低鹽水體下攝食水平對斜帶石斑魚幼魚生長及能量收支的影響
2016-02-09 02:29:26黃建盛王安利張健東王忠良湯保貴
海洋科學 2016年11期
黃建盛, 王安利, 陳 剛, 張健東, 王忠良, 湯保貴
(1. 廣東海洋大學 水產學院, 廣東 湛江 524025; 2. 華南師范大學 生命科學學院, 廣東 廣州 510631)
低鹽水體下攝食水平對斜帶石斑魚幼魚生長及能量收支的影響
黃建盛1, 王安利2, 陳 剛1, 張健東1, 王忠良1, 湯保貴1
(1. 廣東海洋大學 水產學院, 廣東 湛江 524025; 2. 華南師范大學 生命科學學院, 廣東 廣州 510631)
為探討斜帶石斑魚(Epinephelus coioides)在低鹽水體下最適的攝食水平, 在水體鹽度12、溫度28℃±1℃的條件下, 作者投喂人工配合飼料, 研究了不同攝食水平(分別為體質量的2.36%、2.83%、3.31%、3.79%、4.25%及飽食)對斜帶石斑魚幼魚(初始體質量為29.28 g±2.63 g)生長和能量收支的影響,并建立了生長-攝食關系和不同攝食水平下的能量收支方程。結果表明, 在低鹽下幼魚魚體粗蛋白及灰分含量在各攝食水平組間無顯著差異(P>0.05); 脂肪含量在3.79%、4.25%及飽食組顯著高于其他攝食水平組(P<0.05); 干物質含量在4.25%、飽食組顯著高于其他攝食水平組(P<0.05); 能量含量隨攝食水平降低而下降。攝食水平對幼魚攝食率有顯著影響(P<0.05)且隨攝食水平的降低而下降。幼魚濕質量特定生長率、干質量特定生長率、蛋白特定生長率、能量特定生長率均隨著攝食水平的增加而增加。特定生長率與攝食水平呈減速增長曲線關系, 可用對數方程描述。幼魚飼料濕質量轉化效率、飼料干質量轉化效率、飼料蛋白轉化效率均隨攝食水平的增加先增大后減少的變化, 在3.79%組達到最大值,飼料能量轉化效率隨攝食水平的增加呈一直增加, 在飽食組達到最大值。隨著攝食水平的增加, 生長能(G)呈先升后降的變化, 在3.79%組有最大值, 與攝食水平之間可用一元二次方程表述; 排糞能(F)與排泄能(U)呈一直增大的變化, 在飽食組有最大值, 與攝食水平之間均可用線性方程表述; 代謝能(R)則呈降低并逐漸趨于平穩的變化。在3.79%組, 幼魚獲得最佳的能量收支方程為: 100.00C= 12.29F+13.73U+31.33R+42.65G。綜合考慮認為, 斜帶石斑魚幼魚在低鹽水體下最適的攝食水平為體質量的3.79%。
攝食水平; 斜帶石斑魚(Epinephelus coioides); 生長; 能量收支; 低鹽水體
在魚類生物能量學研究中, 攝食水平因子一直都是研究熱門的因子之一。攝食水平與魚類的種類、大小、食性及飼料質量有關, 一般肉食性魚類的攝食量低于雜食性和草食性, 并隨著魚類的生長階段不同, 其攝食水平也會不斷地發生改變[1]。因此確定養殖魚類的最佳攝食水平成為魚類營養學和魚類生態學的研究重點之一。
根據文獻報道的研究結果, 魚類的生長與攝食水平的關系, 主要有兩種模型: 一種呈線性關系[2-5];另一種是減速增長曲線關系[6-9]。對于某一種養殖魚類, 確定其生長-攝食的模型, 具有重要的生產實踐意義。若生長-攝食模型為線性關系的魚類, 其最大的攝食率就是其最佳投喂水平; 若是減速增長曲線上升關系的魚類, 其最佳投喂水平應在接近最大攝食率的攝食水平, 這樣可以避免浪費飼料, 減少養殖成本, 并減輕對養殖環境的污染。從養殖業的角度來看, 目的就是盡可能多的食物能轉化為生產能,提高飼料利用率, 降低代謝消耗, 從而提高養殖的經濟效益和減少對養殖環境的破壞。
關于石斑魚屬(Epinephelus)的生物能量學的研究報道較少, 彭樹鋒等[10]研究了體質量對斜帶石斑魚(Epinephelus coioides)能量收支的影響; 周暉等[11]研究了3種蛋白替代魚粉對褐點石斑魚(Epinephelusfuscoguttatus)能量收支的影響。斜帶石斑魚是廣東、海南、福建等地區的海水網箱養殖的主要品種之一,然而, 近年來養殖業者為了避免沿海地區的惡劣環境因素對生產造成的風險, 逐漸發展池塘低鹽度水體養殖并取得成功。然而尚未見有在低鹽水體下斜帶石斑魚幼魚攝食水平研究相關報道。為此, 本實驗開展攝食水平對斜帶石斑魚幼魚在低鹽水體下生長及能量收支的影響, 所得資料可進一步豐富斜帶石斑魚能量學方面的研究內容并為其在低鹽水體下養殖中投喂模式的優化提供一定的科學依據。
1 材料與方法
1.1 實驗用魚來源與馴化
實驗魚為廣東海洋大學魚類實驗室當年繁育的幼魚, 選取健康、活力好的幼魚1 000尾暫養于室內水泥池(4 m×3 m×1.2 m), 1周后開始分組。暫養期間,持續充氣, 自然光照, 每天上下午飽食投喂2次。溶解氧保持在6 mg/L以上, pH7.8~8.3, 氨氮<0.01 mg/L, 自然海水的鹽度為28~31。
幼魚饑餓2 d, 從暫養水泥池中隨機取出健康、活力好的、個體大小相近的幼魚540尾, 隨機放入18個養殖水槽(70 cm×50 cm×60 cm)中, 充氣, 自然光照, 適應1周后開始逐漸降低鹽度。每天用曝氣后的自來水調節鹽度, 每天降低2~4鹽度至鹽度12左右, 并在此鹽度下養殖2周后開始正式實驗。在此期間估算幼魚最大投喂量, 為正式實驗中設計不同的攝食水平提供依據。在適應階段, 每天飽食投喂2次(8: 00和16: 00)。實驗所用的飼料為廣東越群海洋生物研究開發有限公司生產的石斑魚專用配合飼料(型號: 東丸C6, 營養成分為: 水分9.01%、粗蛋白52.94%、粗脂肪12.21%、粗灰分14.51%)。
1.2 實驗設計與管理
實驗在廣東海洋大學海洋生物研究基地進行。選擇體質量相近的健康、活力好的個體, 平均體質量為29.28 g±2.63 g, 體長為10.59 cm±0.83 cm, 各組初始體質量和體長無顯著差異(P>0.05)。采用隨機分組試驗, 共設置6個處理組, 分別為最大投喂量(飽食)、90%飽食、80%飽食、70%飽食、60%飽食和50%飽食, 修正為以攝食水平表示則分別為4.75%(飽食)、4.25%、3.79%、3.31%、2.83%和2.36%(百分比為每日投喂量占初始試驗魚濕質量的質量百分比)。每個處理3個重復, 每個重復放養30尾魚。每天8: 00和16: 00各投喂1次, 試驗期內每隔2周測量一次各組魚體質量, 相應調整其投喂量。另外稱質量150 g飼料, 分成3份投入水中30 min后回收, 烘干、稱質量計算殘餌損失率, 以此來校正攝食量。每天每缸記錄投喂量, 并在投喂后30 min用虹吸移出殘餌, 烘干并稱質量記錄; 并在投喂1、3 h及投喂前用虹吸移出糞便, 在同一水槽收集集中在一起置于70℃下烘干至恒質量。實驗期間, 連續充氣, 每天換水量50%, 溫度為28℃±1℃, 溶解氧保持在6 mg/L以上, pH7.8, 自然光照。
實驗開始前, 幼魚饑餓2 d, 從暫養池中隨機取30尾魚用丁香酚(100 mg/L)麻醉后稱質量, 作為幼魚初始的體質量值, 隨機分成3份作為初始分析樣本, 70℃下烘干, 用以估算實驗開始時魚體的生化成分及其能值。實驗周期歷時56 d。實驗結束后, 實驗魚饑餓2 d, 用丁香酚(100 mg/L)快速麻醉后, 每缸魚逐尾稱質量, 并每缸隨機選取10尾, 用于實驗后魚體成分分析。
1.3 樣品測定
水分的測定為105℃烘干恒質量法; 粗蛋白的測定為凱氏定氮法(N×6.25, FOSS Kjeltec System 2300, Sweden); 脂肪的測定為索氏抽提法(Extraction System B-811, Switzerland); 粗灰分的測定為箱式電阻爐550℃灼燒法; 能量含量采用IKAC200型熱量儀測定。
1.4 計算方法
1.4.1 攝食和生長指標
攝食率(%)=FI/[(Wt+W0)/2×t]×100
濕質量特定生長率(%/d)=(lnWt-lnW0)/t×100
干質量特定生長率(%/d)=[ln(Wt×CDt)-ln(W0× CD0)]/ t×100
蛋白特定生長率(%/d)=[ln(Wt×CPt)-ln(W0×CP0)]/ t×100
能量特定生長率(%/d)=[ln(Wt×CEt)-ln(W0×CE0)]/ t×100
飼料濕質量轉化效率(%)=(Wt-W0)/FI×100
飼料干質量轉化效率(%)=(Wt×CDt-W0×CD0)/ (FI×CD)×100
飼料蛋白轉化效率(%)=(Wt×CPt-W0×CP0)/(FI× CP)×100
飼料能量轉化效率(%)=(Wt×CEt-W0×CE0)/(FI× CE)×100
上述公式中, FI(g)為平均每尾魚攝食飼料總量,Wt、W0(g)分別為魚體終末和初始濕質量, CDt、CD0(%)分別為終末和初始魚樣平均干物質含量, CPt、CP0(%)分別為終末和初始魚體的平均蛋白質含量, CEt、CE0(kJ/g)分別為終末和初始魚體平均能量含量, CD、CP(%)和CE(kJ/g)分別為飼料的干物質、蛋白質和能量含量, t為實驗天數。
1.4.2 能量收支指標
能量在魚體內轉換基本模型[12]: C=F+U+R+G,攝食能(C)=攝入飼料的干質量×飼料的能量含量;排糞能(F)=糞便干質量×糞便的能量含量; 排泄能(U)依據氮收支平衡式計算[13]: U=(CN-FN-GN) ×24.8,式中, CN為食物中獲取的氮; FN為糞便中損失的氮; GN為魚體中積累的氮; 24.8為每克氨氮的能值(KJ)。這里假定氨是唯一的氮排泄物。生長能(G)=實驗結束時魚體能量-實驗開始時魚體能量。代謝能(R)根據能量收支式R=C-F-U-G計算, 同化能(A)= C-F-U。
1.5 數據分析
數據通過 SPSS統計軟件進行分析, 采用One-way ANOVA過程來檢驗攝食水平對測定指標影響的顯著性, 用Ducan檢驗法進行多重比較, 描述性統計值使用平均值±標準差 (Mean±SD) 表示, P<0.05為顯著性差異。
2 結果
2.1 魚體生化成分
初始魚樣和不同攝食水平飼養56 d后斜帶石斑魚幼魚魚體干物質、粗蛋白、粗脂肪、灰分和能量含量結果見表1。實驗結果表明, 4.25%和飽食組幼魚干物質含量顯著高于其他攝食水平組(P<0.05), 3.79%、3.31%、2.83%和2.36%組之間無顯著性差異(P>0.05);攝食水平對魚體粗蛋白含量和灰分含量無顯著性影響(P>0.05)。隨攝食水平增加, 粗脂肪含量和能量含量呈下降趨勢, 攝食水平對兩者有顯著影響(P<0.05)。
2.2 生長及飼料轉化效率
低鹽水體下斜帶石斑魚幼魚在不同攝食水平的攝食率見表2。由表中的數據顯示, 隨著攝食水平的降低, 攝食率呈下降的趨勢, 統計分析表明, 攝食水平對幼魚的攝食率有顯著性差異(P<0.05)。
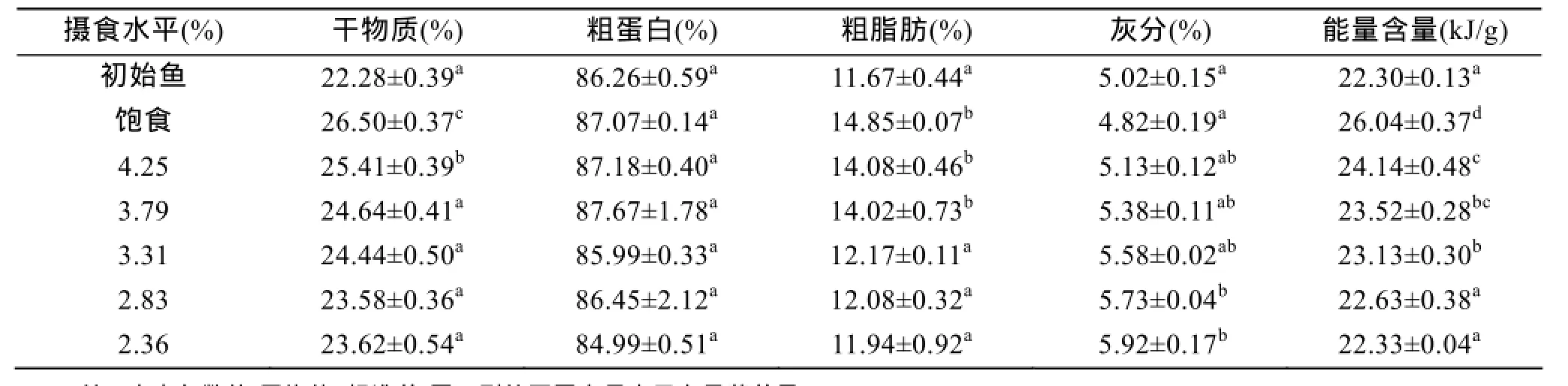
表1 攝食水平對低鹽水體下斜帶石斑魚幼魚魚體生化成分和能量含量的影響Tab. 1 Effect of different feeding levels on the body composition of juvenile E. coioides cultured in low-salt water at the beginning and end of the growth experiment
斜帶石斑魚在6個攝食水平的特定生長率測定數據見表2。結果表明, 濕質量特定生長率、干質量特定生長率、蛋白特定生長率及能量特定生長率均隨著攝食水平的增加而增加。特定生長率與攝食水平呈減速增長曲線關系, 可用對數方程來描述(表3)。令特定生長率=0, 由各回歸方程可得, 斜帶石斑魚在本實驗條件下濕質量、干質量、蛋白和能量維持日糧水平分別為0.93%、1.06%、1.09%和1.27%。
不同攝食水平下斜帶石斑魚飼料轉化效率如表2所示, 飼料濕質量轉化效率、飼料干質量轉化效率、飼料蛋白轉化效率均隨攝食水平增加呈先上升后下降的變化趨勢, 在3.79%組有最大值。飼料能量轉化效率隨攝食水平的增加呈一直增加, 在飽食組達到最大值。
2.3 能量收支
不同攝食水平下的斜帶石斑魚能量收支各組分測定數據見表4所示, 表中數據顯示, 隨著攝食水平的增加, 生長能比例呈增加的趨勢, 到3.79%組時最高, 之后呈下降的趨勢, 方差分析表明, 攝食水平對生長能比例有顯著影響(P<0.05), 其中3.79%、4.25%及飽食組組間無顯著差異(P>0.05), 生長能比例與攝食水平兩者可用一元二次方程來表述(表5)。排糞能比例和排泄能比例均隨攝食水平增加而增加, 方差分析表明, 攝食水平對排糞能比例、排泄能比例有顯著影響(P<0.05), 兩者之間呈線性關系。代謝能比例隨攝食水平的增加而降低, 到3.79%組逐漸趨于平穩(3.79%、4.25%及飽食組組間無顯著性差異)。當能量收支以占同化能比例表示時, 生長能占同化能比值隨攝食水平增加而增加, 在3.79%組達到最大值,能量收支方程為:
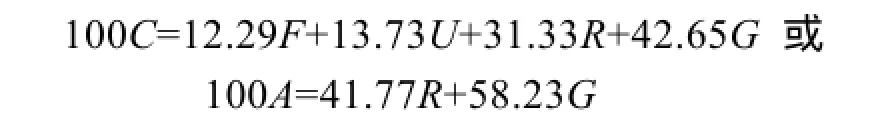
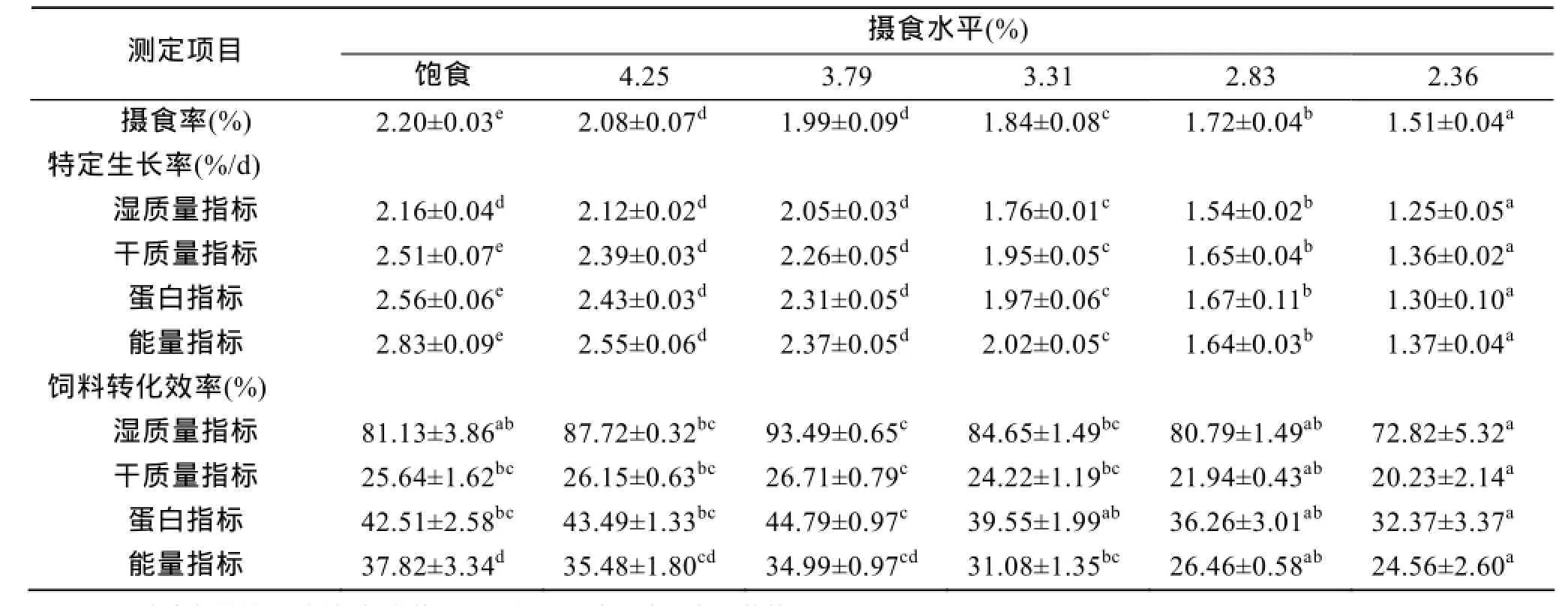
表2 攝食水平對低鹽水體下斜帶石斑魚幼魚生長及飼料轉化效率的影響Tab. 2 Effect of different feeding levels on the growth and feed conversion efficiency of juvenile E. coioides cultured in low-salt water
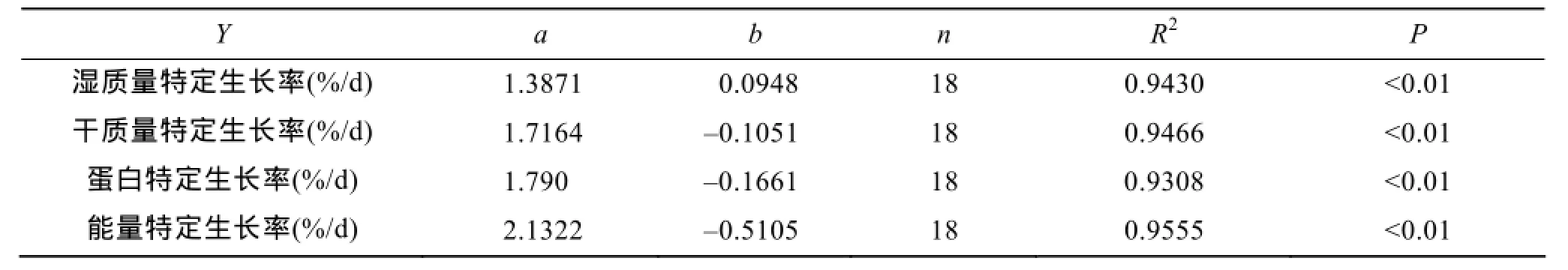
表3 低鹽水體下斜帶石斑魚幼魚特定生長率(RSG)與攝食水平(FL)關系的回歸方程(RSG=aLn(FL)+b)的系數Tab.3 Coefficients for the regression model (RSG= aLn(FL + b)) relating specific growth rate (RSG) to the feeding levels (FLs) of juvenile E. coioides cultured in low-salt water
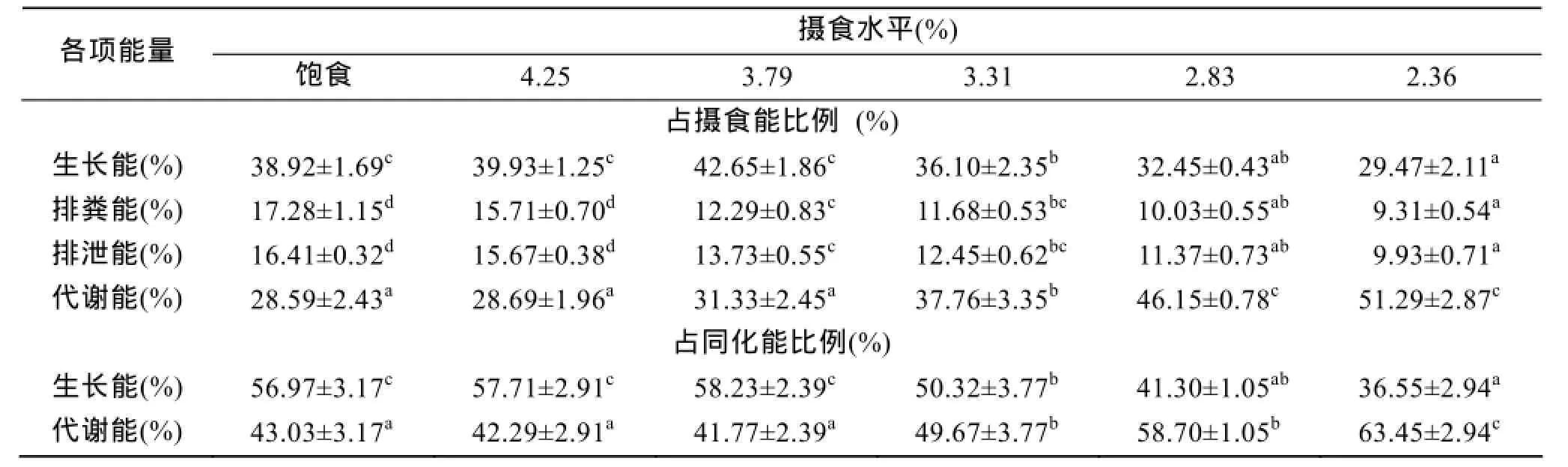
表4 攝食水平對低鹽水體下斜帶石斑魚幼魚能量收支的影響Tab. 4 Effect of different feeding levels on the energy budgets of juvenile E. coioides cultured in low-salt water

表5 攝食水平與生長能比例的回歸方程(Y=aFL2+bFL+c)系數; 與排糞能比例、排泄能比例和代謝能比例的回歸方程(Y=aFL+b)系數Tab. 5 Coefficients of the regression equation (Y = aFL2+ bFL + c) relating to the growth energy content and feeding levels and those of the regression equation (Y = aFL + b) relating to the feces energy content, excretion energy content, metabolism energy content, and feeding levels
3 討論
3.1 魚體成分及能值
根據文獻的報道, 魚體成分受到攝食水平的影響[2,14-16]。之前很多的研究結果指出, 魚體的蛋白質含量與攝食水平無顯著關系, 而魚體的干物質含量、脂肪含量及能值含量受到攝食水平的顯著影響, 均隨著攝食水平的增加而增加, 灰分含量隨攝食水平的增加而減少[1,14-16]。本研究結果表明, 在低鹽水體下斜帶石斑魚幼魚魚體蛋白含量與攝食水平的關系無顯著關系, 魚體脂肪含量隨投喂水平增加而增加,而魚體灰分含量則相反, 研究結果與其他研究結果相類似[2,15-16]。分析其原因就是食物促進魚類的生長,同時高的攝食水平, 使得更多的食物能量轉化為魚體脂肪, 這是一個普遍的現象, 而魚體灰分含量與攝食水平呈負相關, 可能是骨骼生長相對較緩慢的原因[17]。
3.2 生長-攝食的關系
文獻報道關于生長-攝食的關系模型主要的兩種類型, 一種是簡單的直線關系, 采用線性方程來表述[2-5], 另一種是減速增長曲線, 采用對數方程來表述[6-9,17]。本實驗中在無饑餓組的情況下, 研究結果表明斜帶石斑魚幼魚在低鹽水體的生長隨著攝食水平的上升呈減速增長曲線關系, 生長-攝食的關系模型可用對數方程來表示。
確定魚類生長-攝食關系是直線還是曲線, 對魚類養殖有重要的意義[18]。當這一關系直線時, 生長率和食物利用率均在最大攝食水平是最高, 最大攝食水平即為最佳攝食水平; 當這一關系為曲線時, 在較高的攝食水平時生長率增加緩慢, 食物利用率相對減少, 因此最佳的飼料投喂水平應該低于最大攝食水平。在養殖實踐中, 要重視生長-攝食的關系, 為了達到最佳的經濟效益, 應該選擇最佳的投喂水平。但往往在實際中還要考慮其他的因素計算所需要的投喂水平, 比如斜帶石斑魚幼魚在池塘或海上網箱無法過冬, 需要移到室內水泥池進行人工加溫越冬,此期間主要考慮因子是海水升溫成本很高, 因此加溫不能過高, 只要在高于其適宜生長溫度下限就可以, 同時換水量不宜太大, 因此在越冬期間不能投喂太多, 否則會引起水質惡化而導致增加換水量,這就需要具體實際情況從能量學的角度綜合生長、攝食水平、水溫、生產成本等各種因素來分析以達到最佳效果。另一個例子, 當斜帶石斑魚市場價格低迷, 養殖戶不愿出售已達商品規格的成品魚, 多數養殖戶選擇等待節假日時間出售, 因此在等待價格回升的期間, 如果按最佳攝食水平投喂則難于承受飼料成本, 很多養殖戶選擇降低投喂水平, 但是要降到什么程度并不一致, 需要從能量學的知識結合經濟承受能力才能作出更好的安排, 以達到好的效果。另外, 魚類生長-攝食關系受到諸多因子的影響,如溫度[19-20]、體質量[21]、餌料種類[7]等, 因此, 有必要進一步開展低鹽水體下斜帶石斑魚的生物能量學研究, 探討環境因子對其生長-攝食的影響, 尤其應關注因子間的互作效應。Montgomery[22]指出在因子互作效應存在的情況下, 往往比因子主效應更重要。在尼羅羅非魚(Oreochromis niloticus)[23]、大菱鲆(Scophthalmus maximus)[24]、犬齒牙鲆(Paralichthys dentatus)[25]、花狼魚(Anarhichas minor)[26]等研究中證實環境因子對魚類生長有明顯的互作效應。孫麗華等[21]通過研究攝食水平和溫度對軍曹魚生長及氮收支的影響, 預測攝食水平和溫度對魚類氮排泄存在一定的交互作用。本研究中, 生長-攝食關系是在室內可控環境下進行的, 實驗室環境和生產實踐的室外池塘養殖環境、工廠化養殖環境存在一定的差距, 如: 換水量、養殖密度、養殖規模等。因此, 為使得研究結果更好地為生產實踐提供依據, 有必要進一步開展調查研究。
3.3 能量收支方程
本實驗結果顯示, 隨攝食水平增加, 低鹽水體下斜帶石斑魚魚體活動代謝降低, 分析其原因可能是在攝食水平較低的情況, 有限的食物資源使得魚類必須依靠持續的游動以增加捕食機會, 因此會導致活動能量的上升, 但高攝食水平情況下, 魚類攝食后特殊動力作用耗能上升, 由于這兩部分能量上的平衡, 使得代謝能比例隨投喂水平的增加而降低并逐漸趨于平穩。在低鹽環境下的斜帶石斑魚糞便能比例和排泄能比例隨著攝食水平的增加而增加。這一結果與文獻報道梭魚(Mulletliza haematocheila)[27]的結果一致。分析其原因是隨著攝食水平的增加, 攝食食物增多, 斜帶石斑魚不能完全消化吸收, 多余的部分被排出體外, 導致糞便增高。當攝食的食物增多時,斜帶石斑魚攝入體內的蛋白質相對增加, 可能是在低鹽條件下動用一部分蛋白質作為能源物質, 氨基酸代謝進而加強, 氨氮排泄升高。關于攝食水平與糞便能、排泄能的關系, 有些資料結果表明, 糞便能、排泄能隨著攝食水平增加而減少[8,28], 而有些資料結果表明糞便能、排泄能隨著攝食水平的增加變化不大[2,27], 這種差異反映了種間差異和實驗方法不同。
能量收支方程給出一定環境條件下能量收支各組分的定量關系。基于已發表的14種魚類能量分配的數據上, 總結了最大攝食條件下魚類的能量收支,得出平均收支式: 100A=60R+40G[29](A為同化能)。在本研究中, 在低鹽水體下斜帶石斑魚幼魚飽食條件下能量收支方程為: 100A=43.03R+56.97G, 與如上平均分配模式比較發現, 斜帶石斑魚的生長能比例高于平均水平, 與褐牙鲆(Paralichthys olivaceus)[30]、梭魚(Mulletliza haematocheila)[27]相類似。斜帶石斑魚為底層定居魚類, 生態習性為喜歡棲息在巖礁、珊瑚礁底, 在實驗過程觀察到其除了攝食, 大部分時間靜臥在缸底或四周角落, 因此生態習性決定其代謝和排泄所消耗能量較少, 而分配于生長的能量相對較多。然而, 在研究魚類的能量收支時, 影響的因素比較多, 比如魚類的品種、實驗對象的體質量、餌料種類、飼料配方、投喂次數、實驗方法以及實驗條件等會導致各研究結果存在很大的差異。因此, 魚類能量分配模式存在很大的差異, 很難總結出魚類能量收支的一般形式, 而在一定溫度范圍內, 最大攝食水平時能量收支可以相互比較, 前提條件是能量收支在這一溫度范圍內不隨溫度而變化[29], 斜帶石斑魚在低鹽水體下是否存在這一現象以及溫度范圍的大小, 有待進一步研究。
[1] 崔奕波. 魚類生物能量學的理論與方法[J]. 水生生物學報, 1989, 13(4): 369-383. Cui Yibo. Bioenergetics of fishes: theory and methods[J]. Acta Hydrobiologica Sinica, 1989, 13(4): 369-383.
[2] 朱曉鳴, 解綏啟, 崔奕波. 攝食水平對異育銀鯽生長及能量收支的影響[J]. 海洋與湖沼, 2000, 31(5): 471-479. Zhu Xiaoming, Xie Shouqi, Cui Yibo. Effect of ration level on growth and energy budget of the gibel carp Carassius Auratus Gibelio[J]. Oceanologia Et Limnologia Sinica, 2000, 31(5): 471-479.
[3] Paul A, Paul J, Smith R. Energy and ration requirements of juvenile Pacific halibut (Hippoglossus stenolepis) based on energy consumption and growth rates[J]. Journal of Fish Biology, 1994, 44(6): 1023-1031.
[4] Cui Y, Hung S, Zhu X. Effect of ration and body size on the energy budget of juvenile white sturgeon[J]. Journal of Fish Biology, 1996, 49(5): 863-876.
[5] Sun L, Chen H, Huang L, et al. Growth, faecal production, nitrogenous excretion and energy budget of juvenile cobia (Rachycentron canadum) relative to feed type and ration level[J]. Aquaculture, 2006, 259(1): 211-221.
[6] Cui Y, Wootton R. Bioenergetics of growth of a cyprinid, Phoxinus phoxinus: the effect of ration, temperature and body size on food consumption, faecal production and nitrogenous excretion[J]. Journal of Fish Biology, 1988, 33(3): 431-443.
[7] 孫麗華, 陳浩如, 黃洪輝, 等. 攝食水平和餌料種類對軍曹魚幼魚生長及氮收支的影響[J]. 熱帶海洋學報, 2010, 29(4): 94-101. Sun Lihua, Chen Haoru, Huang Honghui, et al. Effects of ration level and feed type on growth and nitrogen budget of young cobia (Rachycentron canadum)[J]. Journal of Tropical Oceanography, 2010, 29(4): 94-101.
[8] 黃建盛, 陳剛, 張健東, 等. 攝食水平對卵形鯧鲹幼魚的生長和能量收支的影響[J]. 廣東海洋大學學報, 2010, 30(1): 18-23. Huang Jiansheng, Chen Gang, Zhang Jiandong, et al. Effect of ration of level on growth and energy budget of Trachinotus ovatus juveniles[J]. Journal of Guangdong Ocean University, 2010, 30(1): 18-23.
[9] Makinen T. Effect of temperature and feed ration on energy utilization in large rainbow trout, Oncorhynchus mykiss (Walbaum)[J]. Aquaculture Research, 1994, 25(2): 213-232.
[10] 彭樹鋒, 王云新, 葉富良, 等. 體重對斜帶石斑魚能量收支的影響[J]. 水生生物學報, 2008, 6(6): 934-940. Peng Shufeng, Wang Yunxin, Ye Fuliang, et al. Effects of body weight on energy budget of Epinephelus coioides[J]. Acta Hydrobiologica Sinica, 2008, 6(6): 934-940.
[11] 周暉, 陳剛, 紀多亮, 等. 3 種蛋白替代魚粉對褐點石斑魚生長, 體成分及能量收支的影響[J]. 廣東海洋大學學報, 2013, 32(6): 10-16. Zhou Hui, Chen Gang, Ji Duoliang, et al. The effects of partial replacement of fish meal by three protein sources on growth performance approximate compositions and energy budget of juvenile brown-marbled grouper (Epinephelus fuscoguttatus)[J]. Journal of Guangdong Ocean University, 2013, 32(6): 10-16.
[12] Warren C E, Davis G E. Laboratory studies on the feeding bioenergetics and growth of fishes[M]. Oxford: Blackwell Scientific Publication, 1967: 175-214.
[13] Cui Y, Liu X, Wang S, et al. Growth and energy budget in young grass carp, Ctenopharyngodon idella Val., fed plant and animal diets[J]. Journal of Fish Biology, 1992, 41(2): 231-238.
[14] EI-Saidy D M S D, Gaber M M A. Effect of dietary protein levels and feeding rates on growth performance, production traits and body composition of Nile tilapia, Oreochromis niloticus (L.) cultured in concrete tanks[J]. Aquaculture Research, 2005, 36(2): 163-171.
[15] Ng W K, Lu K S, Hashim R, et al. Effects of feeding rate on growth, feed utilizationand body composition of a tropical bagrid catfish[J]. Aquaculture International, 2000, 8(1): 19-29.
[16] Cho S H, Lee S M, Park B H, et al. Effect of feeding ratio on growth and body composition of juvenile olive flounder Paralichthys olivaceu fed extruded pellets during the summer season[J]. Aquaculture, 2006, 251(1): 78-84.
[17] Han D, Xie S, Lei W, et al. Effect of ration on the growth and energy budget of Chinese longsnout catfish, Leiocassis longirostris Günther[J]. Aquaculture Research, 2004, 35(9): 866-873.
[18] 崔奕波, 解綬啟. 魚類生長變異的生物能量學機制[J].中國科學院院刊, 1998, 6: 453-455. Cui Yibo, Xie Shouqi.Biological energetics mechanism of growth variation in fish[J]. Bulletin of Chinese Academy of Sciences, 1998, 6: 453-455.
[19] 周洪琪, 潘兆龍, 李世欽, 等.攝食和溫度對草魚氮排泄影響的初步研究[J]. 上海水產大學學報, 1999, 8(4): 293-297. Zhou Hongqi, Pan Zhaolong, Li Shiqin, et al. Effect of feeding and temperature on nitrogenous excretion of grass carp, Ctenopharygodon Indells[J]. Joural of Shanghai Fisheries University, 1999, 8(4): 293-297.
[20] 孫麗華, 陳浩如. 溫度和體質量對軍曹魚生長及氮收支的影響[J]. 水產學報, 2013, 37(10): 1527-1534. Sun Lihua, Chen Haoru.Effects of water temperature and fish size on growth and nitrogen budget of cobia (Rachycentron canadum)[J]. Journal of Fisheries of China, 2013, 37(10): 1527-1534.
[21] 孫麗華, 陳浩如. 攝食水平和溫度對軍曹魚幼魚生長及氮收支的影響[J]. 熱帶海洋學報, 2014, 33(2): 37-44. Sun Lihua, Chen Haoru. Effects of ration and temperature on growth and nitrogen budget of young cobia (Rachycentron canadum)[J]. Journal of Tropical Oceanography, 2014, 33(2): 37-44.
[22] Montgomery D C. Design and analysis of experiments[M]. New York: John Wiley & Sons, 2008.
[23] 王輝, 強俊, 王海貞, 等. 溫度與鹽度對吉富品系尼羅羅非魚幼魚能量代謝的聯合效應[J]. 中國水產科學, 2012, 19(1): 51-61. Wang Hui, Qiang Jun, Wang Haizhen, et al. Combined effect of temperature and salinity on energy metabolism of GIFT Nile tilapia (Oreochromis niloticus) juveniles[J]. Journal of Fishery Sciences of China, 2012, 19(1): 51-61.
[24] Imsland A K, Foss A, Gunnarsson S, et al. The interaction of temperature and salinity on growth and food conversion in juvenile turbot (Scophthalmus maximus)[J]. Aquaculture, 2001, 198(3): 353-367.
[25] Malloy K D, Targett T E. Feeding, growth and survival of juvenile summer flounder Paralichthys dentatus: experimental analysis of the effects of temperature and salinity[J]. Marine Ecology Progress Series, 1991, 72(3): 213-223.
[26] Magnussen A B, Imsland A K, Foss A. Interactive effects of different temperatures and salinities on growth, feed conversion efficiency, and blood physiology in juvenile spotted wolffish, Anarhichas minor Olafsen[J]. Journal of the World Aquaculture Society, 2008, 39(6): 804-811.
[27] 線薇薇, 朱鑫華. 攝食水平對梭魚的生長和能量收支的影響 [J]. 海洋與湖沼, 2001, 32(6): 612-620. Xian Weiwei, Zhu Xinhua. Effect of ration size on the growth and energy budget of the mullet Liza haematocheila (T. et S.)[J]. Oceanologia Et Limnologia Sinica, 2001, 32(6): 612-620.
[28] 陳晶, 吳立新, 吳玉波, 等. 不同攝食水平對牙鲆幼魚生長及能量收支的影響[J]. 水產科學, 2012, 31(1): 1-6. Chen Jing, Wu Lixin, Wu Yubo, et al. Effects of ration levels on growth and energy budgets of juvenile japanese flounder, Paralichthys olivaceus[J]. Fisheries Science, 2012, 31(1): 1-6.
[29] Cui Y, Liu J. Comparison of energy budget among six teleosts-III. Growth rate and energy budget[J]. Comparative Biochemistry and Physiology A, Comparative Physiology, 1990, 97(3): 381-384.
[30] 線薇薇, 朱鑫華. 攝食水平對褐牙鲆幼魚能量收支的影響 [J]. 青島海洋大學學報, 2001, 31(5): 695-700. Xian Weiwei, Zhu Xinhua. Effect of ration on energy budget of juvenile bastard halibut Paralichthys olivaceus[J]. Journal of Ocean University of Qingdao, 2001, 31(5): 695-700.
Received: Nov. 6, 2015
Effect of feeding level on the growth and energy budgets of Epinephelus coioides juveniles cultured in low-salt water
HUANG Jian-sheng1, WANG An-li2, CHEN Gang1, ZHANG Jian-dong1, WANG Zhong-liang1, TANG Bao-gui1
(1. Fishery College, Guangdong Ocean University, Zhanjiang 524025, China; 2. College of Life Science, South China Normal University, Guangzhou 510631, China)
Feeding levels; Epinephelus coioides; growth; energy budget; low-salt water
In this paper, we investigate the optimal feeding level of E. coioides juveniles cultured in low-salt water. We observed the effects of feeding levels (satiation, 4.25%, 3.79%, 3.31%, 2.83%, and 2.36%) on the growth and energy budget of these E. coioides juveniles (initial body weight 29.28 ± 2.63 g) cultured in low-salt water (salinity 12) at a temperature of 28 ±1℃; we fed them a formula diet and established the relationship between specific growth rate (SGR), feeding levels, and the energy budget. The results reveal that there was no significant difference in crude protein and ash contents in the groups of juveniles with P > 0.05. The lipid contents of 3.79%, 4.25%, and satiation were significantly higher than those in the other feeding levels (P < 0.05) after culturing for 56 d. The dry matter contents in the 4.25% and satiation groups were higher than those in the other groups. The energy content decreased significantly with a decrease in feeding levels. The feeding ratio of the juveniles was significantly affected by the feeding levels (P < 0.05), and it decreased with a decrease in feeding levels. The specific growth rate in wet weight, dry weight, protein weight, and energy weight increased significantly with feeding levels. We found a logarithmic relationship between SGR and feeding levels. The feed conversion efficiency in wet weight, dry weight, and protein weight first increased and then decreased as feeding levels increased, and the maximum values were observed in the 3.79% group. However, the feed conversion efficiency in energy weight increased with feeding levels, and the maximum value was observed in the satiation group. With increasing feeding levels, the growth energy content (G) of the juveniles first increased and then decreased, and a quadratic relationship was found between G and the feeding levels. With increasing feeding levels, feces energy content (F) and excretion energy content (U) increased, and the maximum values were observed in the satiation group. There was a linear relationship between F, U, and the feeding levels. As the feeding levels increased, the metabolism energy content (R) of the juveniles decreased and tended toward stability. The maximum G was observed in the 3.79% group, and the optimal energy equation was 100.00 C = 12.29 F + 13.73 U + 31.33 R + 42.65 G. We consider the optimal feeding level to be 3.79% of the body weight of E. coioides juveniles cultured in low-salt water.
Q955
A
1000-3096(2016)11-0113-08
10.11759//hykx 20151106003
(本文編輯: 譚雪靜)
2015-11-06;
2016-01-26
廣東高校優秀青年創新人才培養計劃項目(2013LYM0038);廣東省海洋與漁業科技項目(A2009-08D03、A2010-08D04); 廣東海洋大學“創新強校工程”項目(GDOU2016050251); 廣東海洋大學科研啟動費項目(E15183)
[Foundation: Supported by Foundation for Distinguished Young Talents in Higher Education of Guangdong, China, No. 2013LYM0038; Project of Marine and Fishery Science Technology of Guangdong Province, No.A2009-08D03, No. A2010-08D04; Project of Enhancing School with Innovation of Guangdong Ocean University, No.GDOU2016050251; Scientiwfic Research Project of Guangdong Ocean University, No.E15183]
黃建盛(1981-), 男, 廣東湛江人, 講師, 博士, 主要從事海水魚類生理生態學研究, 電話: 0759-2382462, E-mail: huangjs@ gdou.edu.cn; 陳剛, 通信作者, 教授, E-mail: cheng@gdou.edu.cn
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小讀者(2021年2期)2021-03-29 05:03:48
火花(2019年12期)2019-12-26 01:00:28
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
人大建設(2019年12期)2019-05-21 02:55:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
