distant-water Fisheries,Shanghai 201306,China)
Anchoveta(Engraulis ringens)is a small pelagic fish.As an important commercial fish,fishing of this species has been the world’s largest single fish fishery.However,the catch of anchoveta shows big annual differences.The mechanisms of this yield and Peru upwelling where the fish habitat can be summarized by these five aspects:(1)low latitude geographical position of Peru upwelling;(2)suitable water temperature structure;(3)low dissolved oxygen in ocean;(4)high energy transfer efficiency in food web;(5)complex changes of oceanic environment factors.Oceanic environment has significant effects on the anchoveta fishery biology in many ways.In addition,the regime shift of ocean ecosystem relating to the climate can also influence the anchoveta population dynamics.We suggest that the future studies should highlight the regime shifts with the long temporal scale.At the same time,we should focus on integrating the environment changes,fishing factors,population dynamics and their internal connecting links so that we can build the valid stock assessment or forecast model to give theoretical basis for the sustainable utilization and management of anchoveta.
anchoveta(Engraulis ringens);oceanic environment;population dynamics;Peru upwelling;climate change
S 931
A
1004-2490(2016)02-0206-11
2015-05-25
國家自然科學基金(NSFC41476129;NSFC41276156);國家863計劃(2012AA092303);國家科技支撐計劃(2013BAD13B01)
陳 芃(1992-),男,碩士研究生。E-mail:pengchen@yeah.net
陳新軍,教授。Tel:021-61900306,E-mail:xjchen@shou.edu.cn
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
資源節約與環保(2022年8期)2022-09-20 02:25:22
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
藝術品鑒(2020年7期)2020-09-11 08:04:44
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
當代貴州(2018年28期)2018-09-19 06:39:04
資源再生(2017年3期)2017-06-01 12:20:59
決策(2015年9期)2015-09-10 07:22:44
