沙眼衣原體pORF5質粒蛋白免疫優勢片段的篩選與鑒定①
2016-02-24 00:51:33何戰勝粟勝梅雷文波李忠玉
中國免疫學雜志 2016年1期
何戰勝 鄒 燕 粟勝梅 雷文波 李忠玉
(南華大學醫學院病原生物學研究所/特殊病原體防控湖南省重點實驗室,衡陽421001)
?
沙眼衣原體pORF5質粒蛋白免疫優勢片段的篩選與鑒定①
何戰勝鄒燕②粟勝梅雷文波李忠玉
(南華大學醫學院病原生物學研究所/特殊病原體防控湖南省重點實驗室,衡陽421001)
[摘要]目的:鑒定沙眼衣原體pORF5質粒蛋白的免疫原性,并進一步篩選和確定pORF5質粒蛋白免疫優勢片段。方法:以沙眼衣原體D血清型DNA為模板,設計pORF5基因全長和9個不同片段特異引物進行PCR擴增,PCR產物經BamHⅠ、NotⅠ雙酶切后插入經同樣雙酶切的原核表達載體pGEX-6p中,構建pORF5質粒蛋白不同長度片段的原核表達重組體,重組體經PCR和測序鑒定后,轉化XL1 Blue大腸桿菌表達10種不同長度的GST融合蛋白;ELISA方法檢測pORF5質粒蛋白的免疫原性,Western blot 鑒定pORF5質粒蛋白的免疫反應性;ELISA測定10個不同片段與沙眼衣原體生殖道感染患者血清、鼠免疫血清以及抗pORF5單克隆抗體的免疫反應性,鑒定pORF5質粒蛋白免疫優勢片段。結果:pORF5質粒蛋白免疫原性強,能刺激機體產生高效價抗體;破壞pORF5質粒蛋白天然空間結構,其免疫反應性基本消失;在ELISA反應中,N端缺失66氨基酸的F6片段的免疫反應強度與pORF5全長基本相似,F2與F3出現較弱的免疫反應,其余片段免疫反應消失。結論:pORF5質粒蛋白為構象依賴性免疫優勢抗原,其免疫優勢表位和構象表位位于C端,本研究為進一步探討pORF5質粒蛋白的生物學功能和疫苗的研制提供實驗依據。
沙眼衣原體(Chlamydia trachomatis,Ct)泌尿生殖道感染是一種嚴重危害公眾健康的細菌性感染疾病。自上世紀九十年代以來,Ct已超過淋球菌而成為泌尿生殖道感染性疾病的主要性病病原體。Ct生殖道感染不僅引起女性盆腔炎、不孕和異位妊娠等,也可引起男性尿道炎、附睪炎、睪丸炎和前列腺炎等[1],同時Ct還可增加HIV[2,3]及宮頸癌[4-6]的發病率。由于Ct感染臨床癥狀不明顯,致使該疾病不易被發現而得不到及時治療,造成Ct在宿主體內持續存在引起嚴重并發癥。因此,闡明Ct致病機制,尋找免疫優勢抗原研制出有效疫苗,是預防和控制Ct感染性疾病的關鍵。
Ct 共有A~L 19個血清型,每個血清型幾乎均存在一種約7.5 kb大小的隱蔽性質粒,該質粒共有8個開放閱讀框架(Open reading frame,ORF),編碼8種質粒蛋白。在8種質粒蛋白中,pORF5是唯一一種分泌性質粒蛋白[7]。在自然感染狀態下,pORF5質粒蛋白基因均被激活產生內源性靶蛋白。有研究發現Ct患者血清中存在高滴度的特異性的pORF5抗體,提示pORF5可能為免疫優勢抗原[8,9]。為進一步鑒定pORF5質粒蛋白的免疫原性,篩選和確定pORF5質粒蛋白免疫優勢片段,本研究采用基因重組技術構建Ct質粒蛋白pORF5全長和9個不同長度的原核表達重組體,將重組質粒轉化宿主菌XL1 Blue,并對表達的蛋白進行免疫原性研究,本研究為Ct基因工程疫苗和免疫診斷試劑的研制提供實驗依據。
1材料與方法
1.1主要材料Ct D標準株、XL1-Blue大腸桿菌和pGEX-6p原核表達載體為本研究所保存;限制性內切酶BamHⅠ、NotⅠ、pfxDNA聚合酶、蛋白分子量標準等為Invitrogen公司產品;QuickClean PCR 純化試劑盒、質粒提取試劑盒購自美國Qiagen公司;Glutathione Sepharose 4B Beads為Phatnacia 公司產品。
1.2pORF5不同片段基因克隆及原表達重組體的構建根據GenBank提供的Ct pORF5基因序列設計10對引物(表1),并在各對引物的5′加上BamH Ⅰ酶、Not Ⅰ酶切位點及保護堿基。PCR方法擴增pORF5基因全長(FL)和9個不同長度的pORF5基因片段(F1~F9),PCR產物經BamHⅠ、NotⅠ酶切后,連接至經相同酶消化后的pGEX-6p載體中,構建pGEX-6p/pORF5-FL、F1~F9原核表達重組體,重組體轉化感受態大腸桿菌XL1 Blue,在含氨芐青霉素的LB培養基上挑取單個菌落,小量提取質粒,PCR篩選和DNA測序鑒定。
表1pORF5基因不同片段引物
Tab.1Primers for different fragments of pORF5 gene

Name(Length)PrimersequencesFL(M1-A264)5'-CGC-G▼GATCC-ATGGGAAATTCTGGTTTTTATTTG-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTAAGCGTTTGTTTGAGGTATTA-3'F1(M1-S66)5'-CGC-G▼GATCC-ATGGGAAATTCTGGTTTTTATTTG-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTAAGAAGCATTGGTTGATGAATT-3'F2(M1-N132)5'-CGC-G▼GATCC-ATGGGAAATTCTGGTTTTTATTTG-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTAGTTGCATTGAATTTTATTAGTG-3'F3(M1-S198)5'-CGC-G▼GATCC-ATGGGAAATTCTGGTTTTTATTTG-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTATGAGTATCCATAACTAATCG-3'F4(I67-N132)5'-CGC-G▼GATCC-ATTACAATTGGTTTGGTAGCGG-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTAGTTGCATTGATTTTATTAGTG-3'F5(I67-S198)5'-CGC-G▼GATCC-ATTACAATTGGTTTGGTAGCGG-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTATGAGTATCCATAACTAATCG-3'F6(I67-A264)5'-CGC-G▼GATCC-ATTACAATTGGTTTGGTAGCGG-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTAAGCGTTTGTTTGAGGTATTA-3'F7(G133-S198)5'-CGC-G▼GATCC-GGGTTATTCACTCCCAGTAAC-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTATGAGTATCCATAACTAATCG-3'F8(G133-A264)5'-CGC-G▼GATCC-GGGTTATTCACTCCCAGTAAC-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTAAGCGTTTGTTTGAGGTATTA-3'F9(S199-A264)5'-CGC-G▼GATCC-TCAGGCATTCCTAATTTATGTAG-3'5'-TTTTCCTTTT-GC▼GGCCGC-TTAAGCGTTTGTTTGAGGTATTA-3'
1.3pORF5不同長度融合蛋白的表達與純化將鑒定后的陽性克隆接種于含100 μg/ml氨芐的LB培養液中,37℃震蕩培養過夜。次日將過夜培養物以1∶100的比例加入含氨芐的LB培養液中,37℃震蕩培養至OD600約為0.8時,加入終濃度為0.2 mmol/L的IPTG,30℃誘導3 h后低速離心收集細菌,超聲破菌后高速離心收集上清,上清經Glutathione Sepharose 4B Beads純化得到pORF5不同長度的融合蛋白(FL、F1~F9),純化的融合蛋白經12% SDS-PAGE電泳分析,觀察其表達情況和純化效果。
1.4pORF5免疫原性分析將pORF5、CPAF(衣原體蛋白酶樣活性因子)、HSP60(熱休克蛋白60)、MOMP(主要外膜蛋白)融合蛋白進行1∶10稀釋后加入谷胱苷肽包被的酶標板,同時設置GST和空白對照孔,4℃過夜后,0.05%PBST洗滌酶標板以除去未吸附的游離的蛋白抗原;加入2.5%脫脂牛奶室溫封閉1 h,再加入經含GST的大腸桿菌裂解液吸附后的Ct泌尿生殖道感染患者血清,室溫靜置2 h,PBST洗滌后加入1∶2 000稀釋的HRP標記的抗人二抗,室溫2 h;最后加入ABTS顯色劑,室溫避光反應5~10 min后,2 mmol/L H2SO4終止反應,酶標儀讀取450nm波長的A值,當(實驗組A450值-空白組A450值)/(對照組A450值-空白組A450值)≥2.1時為陽性。同時以pORF5、CPAF、HSP60融合蛋白為抗原,以不同稀釋的Ct泌尿生殖道感染患者血清為一抗、HRP標記的羊抗人IgG為二抗,Western blot 分析pORF5質粒蛋白的免疫反應性。
1.5pORF5免疫優勢片段分析將所制備的pORF5 F1~F9融合蛋白及pORF5蛋白全長作為ELISA反應抗原,與酶標板已包被的谷胱苷肽4℃結合過夜,2.5%脫脂牛奶封閉后,分別加入Ct泌尿生殖道感染患者血清、鼠抗GST-pORF5多克隆抗體(pAb)和pORF5 單克隆抗體(mAb)2p和4e6,抗原抗體充分反應后,再分別加入HRP標記的抗鼠、抗人抗體,最后加入顯色劑,酶標儀讀取405nm吸收值,分析pORF5蛋白各個片段的免疫反應性。
2結果
2.1pGEX-6p/pORF5-FL與pGEX-6p/pORF5 F1~F9重組質粒的構建和鑒定以Ct D血清型DNA為模版,應用10對特異性引物對pORF5不同長度基因片段進行擴增,結果擴增出了特異性目的基因片段,與預期目的基因片段大小相符(圖1)。pGEX-6p/pORF5-FL及pGEX-6p/pORF5 F1~F9重組質粒經BamHⅠ和NotⅠ雙酶切后,可切出2個片段,小片段與各目的片段基因大小一致,大片段約4.9 kb,與空質粒大小一致(圖2)。重組質粒測序結果顯示各基因片段與GenBank登陸的D型Ct pORF5質粒堿基序列完全一致。
2.2pORF5各基因片段融合蛋白的表達將pORF5全長和9個不同片段的重組質粒轉化E.coli XL1Blue,IPTG誘導表達10個融合蛋白,Glutathione Sepharose 4B beads 純化融合蛋白,純化的蛋白經SDS-PAGE電泳分析,發現重組質粒菌在各自相應的位置有一明顯蛋白條帶,結果與預期各融合蛋白分子量相符(圖3)。
2.3pORF5 免疫原性分析ELISA結果顯示,pORF5與病人血清出現了強烈的免疫反應,病人血清中存在高滴度pORF5抗體,與傳統的衣原體免疫優勢抗原比較,pORF5 與CPAF免疫原性基本相似,但略高于MOMP、HSP60抗原(圖4)。在Western blot分析pORF5免疫特性時,發現病人血清進行1∶4 000稀釋后,pORF5出現較弱的反應條帶,而CPAF、MOMP和HSP60出現強陽性反應條帶,病人血清做1∶20 000稀釋時,pORF5反應帶消失,而CPAF、MOMP和HSP60蛋白仍可見明顯的條帶(圖5)。
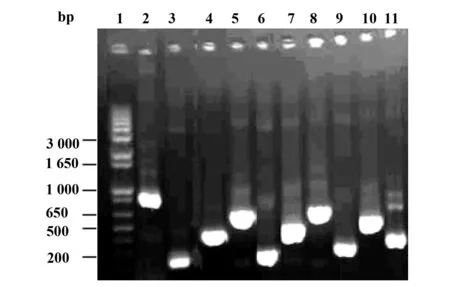
圖1 Ct pORF5質粒基因不同片段PCR擴增產物Fig.1 PCR products of different fragments of pORF5 plasmid gene from Chlamydia trachomatisNote: 1.DNA marker;2.PCR product of pORF5 gene;3.PCR product of pORF5 F1 fragment;4.PCR product of pORF5 F2 fragment;5.PCR product of pORF5 F3 fragment;6.PCR product of pORF5 F4 fragment;7.PCR product of pORF5 F5 fragment;8.PCR product of pORF5 F6 fragment;9.PCR product of pORF5 F7 fragment;10.PCR product of pORF5 F8 fragment;11.PCR product of pORF5 F9 fragment.
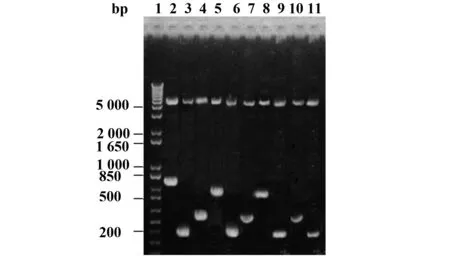
圖2 pGEX-6p/pORF5-FL及pGEX-6p/pORF5 F1~F9重組質粒雙酶切鑒定Fig.2 Identification of pGEX-6p/pORF5-FL and pGEX-6p/pORF5 F1-F9 recombinant plasmids by double digestionNote: 1.DNA marker;2.pGEX-6p/pORF5-FL recombinant plasmid digested with BamHⅠand NotⅠ;3.pGEX-6p/pORF5-F1 recombinant plasmid digested with BamHⅠand NotⅠ;4.pGEX-6p/pORF5-F2 recombinant plasmid digested with BamHⅠand NotⅠ;5.pGEX-6p/pORF5-F3 recombinant plasmid digested with BamHⅠand NotⅠ;6.pGEX-6p/pORF5-F4 recombinant plasmid digested with BamHⅠand NotⅠ;7.pGEX-6p/pORF5-F5 recombinant plasmid digested with BamHⅠand NotⅠ;8.pGEX-6p/pORF5-F6 recombinant plasmid digested with BamHⅠand NotⅠ;9.pGEX-6p/pORF5-F7 recombinant plasmid digested with BamHⅠand NotⅠ;10.pGEX-6p/pORF5-F8 recombinant plasmid digested with BamHⅠand NotⅠ;11.pGEX-6p/pORF5-F9 recombinant plasmid digested with BamHⅠand NotⅠ.

圖3 純化pORF5不同片段的SDS-PAGE分析Fig.3 SDS-PAGE analysis of purified pORF5 diff-erent fragmentsNote: 1.Protein marker;2.Purified pORF5 full length;3.Purified pORF5 F1;4.Purified pORF5 F2;5.Purified pORF5 F3;6.Purified pORF5 F4;7.Purified pORF5 F5;8.Purified pORF5 F6;9.Purified pORF5 F7;10.Purified pORF5 F8;11.Purified pORF5 F9.
2.4pORF5質粒蛋白免疫優勢片段分析為篩選pORF5質粒蛋白免疫優勢片段,將pORF5 9個不同片段分別與Ct生殖道感染患者血清、鼠抗pORF5 pAb、pORF5 mAb (2p和4e6)進行ELISA分析,結果顯示:不同pORF5片段與血清反應強度不一,相同的片段在不同的血清中反應強度也不盡相同;N端缺失66氨基酸的F6片段在Ct感染患者血清ELISA反應中最為強烈,與pORF5全長反應強度基本相似,其次是F2、F3片段,F4、F7、F9反應性最弱;來源于GST-pORF5免疫小鼠產生的抗體能強烈識別F6及全長,但也能識別其他各個片段。pORF5 mAb 2p和4e6 ELISA反應模式相同,只識別F6及全長(圖6)。

圖4 ELISA 方法分析pORF5質粒蛋白免疫原性Fig.4 Immunogenic analysis of pORF5 plasmid protein by ELISA method

圖6 pORF5質粒蛋白免疫優勢片段分析Fig.6 Analysis of immunodominant fragment of pORF5 plasmid protein

圖5 Western blot 分析pORF5質粒蛋白免疫原性Fig.5 Immunogenic analysis of pORF5 plasmid protein by Western blot
3討論
Ct除含有約1 Mb的基因組外,各血清型中亦存在一種隱蔽性質粒,質粒不是衣原體生存所必需的,但質粒缺失后,衣原體毒力高度降低,不能上行引起生殖道的病變[10-12],提示質粒與衣原體毒力密切相關。在質粒編碼的8種質粒蛋白中,pORF5是唯一一種分布于宿主細胞胞漿的分泌性質粒蛋白,該蛋白在Ct感染12 h后可在感染細胞胞漿中檢測其表達,隨著時間的延長,表達量逐漸增加,直至維持到整個細胞發育周期[7]。已有研究證實pORF5可通過誘生炎癥因子、拮抗LL37抗菌肽活性等多種機制參與衣原體的致病過程[13-17]。
pORF5質粒蛋白由264氨基酸編碼,生物信息學分析該蛋白缺乏信號肽序列,具有多個親水性區域,提示可能存在多個抗原表位。Donati等研究發現衣原體感染的患者血清中存在高滴度的特異性抗體[8,9],為探討pORF5質粒蛋白的免疫原性,本研究采用基因重組技術構建Ct質粒蛋白pORF5原核表達重組體,將重組質粒轉化宿主菌XL1 Blue,經IPTG誘導以及Glutathione Sepharose 4B Beads純化后制備了pORF5融合蛋白。采用ELISA方法檢測Ct泌尿生殖道感染患者血清中抗體的滴度,結果發現pORF5抗原與患者血清出現強烈的陽性免疫反應,其免疫反應要高于經典的Ct免疫優勢抗原MOMP、HSP60,與CPAF基本相似; pORF5不僅與全部患者血清發生反應,而且刺激機體產生抗體的效價最高,提示pORF5可作為一種新的衣原體免疫診斷試劑,有望應用于Ct感染的實驗室診斷,同時pORF5亦作為一種新的免疫優勢抗原,可為衣原體疫苗的研制提供一個新的潛在靶抗原。
雖然pORF5 與患者的血清在ELISA反應中出現了強烈的免疫反應,但是當用相同的抗原與抗體進行Western blot 分析時,卻顯示出較弱的免疫反應,其反應強度遠低于MOMP、HSP60和CPAF,其原因可能是在進行Western blot 分析時,抗原需要經過加熱及SDS作用,破壞了pORF5的天然空間構象,而蛋白質的一級、二級、三級和四級結構決定了它的物理、化學、生物化學、物理化學和生物學性質,空間構象改變,影響抗原與抗體的結合。提示pORF5存在構象依賴性表位,pORF5是一種構象依賴性免疫優勢抗原。
為進一步篩選和確定pORF5抗原免疫優勢片段,我們將pORF5抗原分成9個片段,在pORF5 9個片段與Ct生殖道感染患者血清、pORF5多克隆抗體、pORF5單克隆抗體的反應中,發現不同pORF5片段與血清反應強度不一,相同的片段在不同的血清中反應強度也不盡相同;N端缺失66氨基酸的F6片段在Ct感染患者血清ELISA反應中最為強烈,與pORF5全長反應強度基本相似,其次是F2、F3片段,F4、F7、F9反應性最弱;來源于GST-pORF5免疫小鼠產生的抗體能強烈識別F6及全長,但也能識別其他各個片段,提示pORF5 9個片段都具有免疫原性。但當去除F6蛋白N端或C端66個氨基酸時,其免疫反應性大大的減弱,說明了pORF5 抗原優勢表位主要位于C端。
本實驗證實pORF5質粒蛋白為Ct免疫優勢抗原,在自然感染狀態下,pORF5質粒蛋白可誘導機體產生高滴度抗體,具有強烈的免疫原性,其免疫優勢表位位于C端;同時pORF5質粒蛋白具有強烈的免疫反應性,pORF5質粒蛋白對抗體的識別呈高度的構像依賴性,證實pORF5質粒蛋白是一種構象依賴性免疫優勢抗原。本研究為進一步闡明Ct pORF5質粒蛋白的功能,Ct基因工程疫苗和免疫診斷試劑的研制提供實驗依據。
參考文獻:
[1]Gallegos G,Ramos B,Santiso R,etal.Sperm DNA fragmentation in infertile men with genitourinary infection by Chlamydia trachomatis and Mycoplasma[J].Fertil Steril,2008,90(2):328-334.
[2]Schust DJ,Ibana JA,Buckner LR,etal.Potential mechanisms for increased HIV-1 transmission across the endocervical epithelium during C.trachomatis infection[J].Curr HIV Res,2012,10(3):218-227.
[3]Sperling R,Kraus TA,Ding J,etal.Differential profiles of immune mediators and in vitro HIV infectivity between endocervical and vaginal secretions from women with Chlamydia trachomatis infection:a pilot study[J].J Reprod Immunol,2013,99(1/2):80-87.
[4]Seraceni S,De Seta F,Colli C,etal. High prevalence of hpv multiple genotypes in women with persistent chlamydia trachomatis infection [J]. Infect Agent Cancer,2014,9:30.
[5]Deluca GD,Basiletti J,Schelover E,etal.Chlamydia trachomatis as a probable cofactor in human papillomavirus infection in aboriginal women from northeastern Argentina[J].Braz J Infect Dis,2012,15(6):567-572.
[6]Bellaminutti S,Seraceni S,De Seta F,etal.HPV and chlamydia trachomatis co-detection in young asymptomatic women from high incidence area for cervical cancer[J].J Med Virol,2014,86(11):1920-1925.
[7]Li Z,Chen D,Zhong Y,etal.The chlamydial plasmid-encoded protein pgp3 is secreted into the cytosol of Chlamydia-infected cells[J].Infect Immun,2008,76(8):3415-3428.
[8]Donati M,Laroucau K,Storni E,etal.Serological response to pgp3 protein in animal and human chlamydial infections[J].Vet Microbiol,2009,135(1/2):181-185.
[9]Horner P,Soldan K,Vieira SM,etal.C.trachomatis Pgp3 antibody prevalence in young women in England,1993-2010[J].PLoS One,2013,8(8):e72001.
[10]O′connell CM,Ingalls RR,Andrews CW,etal.Plasmid-deficient Chlamydia muridarum fail to induce immune pathology and protect against oviduct disease[J].J Immunol,2007,179(6):4027-4034.
[11]Frazer LC,Darville T,Chandra-Kuntal K,etal.Plasmid-cured Chlamydia caviae activates TLR2-dependent signaling and retains virulence in the Guinea pig model of genital tract infection[J].PLoS One,2012,7(1):e30747.
[12]O′connell CM,Abdelrahman YM,Green E,etal.Toll-like receptor 2 activation by Chlamydia trachomatis is plasmid dependent,and plasmid-responsive chromosomal loci are coordinately regulated in response to glucose limitation by C.trachomatis but not by C.muridarum[J].Infect Immun,2011,79(3):1044-1056.
[13]鄧紅玉,李忠玉,吳移謀,等.沙眼衣原體pORF5質粒蛋白誘發小鼠生殖道免疫損傷初步研究[J].中華微生物學和免疫學雜志,2013,33(2):107-111.
[14]Zhou H,Huang Q,Li Z,etal.PORF5 plasmid protein of Chlamydia trachomatis induces MAPK-mediated pro-inflammatory cytokines via TLR2 activation in THP-1 cells[J].Sci China Life Sci,2013,56(5):460-466.
[15]Liu Y,Huang Y,Yang Z,etal.Plasmid-encoded Pgp3 is a major virulence factor for Chlamydia muridarum to induce hydrosalpinx in mice[J].Infect Immun,2014,82(12):5327-5335.
[16]馬康康,李忠玉,粟盛梅,等.pORF5質粒蛋白拮抗LL37抗菌肽增強沙眼衣原體感染的初步研究[J].中華皮膚科雜志,2014,47(10):703-707.
[17]曹文娟,戴文婷,楊曉玉,等.沙眼衣原體pORF5質粒蛋白激活NALP3炎性復合體誘導THP-細胞產生IL-1β和IL-18[J].中國免疫學雜志,2015,31(5):590-594.
[收稿2015-07-10修回2015-08-17]
(編輯張曉舟)
[關鍵詞]沙眼衣原體;pORF5質粒蛋白;免疫原性;免疫優勢表位
Screen and identification of immunodominant fragment of pORF5 plasmid protein from Chlamydia trachomatis
HEZhan-Sheng,ZOUYan,SUSheng-Mei,LEIWen-Bo,LIZhong-Yu.PathogenicBiologyInstitute,SchoolofMedicine,UniversityofSouthChina/HunanProvincialKeyLaboratoryforSpecialPathogensPreventionandControl,Hengyang421001,China
[Abstract]Objective:To investigate the immunogenicity of pORF5 plasmid protein,and further to screen and identify its immunodominant domian.Methods: 10 different fragments of pORF5 gene including full length were amplified from the DNA of Chlamydia trachomatis serovar D by PCR and cloned into appropriate site of pGEX-6p vector to construct recombinant vectors after digested with BamHⅠ and NotⅠ restriction endonucleases.After identification by PCR and sequencing,the recombinant plasmids were transformed into XL1 Blue E.coli to express the GST fusion proteins.ELISA and Western blot were carried out to identify the immunogenicity and immunoreaction of pORF5 plasmid protein.10 different fragments were reacted with sera from patients urogenitally infected with Chlamydia trachomatis,mouse polyclonal antibodies and mouse monoclonal antibodies of pORF5 plasmid protein with ELISA method.Results: pORF5 plasmid protein displayed strong immunogenicity and could induce a strong antibody response in human.The reactivity of human antibodies almost completely disappeared,when the native structure of pORF5 plasmid protein was destroyed.F6 that only lacked the N-terminal 66 amino acids was recognized by antibodies in ELISA as strongly as the whole pORF5 plasmid protein was.However,no other fragments were significantly recognized although there was a minimal reactivity of F2 and F3 with antibodies.Conclusion: pORF5 plasmid protein was an immunodominant antigen containing conformation-dependent epitope,and the C-terminal three quarters of pORF5 amino acid sequence was required for maintaining its immune dominance and conformation.The significance of the above findings lay a foundation for the further study on pORF5 protein function and vaccine development.
[Key words]Chlamydia trachomatis;pORF5 plasmid protein;Immunogenicity;Immunodominant epitope
通訊作者及指導教師:李忠玉(1972年-),女,教授,主要從事衣原體致病機制與防治研究,E-mail:lzhy1023@hotmail.com。
作者簡介:何戰勝(1971年-),女,主要從事病原體致病機制研究,E-mail:2241332533@qq.com。
中圖分類號R374+1
①本文為國家自然科學基金(No.81102230、No.31470277)、湖南省高校創新平臺開放基金(13K081)、湖南省重點研發計劃(2015JC3087)、湖南省高層次衛生人才“225”工程培養項目(2013-13)、湖南省普通高校帶頭人培養項目(2014-186)、特殊病原體防控湖南省重點實驗室(2014-5)和湖南省高等學校“分子靶標新藥研究”協同創新中心(2014-405)資助項目。
②共同第一作者。
文獻標志碼A
文章編號1000-484X(2016)01-0059-06
doi:10.3969/j.issn.1000-484X.2016.01.013
·免疫學技術與方法·