基于小鼠大腦MRM圖像灰度分布的個體結構網絡構建的研究
2016-03-02 10:07:33付振榮林嵐張柏雯吳水才
中國醫療設備 2016年12期
付振榮,林嵐,張柏雯,吳水才
北京工業大學 生命科學與生物工程學院,北京 100124
基于小鼠大腦MRM圖像灰度分布的個體結構網絡構建的研究
付振榮,林嵐,張柏雯,吳水才
北京工業大學 生命科學與生物工程學院,北京 100124
結構磁共振成像腦網絡分析技術結合了sMRI成像、醫學圖像處理、復雜網絡及圖論分析等技術方法,通過對大腦不同感興趣區之間的特征關系進行分析,能夠更好地分析腦組織結構之間的關系及與疾病的關聯性等。目前,基于人腦sMRI的大腦灰質結構網絡已成為腦網絡研究的一個熱點,但是基于小鼠MRM的大腦灰質網絡分析還相對較少。本文首先通過構建基于KL散度的小鼠MRM個體灰質腦網絡,并在此基礎上進行網絡拓撲屬性的分析,該方法可以應用于腦老化和因疾病帶來的小鼠腦網絡變化的研究。
小鼠大腦;個體腦網絡;KL散度
近年來,隨著神經影像數據采集設備和技術的快速進步,研究者們逐漸將復雜網絡的相關理論應用到大腦研究之中,腦網絡分析也隨之成為神經影像學領域的研究熱點[1-3]。基于彌散張量成像(Diffusion Tensor Imaging,DTI)[4]、功能性磁共振成像(Functional Magnetic Resonance Imaging,fMRI)[5]、結構磁共振成像(Structure Magnetic Resource Imaging,sMRI)[6]、腦電圖(Electroencephalo-Graph,EEG)[7]和腦磁圖(Magnetoencephalogram,MEG)[8]等腦成像技術被用于構建復雜腦網絡。通過對復雜網絡拓撲屬性的分析,可以在一定程度上幫助研究者了解大腦結構與功能之間的相互關系[9],揭示大腦傳遞信息與處理信息的運行機理,還能夠用于大腦年齡預測[10-11]以及疾病的診斷和預測[12-14]等研究。
由于sMRI易獲取、包含腦區間的結構連接信息等優勢,基于它的結構網絡也越來越受到研究者們的重視。腦網絡構建主要包含兩個部分,即節點的定義和節點間的連接強度(邊)的定義。Gene等[15]依據大腦中不同腦區間存在協同變化的原理,采用一種改良的主成分分析方法(Scaled Subprof le Model,SSM),得到了反映載脂蛋白ApoE ε4等位基因攜帶者的結構特征網絡。此方法可以用來衡量不同個體對該結構特征網絡的表示程度,但此方法無法通過圖論進行復雜網絡分析。He等[16]通過將對象分組大腦分區,計算不同組對象各個腦區間的相關性得到全腦的相關性矩陣,并進一步通過圖論分析發現該網絡具有小世界特征和無標度性。該方法雖能夠采用不同的結構特征構建網絡,但只能基于一組數據構建一個平均腦網絡。由于該方法構建的是一組的平均腦網絡,對數據量要求較大。同時,平均網絡是基于一組對象的平均特征,無法準確反應個體對象間的差異。針對平均網絡的不足之處,研究人員提出了構建個體網絡的新方法。Tijms等[17]將大腦定義為6982個立方體的腦區節點,各個節點內部包含的27個體素可以組成特征向量,通過計算這些特征向量間的相關性可以得到腦區連接的強度。該方法可以構建個體對象的結構網絡,但立方體大小和其中的旋轉操作存在一定的任意性。Kong等[18]提出了基于KL散度(Kullback-Leibler_divergence)得到KL散度相似性度量(Kullback-Leibler_divergence Similarity,KLS)構建大腦個體灰質網絡的方法。KL散度能夠間接反映不同感興趣區之間的相似度,通過計算不同腦區間特征的概率密度分布得到不同腦區之間的KL散度。
動物模型在醫學領域的研究中有著不可替代的作用。與人類相比,動物模型具有可操作性強、生命周期短、以及能夠批量生產等優勢,能夠幫助科研人員加快研究進度以及更全面的認識疾病等研究內容的本質。隨著高分辨率影像技術的快速發展,能夠采集小動物大腦圖像的磁共振顯微成像技術(Magnetic Resource Microscopy,MRM)[19]為利用動物模型進行神經影像的研究提供了技術支持。MRM技術不僅能夠采集高分辨率的小鼠活體三維圖像,而且避免了組織切片中的空間變形。基于MRM 的小鼠腦圖像分析的發展吸引了不少研究人員的關注[20]。
目前,基于人腦sMRI的灰質結構網絡的研究正處于高速發展階段[21-22],但基于動物模型構建結構腦網絡的研究還相對較少。人腦和鼠腦無論是在形態還是體積上都存在非常大的差異,例如,鼠腦的體積一般僅為人腦的三千分之一,并且鼠腦的表面較光滑,沒有復雜的腦溝與腦回。本文將KL散度的腦網絡構建進一步從人腦延伸到了小鼠腦MRM之中。本研究主要基于小鼠的T2 MRM影像展開。T2 MRM影像反映的是組織的橫向磁化強度Mxy衰減的快慢。不同腦區間的圖像灰度分布的不同,反映了不同腦區間橫向弛豫時間的差異。而這種差異在一定程度上可以反映不同腦區間組織特征的差異性(如神經元細胞的種類、神經元細胞的密度等)。本文基于小鼠不同腦區間的KLS建立小鼠大腦的個體灰質網絡,并通過進一步分析網絡的拓撲屬性,驗證這種方法在小鼠MRM個體腦網絡構建中的適用性及延展性。
1 數據采集
本次研究采用的數據來自于新加坡國立大學(Computational Functional Anatomy Lab)的小鼠影像數據庫(http://www.bioeng.nus.edu.sg/cfa/mouse_atlas.html)。該數據庫包含5只雄性C57BL/6小鼠T2 MRM 影像。圖像采用德國布魯克7-T/20-cm ClinScan MRI進行采集,其中掃描序列為3DT2加權快速自旋回波(重復時間/回波時間=2000 ms/46 ms,視野范圍=9 mm×13 mm×25 mm,體素尺度= 100×98×98 micron),采集時間為1小時23分[23]。由于7-T磁場會給圖像帶來較強的不均勻性[24-25],數據庫中的MRM圖像都采用N3(Nonparametric Non-uniform Intensity Normalization)[26]進行非均勻場校正,并且灰度被標準化到0~255。數據庫中小鼠圖像均被手工分割為39個感興趣區。腦區劃分流程為,首先將圖像剛體配準到Franklin and Paxinos小鼠腦圖譜空間中[27-28],再將MRM圖像進行手工分割。圖譜劃分原則為:首先將容易分辨的腦區劃分出來,然后再將不易識別、輪廓不清晰的腦區嚴格按照組織切片圖譜來進行劃分,在很大程度上保持了腦區劃分的準確性。由于在腦網絡分析中,只需要大腦灰質和神經核團。本研究利用已分割的小鼠大腦個體腦區圖譜去除14個相關腦區,其分別屬于腦白質、腦脊液、小腦和腦干等部分。然后,剩余的25個腦區被用于腦網絡構建。25個灰質腦區詳細定義見表1。
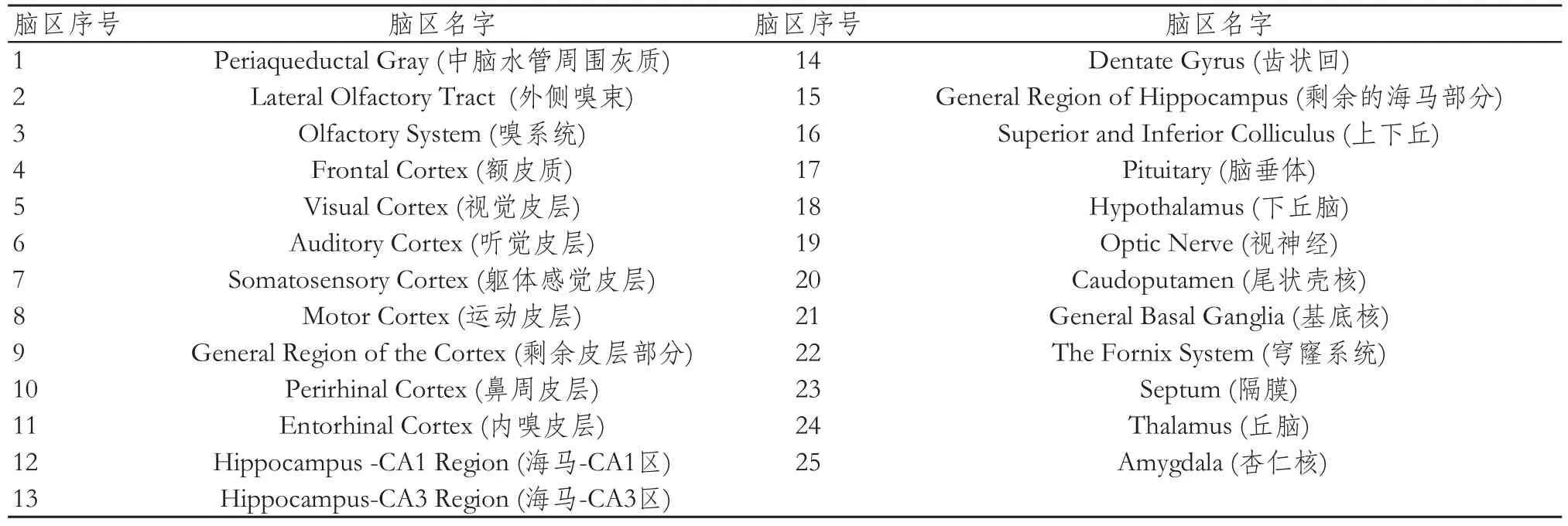
表1 小鼠腦區對照表
2 方法及結果
本研究構建了兩種類型的腦網絡:一種為基于小鼠腦T2 MRM圖像構建的灰質個體網絡;另一種為將小鼠腦T2 MRM進行高斯平滑處理 (FWHM為4個體素)后所構建的網絡。兩種方法網絡構建的流程基本一致,詳見圖1。
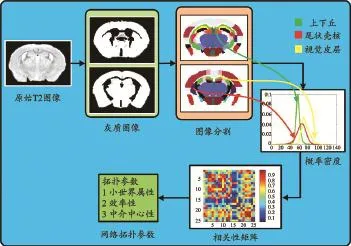
圖1 基于小鼠腦T2 MRM圖像的個體灰質網絡構建流程圖
該流程主要分為以下3個步驟:
(1)基于已有的小鼠腦個體感興趣區分區對小鼠腦MRM圖像進行分割。
(2)計算各個感興趣區的灰度概率密度分布,得到反映兩兩感興趣區相關關系的KLS相關性矩陣。
(3)基于KLS相關性矩陣構建網絡,并進行復雜網絡計算與分析。
2.1 網絡節點及邊
在基于腦區水平的大尺度的腦網絡構建中,定義網絡節點和網絡連接(邊)是最關鍵的兩步。本文中25個感興趣區被定義為大腦結構網絡的節點。MRM影像中的灰度差異反映了小鼠不同腦組織間的T2值的差異。這里我們通過基于KL散度的相似性度量KLS來計算兩個腦區之間的圖像灰度概率密度分布的相關性。KL散度為一種通用的可以計算兩個概率密度分布相似度的度量方法。其中,各個腦區的灰度密度分布通過核密度估計(Kernel Density Estimation,KDE)來進行計算(http://cn.mathworks.com/matlabcentral/fileexchange/14034-kernel-density-estimator?Requested Domain = www.mathworks.com)。由于數據庫中的小鼠圖像的灰度已經被標準化到0~255之間,這里將KDE的采樣點個數定為256個。在得到每個腦區的灰度概率密度分布之后,計算各個腦區間的KL散度,然后再通過KL散度來計算KLS,即得到相關性矩陣。KLS的范圍為0到1,1表示兩個腦區具有完全相同的灰度概率密度分布,而0表示兩個腦區的灰度概率密度分布不存在任何交集。KL距離的計算公式為:
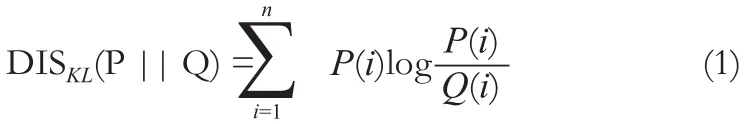
其中n為灰度分布中樣本點的個數,P和Q代表不同腦區的灰度分布。由于DISKL(P||Q)和DISKL(Q||P)不是等值的,我們用對稱的方法來消除這種不等值帶來的誤差,即將DISKL(P||Q)與DISKL(Q||P)相加得到一個新的KL距離。
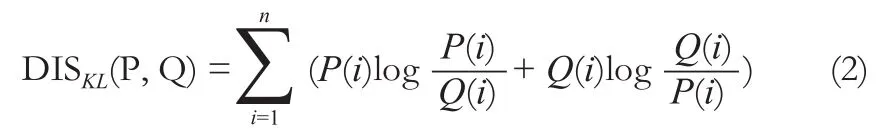
最后,KLS的計算公式為:
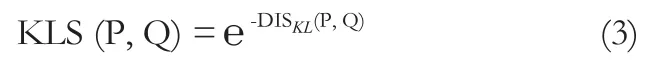
2.2 網絡分析
基 于MATLAB的GRETNA 工 具 包(http://www.nitrc.org/projects/gretna/)[29]被 用 來 進 行復 雜 網 絡 分 析。GRENTA是 北 京 師 范 大 學 開發的具有圖形化界面的能夠進行網絡分析的工具箱。本文中的主要網絡指標[30]包括:聚類系數(clustering coeff cient,Cp)、標準化聚類系數(標準化Cp)、最短路徑長度(Characteristic Pathlength,Lp)、標準化最短路徑長度(標準化Lp)、全局效率(global eff ciency,Eglob)、標準化全局效率(標準化Eglob)、局部效率(local eff ciency,Eloc)和標準化局部效率(標準化Eloc)和網絡中心節點。隨機網絡的構建是通過拓撲重新布線算法(Topological rewiring Algorithm)[31]產生100個與真實網絡具有一致的節點數、邊數和度分布的隨機網絡。此外,筆者還計算了以上各個參數在所定義的稀疏度范圍內的積分,即曲線下面積(Area Under Curve, AUC):聚類系數AUC(aCp)、最短路徑長度AUC(aLp)、全局效率AUC(aEglob)、局部效率AUC(aEloc)及其相應的標準化參數。這些參數值使得我們可以獨立于具體稀疏度閾值來分析網絡拓撲屬性。每個加權連接矩陣中的相關系數都需要取特定的閾值進行二值化,二值化網絡保留了相關系數中相對較大的連接。目前尚無確定的方法來選擇一個唯一的稀疏度,因此我們將稀疏度范圍定為0.15~0.4,每隔1%選取一個稀疏度。
2.3 結果分析
圖2為小鼠腦MRM圖像和平滑后的MRM圖像,通過KLS計算生成的平均相關性矩陣,其中二值化網絡的網絡稀疏度為20%。圖2中的左圖為平均加權相關性矩陣,相對未平滑的圖像,經過平滑處理后的圖像各腦區間具有更強的相關性。經過平滑運算,每個腦區和它相臨近的腦區間的相關性都會得到提升,具體表現為腦區相關性的整體提高。由于平滑處理對各個腦區邊緣的灰度分布影響更大,因此它對于各個腦區的影響是不一致的。一般腦區體積越小,與周邊其它腦區的灰度分布差異越大,平滑操作的影響也就越大。圖2中的右圖為二值化矩陣,平滑前后,網絡的整體拓撲結構有了較大的變化。
小世界網絡是介于隨機網絡和規則網絡之間[32-34]的一種網絡。小世界網絡具有較短的最短路徑長度和較大的聚類系數這兩個重要的特點。聚類系數越大的網絡,局部效率就會越高;最短路徑長度越長,全局效率就會越低。規則網絡的聚類系數較高,但是最短路徑長度較長;隨機網絡的聚類系數較低,但最短路徑長度較短。小世界網絡作為這兩種網絡的中間產物,具有比規則網絡高的聚類系數,以及比隨機網絡低的最短路徑長度。腦網絡如果是小世界網絡,與隨機網絡相比,應具有更高的聚類系數值與局部效率(標準化 Cp>1,標準化 Eloc >1),同時具有相近的最短路徑長度和全局效率(標準化 Lp~1 ,標準化 Eglob~1)。圖3為兩種不同處理流程(數據庫中MRM圖像與平滑后圖像)處理后,標準化的腦網絡參數在不同稀疏度下的分布圖。對于未平滑處理的圖像生成的網絡,標準化Cp的值隨稀疏度的增大呈下降趨勢,標準化Cp的最大值為5.78,最小值為1.40,總體均值為2.27,標準差為0.78,小鼠結構腦網絡與隨機網絡相比具有較高聚類系數;標準化 Eloc的值隨稀疏度的增大呈下降趨勢,標準化 Eloc 的最大值為5.75,最小值為1.12,總體均值為1.90,標準差為0.79,小鼠結構腦網絡與隨機網絡相比具有較高的局部效率;標準化Lp的值隨稀疏度的增大呈緩慢下降趨勢,趨向于1,標準化Lp的最大值為2.47,最小值為1.10,總體均值為1.29,標準差為0.24,小鼠結構腦網絡與隨機網絡相比最短路徑長度相近;標準化Eg的值隨稀疏度的增大呈緩慢上升趨勢,趨向于1,標準化Eg的最大值為0.92,最小值為0.40,總體均值為0.79,標準差為0.11,小鼠結構腦網絡與隨機網絡相比全局效率相近。對于平滑后圖像生成的網絡,標準化Cp的值隨稀疏度的增大呈下降趨勢,標準化Cp的最大值為4.41,最小值為1.48,總體均值為2.52,標準差為0.79;標準化 Eloc的值隨稀疏度的增大呈下降趨勢,標準化 Eloc的最大值為4.31,最小值為1.22,總體均值為2.06,標準差為0.83;標準化Lp的值隨稀疏度的增大呈緩慢下降趨勢,標準化Lp的最大值為1.92,最小值為1.07,總體均值為1.29,標準差為0.26;標準化Eg的值隨稀疏度的增大呈緩慢上升趨勢,標準化Eg的最大值為0.94,最小值為0.52,總體平均值為0.80,標準差為0.13。該網絡與隨機網絡相比,具有較高聚類系數、局部效率和相近的最短路徑長度與全局效率。兩種網絡均滿足小世界屬性。
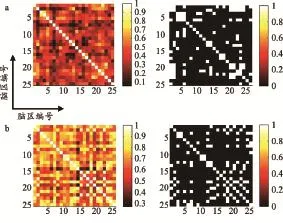
圖 2 平均相關性矩陣示意圖
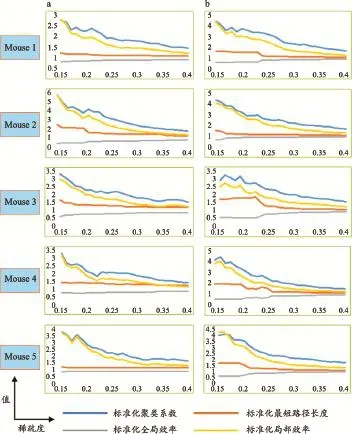
圖3標準化腦網絡參數分布圖
圖4為基于AUC值的各個樣本的全局網絡參數示意圖。五只不同小鼠個體網絡的聚類系數AUC的均值為0.13,標準差為0.02;最短路徑長度AUC的均值為0.72,標準差為0.12;標準化聚類系數AUC的均值為0.56,標準差為0.12;標準化最短路徑長度AUC的均值為0.32,標準差為0.05,全局效率AUC的均值為0.09,標準差為0.02、局部效率AUC的均值為0.15,標準差為0.02,標準化全局效率AUC的均值為0.20,標準差為0.02;標準化局部效率AUC的均值為0.47,標準差為0.09。對于平滑后圖像生成的網絡參數,五個不同小鼠個體網絡的聚類系數AUC的均值為0.17,標準差為0.01;最短路徑長度AUC的均值為0.62,標準差為0.06;標準化聚類系數AUC的均值為0.63,標準差為0.02;標準化最短路徑長度AUC的均值為0.32,標準差為0.02、全局效率AUC的均值為0.11,標準差為0.01;局部效率AUC的均值為0.19,標準差為0.004、標準化全局效率AUC的均值為0.20,標準差為0.01;標準化局部效率AUC的均值為0.51,標準差為0.04。不同小鼠的網絡參數的AUC值在平滑后個體差異性減少。
網絡中的hub節點是網絡通訊中重要的中心節點,定義hub節點的方法有很多,比如度、中介性指標等。我們前期的研究發現這些定義指標間具有很強的相關性[35],因此這里采用中介性指標來定義Hub。首先計算中介性指標在25個腦區的平均值,各個腦區中介性指標的大小反映了該腦區節點在網絡中的重要性。然后根據以下公式計算各個小鼠個體腦區節點重要性的相似度和特異性。
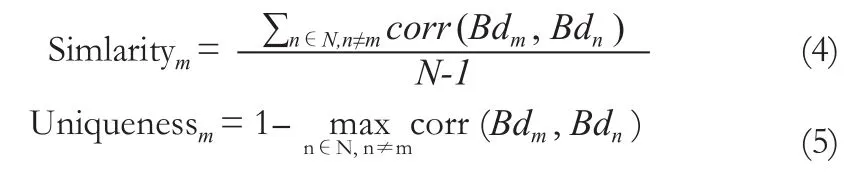
m和n為小鼠的編號,N為小鼠個體的總數,Bdm與Bdn為整體網絡稀疏度范圍內的平均節點中介性指標。圖5為5只小鼠的個體網絡腦區節點中介性指標的相似度與特異性的平均值與標準差。不同個體的腦區節點中介性指標具有較大的特異性,相似度較低。平滑處理可以提高相似度,降低它們之間的特異性。
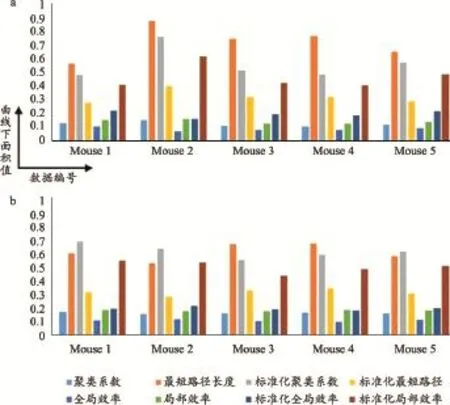
圖4 網絡參數的AUC值
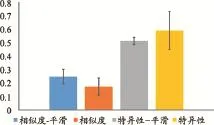
圖5 個體網絡腦區節點中介性指標的相似度與特異性
3 討論
隨著sMRI成像技術的快速發展,復雜網絡及圖論分析的方法被用來分析大腦網絡的內部關聯性。由于sMRI的易獲取等優點,利用sMRI構建大腦結構網絡得到了專家學者們的高度重視,各種新的方法也不斷涌現。本研究將基于KL散度的個體腦網絡成功延伸到了小鼠腦MRM個體結構網絡的研究中,構建的個體灰質網絡滿足小世界屬性,這應該是第一個基于結構特征的小鼠腦網絡。與隨機網絡相比,構建的灰質網絡具有較高的聚類系數、局部效率,以及與隨機網絡相近的最短路徑長度與全局效率。當前,對于灰質結構網絡的具體生理意義還不是十分明確,筆者認為它有可能與大腦發育過程中各個腦區的協同工作有關。即使一個簡單的功能,大腦都需要調動多個腦區的協同工作來完成。當這些腦區協同工作時,非常可能會促進這些腦區內部的細胞協同發育,而在腦圖像中這些腦區可能會出現較相近的神經細胞分布。
空間平滑操作是神經影像處理中的一種常用操作,它可以用來提高圖像的信噪比和減小配準誤差帶來的腦區劃分差異。由于小鼠大腦體積較小,每個體素的體積也非常小,圖像信噪比較差。同時,由于小鼠大腦的個體差異性相對人腦也較少,所以對配準算法提出了較高要求[36]。因此空間平滑操作對于小鼠圖像是存在一定必要性的。結果顯示,平滑圖像生成的腦網絡仍然具有小世界特性,同時多種網絡參數的AUC結果顯示,個體對象的網絡參數的個體特異性減小。腦區重要性的研究也顯示平滑處理可以提高不同個體腦區節點重要性的相似度和降低其特異性。由于平滑操作會在一定程度上改變腦網絡的拓撲結構,因此在未來的研究中還需要進一步研究不同平滑程度對網絡的影響。一個好的平滑參數應該是可以在降低噪聲的同時,具有較高的網絡檢測敏感性,可以檢測出各種其它因素(如年齡,疾病等)對腦網絡的影響。Hub的研究結果顯示,對于不同個體,網絡節點的重要性具有較大的特異性和較小的相似度,這可能是由于多種原因造成的。首先,小鼠MRM圖像的體素大小為人腦的千分之一,圖像的信噪比較低。由于網絡中連接邊的計算是基于各腦區的灰度分布實現的,噪音對于結果的影響可能較大。后期工作一方面需要通過模擬仿真研究信噪比對網絡特異性的影響,另一方面可以加長對活體小鼠圖像采集時間,或采用離體小鼠圖像,提高圖像的信噪比。其次,采用的數據庫中的小鼠圖像前期已經做過灰度歸一化,這部分可能存在舍入誤差,對網絡分析結果造成一定影響。
當然,本研究還存在一些不足之處。首先,基于圖像灰度建立的結構網絡,雖然在一定程度上可以反映腦區中神經細胞的分布信息,但并不能反映小鼠大腦中各個腦區的形態變化。后期可能需要通過變形域對圖像進行調制,使得網絡可以反映形態學的變化。另外,研究對象的腦區劃分并沒有區分左右腦,在下一步的研究中筆者希望將腦區劃分為左右腦,進一步研究左右腦的協同化工作以及腦區間存在的長程連接。
[1]Sporns O,Tononi G,K?tter R.The human connectome: A structural description of the human brain[J].Plos Computational Biology, 2005,1(4):e42.
[2]Sporns O.The human connectome: origins and challenges[J].Neuroimage,2013,80(1):53-61.
[3]Sporns O.Networks of the Brain[M].USA:MIT Press,2011:18578-18578.
[4]林嵐,付振榮,張柏雯,等.DTI腦連接組在大腦疾病方面的研究進展[J].中國醫療設備,2015,30(6):1-6.
[5]Sheline YI,Price JL,Yan Z,et al.Resting-state functional MRI in depression unmasks increased connectivity between networks via the dorsal nexus[J].P Natl Acad Sci USA,2010,107(24):11020-11025.
[6]林嵐,張柏雯,徐小亭,等.大腦老化對大規模皮層網絡拓撲結構的影響[J].中國醫療設備,2015,30(6):12-17.
[7]Scheeringa R,Petersson KM,Oostenveld R,et al.Trial-by-trial coupling between EEG and BOLD identif es networks related to alpha and theta EEG power increases during working memory maintenance[J].Neuroimage,2009,44(3):1224-1238.
[8]De PF,Della PS,Snyder AZ,et al.Temporal dynamics of spontaneous MEG activity in brain networks[J].P Natl Acad Sci USA,2010,107 (13):6040-6045.
[9]Petersen SE,Sporns O.Brain Networks and Cognitive Architectures[J].Neuron,2015,88(1):207-219.
[10]Lin L,Jin C,Fu Z,et al.Predicting healthy older adult’s brain age based on structural connectivity networks using artificial neural networks[J].Comput Meth Prog Bio,2016,125:8-17.
[11]Franke K,Ziegler G,Kl?ppel S,et al.Estimating the age of healthy subjects from T1-weighted MRI scans using kernel methods: Exploring the influence of various parameters[J].Neuroimage,2010,50(3):883-892.
[12]Seeley WW,Crawford RK,Zhou J,et al.Neurodegenerative diseases target large-scale human brain networks[J].Neuron,2009,62(1):42-52.
[13]Supekar K,Menon V,Rubin D,et al.Network Analysis of Intrinsic Functional Brain Connectivity in Alzheimer’s Disease[J].Plos Comput Bio,2008,4(6):1-11.
[14]Bassett DS,Bullmore ET.Human Brain Networks in Health and Disease[J].Curr Opin Neurol,2009,22(4):340-347.
[15]Alexander GE,Bergfield KL,Chen K,et al.Gray matter network associated with risk for Alzheimer’s disease in young to middle-aged adults[J].Neurobiol Aging,2012,33(12):2723.
[16]He Y,Chen ZJ,Evans AC.Small-World Anatomical Networks in the Human Brain Revealed by Cortical Thickness from MRI[J].Cerebral Cortex,2007,17(10):2407-2419.
[17]Tijms BM,Seriès P,Willshaw DJ,et al.Similarity-Based Extraction of Individual Networks from Gray Matter MRI Scans[J].Cerebral Cortex, 2012,22(7):1530-1541.
[18]Kong XZ,Wang X,Huang L,et al.Measuring individual morphological relationship of cortical regions[J].J Neurosci Meth,2014,237:103-107.
[19]Benveniste H,Blackband S.MR microscopy and high resolution small animal MRI: applications in neuroscience research[J].Prog Neurobiol,2002,67(5):393-420.
[20]Lin L,Fu Z,Xu X,et al.Mouse brain magnetic resonance microscopy: Applications in Alzheimer disease[J].Microsc Res Techniq, 2015,78(5):416-424.
[21]Wang H,Jin X,Zhang Y,et al.Single-subject morphological brain networks: connectivity mapping,topological characterization and test–retest reliability[J].Brain Behav,2016,6(4):448.
[22]Bassett DS,Bullmore E,Verchinski BA,et al.Hierarchical organization of human cortical networks in health and schizophrenia[J].J Neurosci,2008,28(37): 9239-9248.
[23]Bai J,Trinh TLH,Chuang KH,et al.Atlas-based automatic mouse brain image segmentation revisited: model complexity vs.image registration[J].Magn Reson Imag,2012,30(6):789-798.
[24]Lin L,Wu S,Bin G,et al.Intensity inhomogeneity correction using N3 on mouse brain magnetic resonance microscopy[J].J Neuroim aging,2013,23(4):502-507.
[25]林嵐,郝冬梅,白燕萍,等.7T MRI 系統在腦圖像中的應用研究進展[J].生物醫學工程學雜志,2013,30(5):1127-1130.
[26]Sled JG,Zijdenbos AP,Evans AC.A nonparametric method for automatic correction of intensity nonuniformity in MRI data[J].IEEE Trans Med Imag,1998,17(1):87-97.
[27]Paxinos G,Franklin KBJ.The mouse brain in stereotaxic coordinates[M].Oxford:Gulf Professional Publishing,2004.
[28]MacKenzie-Graham A,Lee EF,Dinov ID,et al.A multimodal, multidimensional atlas of the C57BL/6J mouse brain[J].J Anat, 2004,204(2):93-102.
[29]Wang J,Wang X,Xia M,et al.GRETNA: a graph theoretical network analysis toolbox for imaging connectomics[J].Front Human Neurosci,2015,9:386.
[30]Wang JH,Zuo XN,Gohel S,et al.Graph Theoretical Analysis of Functional Brain Networks: Test-Retest Evaluation on Shortand Long-Term Resting-State Functional MRI Data[J].Plos One,2011,6(7):e21976-e21976.
[31]Maslov S,Sneppen K.Specificity and stability in topology of protein networks[J].Science,2002,296(5569):910-913.
[32]Bassett DS,Bullmore E.Small-World Brain Networks[J].Neurosci
entist,2006,12(6):512-523.
[33]Achard S,Salvador R,Whitcher B,et al.A resilient,low-frequency, small-world human brain functional network with highly connected association cortical hubs[J].J Neurosci,2006,(26):63-72.
[34]Batalle D,Eixarch E,Figueras F,et al.Altered small-world topology of structural brain networks in infants with intrauterine growth restriction and its association with later neurodevelopmental outcome[J].Neuroimage,2012,60(2):1352-1366.
[35]Jin C,Lin L,Kuo LW,et al.The relationships between the identif ed critical nodes within DTI-based brain structural network using hub measurements and vulnerability measurement[C].37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC).IEEE,2015:422-425.
[36]Fu Z,Lin L,Jin C.Symmetric image normalization for mouse brain magnetic resonance microscopy[C].International Conference on Advances in Mechanical Engineering and Industrial Informatics,
Study on the Construction of Individual Structure Network Based on Gray Level Distribution of Mouse Brain MRM Image
The Structure Magnetic Resource Imaging (sMRI) network analysis, one of the most popular techniques for neuroimaging analysis, which integrates the techniques of the sMRI, imaging processing, complex networks and graph theory analysis et al, gives us a good understanding of the brain structural organization and its relationship with diseases. Nowadays, the network analysis for human brain based on sMRI has attracted researchers’ attention, while the study for mouse brain magnetic resource microscopy is relatively rare. In this paper, the individual mouse structural networks were constructed based on Kullback-Leibler divergence of regional intensity distribution and a further analysis of network topology properties was made. This method can be used to study the age and disease related brain network changes in mouse brain.
mouse brain; individual brain network; Kullback-Leibler divergence
FU Zhen-rong, LIN Lan, ZHANG Bai-wen, WU Shui-cai
College of Life Science and Bioengineering, Beijing University of Technology, Beijing 100124, China
R445.2
A
10.3969/j.issn.1674-1633.2016.12.004
1674-1633(2016)12-0017-06
2016-11-15
北京市自然科學基金資助項目(7143171);國家科技支撐計劃課題(2015BAI02B03)。
林嵐,副教授。
通訊作者郵箱:lanlin@bjut.edu.cn
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
口腔護理用品工業(2021年4期)2021-11-02 08:22:56
學苑創造·A版(2020年9期)2020-10-13 09:41:02
機械工業標準化與質量(2018年5期)2018-05-30 09:48:17
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國公路(2017年9期)2017-07-25 13:26:38
質量與標準化(2015年7期)2015-07-12 12:21:02
汽車維修與保養(2015年8期)2015-04-17 03:32:51
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國質量與標準導報(2014年9期)2014-02-28 22:25:45
