論鳥類飛行起源的“樹棲”與“地棲”假說之爭
2016-01-26 07:19:36季燕南王旭日黑須球子
地質學刊 2015年2期
季燕南,王旭日,黑須球子,季 強
(1.中國地質環境監測院,北京100081; 2.中國地質科學院地質研究所,北京100037; 3.中國地質大學(北京),北京100083)
論鳥類飛行起源的“樹棲”與“地棲”假說之爭
季燕南1,王旭日2,黑須球子3,季強2
(1.中國地質環境監測院,北京100081; 2.中國地質科學院地質研究所,北京100037; 3.中國地質大學(北京),北京100083)
摘要:簡要介紹了鳥類飛行起源的2種假說:“樹棲起源”假說與“地棲起源”假說。詳細討論了2種假說的理論支撐點及存在的問題,認為鳥類的祖先和原始鳥類可以借助自然界中任何有利的地形和地物來幫助滑翔和起飛,而不是僅僅依賴直立高大的樹木。“地棲起源”假說更合乎情理,也更接近或更符合客觀事實,同時提出了“地棲起源”假說的新模式圖。
關鍵詞:鳥類飛行起源;“樹棲”假說;“地棲”假說;“翼助斜坡奔跑”假說
中圖分類號:Q915.5;Q915.865
文獻標識碼:A
文章編號:1674-3636(2015)02-0201-06
收稿日期:2015-04-26;編輯:陸李萍
基金項目:科學技術部“973”項目(2012CB822004);中國地質調查局地質調查項目(121202114026801)
作者簡介:季燕南(1983—),男,工程師,碩士,主要從事地層古生物和地質遺跡保護研究工作,E-mail:jirod@cags.ac.cn
doi:10.3969/j.issn.1674-3636.2015.02.207
0引言
中國遼西是晚中生代的化石寶庫,以產出豐富的長羽毛恐龍聞名于世。自1996年發現中華龍鳥以來,鳥類起源研究取得了革命性進展,“鳥類是由小型獸腳類恐龍演化而來”的觀點已成為共識,也成為當今國際科學界的主流科學思想。
鳥類的飛行起源與鳥類起源是2種性質完全不同但又有一定關系的科學問題。長期以來,關于鳥類的飛行起源存在2種觀點對立、解釋相背的理論,即“樹棲起源”假說與“地棲起源”假說。
脊椎動物中主要有3種具有翅膀、會飛行的動物:爬行動物中的翼龍、鳥類和哺乳動物中的蝙蝠,它們的翅膀結構各不相同。翼龍的前肢十分發育,一般具有4指,第5指缺失,其第4指(飛行指)極度拉長,與皮膜一起形成翅膀,從而具有飛行能力。蝙蝠的前肢發育5指,第1指短小,其余4指明顯拉長,似“傘骨”狀,與皮膜一起形成翅膀,從而具有了飛行能力(圖1)。鳥類的前肢比較特殊,通過手部骨骼的愈合和消失等變化與羽毛一起形成翅膀。粗壯的肱骨構成上臂;細長的尺骨和橈骨構成前臂;腕骨退化,近端腕骨僅有2塊,遠端腕骨與掌骨愈合成1塊腕掌骨;指骨僅發育第2、3、4指,第2指為小翼羽的著生處。鳥類翅膀的這種結構究竟與“爬樹”有關,還是與其他因素有關?本研究將簡要介紹3種主要假說,并討論鳥類飛行起源各種可能的原因。

圖1 翼龍、鳥類和蝙蝠翅膀的形態結構對比圖Fig.1 Comparison of morphological structures of pterosaurs, birds and bat wings
1“樹棲起源”假說
“樹棲起源”假說最早由Marsh(1880)提出,當時主要受到松鼠、飛蜥、樹蛙等動物可以進行短距離滑翔現象的影響和啟迪。他推測鳥類的祖先或早期鳥類的前肢可能發育了雛形羽毛,經過長期的自然選擇,增強了它們在樹林間跳躍時的滑翔能力,雛形羽毛不斷進化,演變成飛行羽毛,最終獲得了飛行能力。這一假說后來得到了許多學者的認可和支持,其中Heilmann(1926)還給出了鳥類的祖先利用四肢攀爬樹枝的生態復原圖(圖2),表明鳥類的祖先具有利用四肢攀援樹枝和滑翔的能力。Bock(1969,1986)進一步發展了這種假說。他認為鳥類的祖先應是地棲爬行動物,經過長期的自然選擇,進化為兩足行走、可以攀援樹干的地棲原始鳥類。原始鳥類一開始在樹枝間進行短距離跳躍,然后進行稍遠距離的跳躍,從而逐漸獲得了滑翔能力,進而可以從高樹滑翔到矮樹,甚至滑翔到地面,最終具有了遠距離主動飛行的能力。Xu等(2000)報道了產于我國遼西早白堊世義縣組的一種小型奔龍化石——趙氏小盜龍(圖3)。通過研究,認為趙氏小盜龍是一種可以振翅飛行并且棲居樹上的小型恐龍,有力支持了“樹棲起源”假說。

圖2 “樹棲起源”假說生態復原圖(據Heilmann,1926)Fig.2 Ecological restoration plan of the “arborealorigin” hypothesis(after Heilmann,1926)

圖3 趙氏小盜龍生態復原圖(據Xu等, 2000)Fig.3 Ecological restoration plan of the Microraptorzhaoianus(after Xu et al, 2000)
2“地棲起源”假說
“地棲起源”假說亦稱“陸地奔跑起源”假說,最早由Williston(1879)首次提出。他認為兩足行走的恐龍在快速奔跑過程中,由輔助性的捕食、抓握和平衡功能的前肢逐漸拉長,演變為具有羽毛的翅膀,進而獲得了滑翔乃至飛行的能力。Nopsca(1907,1923)撰文支持這種假說,認為鳥類起源于兩足疾走的長尾爬行動物,鳥類的祖先或原始鳥類在快速奔跑時,其前肢是輔助后肢加快奔跑速度的“推動器”。此后,Ostrom(1974,1976,1979)、Padian(1982,1985)等通過對始祖鳥骨骼解剖的研究,進一步完善了“地棲起源”假說,提出“昆蟲網捕”理論,認為腳趾的對握與否是判別鳥類的祖先或鳥類是否具有樹棲能力的重要特征(圖4)。他們認為始祖鳥的后肢非常發育,脛骨長,跗跖骨與股骨的長度比為2∶1,這與奔跑的機理結構相吻合。始祖鳥的第1趾爪(母趾爪)沒有特別拉長,明顯短于其他趾爪,而且其所有趾爪都不強烈彎曲,也不銳利,屈肌突小,這些特征更接近于現生地棲鳥類的趾爪,因而始祖鳥的腳不具有抓握或樹棲功能。Maye等(2005)對新標本的研究也證明始祖鳥的第1趾沒有反轉,不具有抓握或樹棲功能。季強等(2002)報道了1件產自中國遼西早白堊世義縣組的初鳥類化石——中華神州鳥。認為中華神州鳥具有較強的飛行能力,但其第1趾(母趾)沒有反轉,與其他趾的方向相同,表明其具有陸地奔跑能力,而不具有抓握或樹棲功能,有力地支持了“地棲起源”假說。

圖4 “地棲起源”假說中的昆蟲網捕理論示意圖(據Currie,1991)Fig.4 Sketch showing the insect net theory of the“terrestrial origin” hypothesis(after Currie, 1991)
3斜坡起源假說
“斜坡起源”假說系Dial(2003)首次提出,主要依據實驗室中觀察到的一些有趣的現象。將一些尚不具備飛行能力的幼鳥放在不同角度的斜板上反復進行試驗,觀察它們在不同條件下的反應。結果表明,這些幼鳥在30°的斜板上可以輕松地利用后肢從底部跑到頂部;在45°的斜板上仍可以利用后肢從底部跑到頂部;在60°的斜板上利用后肢跑顯得有些吃力,開始扇動翅膀,困難地從底部跑到頂部;在大于70°的斜板上根本就跑不上去。不過,當將一些成鳥放在大于70°的斜板上時,它們就會利用翅膀的扇動輔助后肢從底部“跑”到頂部(圖5)。

圖5 “斜坡起源”假說示意圖(據李志恒等,2008)Fig.5 Sketch showing the “slope origin” hypothesis(after Li et al, 2008)
4問題討論
通過上述介紹可以看出,“樹棲起源”假說特別強調鳥類的祖先或原始鳥類最初必須具有爬樹能力,因為樹木的高度為“滑翔”或“飛行”提供了看似合乎情理的客觀條件,進而認為始祖鳥后肢的母趾與其他3趾相對是適于攀援、抓握的有力證據(實際上最新的研究證明始祖鳥的母趾并沒有反轉,也沒有樹棲能力)。表面看來,“樹棲起源”假說似乎比較合乎情理,但細究之下有許多方面令人難以理解。鳥類的祖先為什么非要爬樹不可?它們爬樹的動機是什么?是為了捕食還是為了躲避敵害?
首先,如果說這種假說的支撐點是產生滑翔所需的“高差”的話,高大直立的樹木是一種可能,但產生滑翔所需的“高差”在自然界中隨處可見,如土堆、陡坎、山崖、斜倒的樹干等均可形成高差。也就是說,高大直立的樹木并不是產生“高差”的唯一可能,特別強調“具有爬樹能力”就顯得太牽強附會了。“爬樹”是眾多可能條件中的一種條件,但絕不是必要的充分條件。
其次,鳥類的祖先是小型食肉性恐龍,食物來源一般是地面生活的小型動物,如哺乳動物、爬行動物以及昆蟲等,它們不可能放棄地面上輕易可得的食物,非要上樹尋找其他食物不可。顯然,為了捕食而上樹看來不能作為支撐這種假說的主要原因。
再者,鳥類的祖先是小型食肉性的恐龍,它們處于食物鏈的頂層,是捕食者,一般利用它們的銳利的牙齒和強壯的后肢去攻擊敵害或捕食小動物。偶爾遇到比它們強大的敵害的情況可能會發生,慌不擇路偶爾爬樹躲避敵害的可能是有的,但絕不是普遍常見的現象。這也表明,鳥類的祖先為了躲避敵害而上樹的理由顯得很不充分。
還有,從骨骼解剖學的角度來看,無論是奔龍還是傷齒龍,它們的后肢長而強壯,明顯長于前肢,除第2趾爪外,其余的趾爪均不強烈彎曲。這些特征表明,它們是善于快速奔跑的捕食者。季強等(2003)專文討論了鳥類飛行的起源,認為鳥類的祖先和原始鳥類在一定階段可能具有一定的(有限的)“爬樹”能力,但并不意味著“爬樹”是獲得飛行能力的“必由之路”,它們可以利用自然界中的各種有利條件(地形、地物)來幫助它們滑翔和起飛。李志恒等(2008)也專文討論了鳥類飛行起源問題,認為鳥類飛行起源的進化途徑只有3種:地面進化、樹上進化和斜坡進化。筆者認為,簡單地把飛行起源進化歸為3種是不科學的,也不符合客觀實際情況,進化途徑應該是多種多樣的。
“地面進化”思想應當來自Williston(1879)提出的“陸地奔跑起源”假說(“地棲起源”假說)。當時僅僅是提出一個簡單的構想和描述,此后該假說在相當長的一段時間內并沒有引起人們太多的關注。美國學者Ostrom(1974,1976,1979)的研究對于該假說的復蘇發揮了非常重要的作用,他詳細研究了鳥類飛行中的關鍵動作“扇翅”的起源,提出了“昆蟲網捕”理論并加以解釋(圖6)。筆者認為“昆蟲網捕”理論成立與否并不重要,始祖鳥是一種陸地快速奔跑的兩足獵食動物,而且不能在樹上棲居,為什么還要將這種理論與“上樹”聯系在一起?

圖6 “地棲起源”假說示意圖(據李志恒等,2008)Fig.6 Sketch of the “terrestrial origin” hypothesis(after Li et al, 2008)
“樹上進化”思想應當來自Marsh(1880)提出的“樹棲起源”假說,此后得到了許多學者的接受和支持。李志恒等(2008)認為,趙氏小盜龍、寧城樹棲龍、顧氏小盜龍等恐龍的發現是支持“樹棲起源”假說的有力證據,它們具有明顯的樹棲特征,如相對較低的第1趾(母趾)位置,較大的趾爪弧度,倒數第2趾加長,相對長的前肢等(圖7)。筆者認為,他們提出的這些特征在奔龍、疾走龍、傷齒龍等中是十分常見的,是快速奔跑的特征,而不是樹棲的特征。以趙氏小盜龍為例:其后肢明顯長于前肢,前肢的3指短小,指爪也不強烈彎曲;其第1趾(母趾)沒有反轉,位置較低,趾爪短且不強烈彎曲;其他各趾均顯得細長,趾爪既不強烈彎曲也不銳利。這些特征均表明趙氏小盜龍更適于陸地奔跑,而不善于爬樹(圖8、圖9、圖10、圖11、圖12)。它的趾爪不具有抓握功能,又如何能在樹上棲居?另一方面,如果趙氏小盜龍的前肢適于爬樹,在攀爬的過程中勢必會不斷強化它的攀爬功能,那么又怎么可能導致它的前肢向鳥類的翅膀演變呢?

圖7 “樹棲起源”假說示意圖(據李志恒等,2008)Fig.7 Sketch of the “arboreal origin” hypothesis(after Li et al, 2008)

圖8 美頜龍類恐龍腳趾形態結構圖Fig.8 Morphological structures of toes of Compsognathus dinosaurs

圖9 奔龍類恐龍腳趾形態結構圖Fig.9 Morphological structures of toes of Dromaeosaur dinosaurs

圖10 傷齒龍類恐龍腳趾形態結構圖Fig.10 Morphological structures of toes of Troodontids dinosaurs
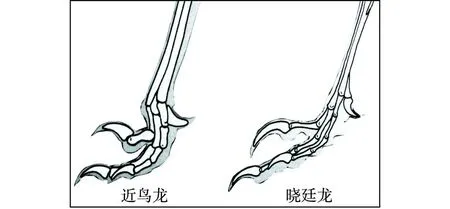
圖11 傷齒龍類恐龍腳趾形態結構圖Fig.11 Morphological structures of toes of Troodontids dinosaurs

圖12 竊蛋龍類恐龍與初鳥類腳趾形態結構圖Fig.12 Morphological structures of toes of Oviraptorid dinosaurs and primitive birds
“斜坡進化”理論實際就是Dial(2003)提出的“斜坡起源”假說。有些人認為對于解釋鳥類飛行起源具有潛在意義,因為具有早成性特征的鳥類祖先可能最初也經歷過這種爬坡過程,而后獲得了飛行能力。筆者不完全茍同于這些學者的看法,將個別特定的自然條件(如上樹、爬坡等)來解釋復雜的飛行起源問題顯得局限、狹隘并帶有濃厚的慣性思維色彩。實驗結果恰恰表明,鳥類的祖先或原始鳥類在獲得飛行能力之前主要是依賴后肢進行奔跑,為了加快奔跑速度或在一定的困難條件下,它們才利用前肢或翅膀扇動產生的推動力來幫助滑翔和起飛,同時也說明鳥類的祖先是兩足奔跑的動物,具體來講,就是小型食肉性恐龍。它們可以利用樹木或斜坡,但不意味著必須依賴樹木或斜坡來獲得滑翔能力和飛行能力。
恐龍或爬行動物畢竟是動物,它們不可能具有人類的思維,更不可能在爬樹之初就預想到幾百萬年后會長出羽毛,演變成鳥類而飛向天空。現在的研究表明,羽毛不是鳥類的專屬特征,許多獸腳類恐龍早在鳥類之前就長出了羽毛。羽毛出現之初的主要功能是保持體溫,是變溫動物向恒溫動物演變的必然結果。隨著自身的發展演化,羽毛后來才逐漸具有了保護身體、運動平衡、性別展示、滑翔、飛行等功能。盡管現生鳥類的羽毛,特別是飛行羽毛,在飛行中發揮了重要作用,但現在還沒有任何證據證明鳥類的祖先或原始鳥類羽毛的出現和早期演化與爬樹有關。任何生物,包括恐龍和鳥類,保護自己的生命是首先必須想到的。恐龍的祖先在沒有發育羽毛或羽毛不夠發育的情況下是不會從高處向下跳的。相反,正是羽毛發育到一定的程度,足以保護自身的生命,它們才有可能從一定的高度向下滑落,而不至于摔傷或摔死。
目前,有關鳥類飛行起源的研究與討論還沒有結束,現有的研究材料還不足以對此問題過早得出結論。也許2種假說都有一定的道理,爭論的焦點在于哪一種假說更合理,更具有代表性和普遍性。筆者認為,“地棲假說”更合乎邏輯,更有道理。鳥類的祖先在一定條件下,完全可以本能地借助自然界中的各種有利地形、地物來滑翔或起飛,如較大的巖石塊體、土堆、斜坡、陡坎、山崖、倒臥的樹干、低矮的灌木等,而不是僅僅依賴于直立高大的樹木。后來有一天,它們突然發現自己展開雙翅后,能夠從高處向低處滑翔,繼而進行短距離飛行,最終能夠自由翱翔于藍天與大地之間。為此,筆者給出了一個全新的“地棲起源”假說示意圖(圖13)。

圖13 新的“地棲起源”假說示意圖Fig.13 Sketch showing the new “terrestrial origin”hypothesis
國際上早就在討論鳥類起源、鳥類的飛行起源、四翼恐龍、羽毛的起源與演化等諸多問題,有些是肯定的意見,也有許多不同的聲音,但國內知之甚少,究其原因主要是一些專家壟斷了資源、信息和新聞,各種不同或反對的意見難以在國內見諸于世。若這種情況再繼續下去,則會影響今后研究工作的開展,不利于我國科學技術的發展與進步。
2014年年末,《Science》評選出了2014年度“國際十大科技進展”,鳥類起源與鳥類飛行起源研究排名第二,文中的最后一句話十分耐人尋味:“盡管鳥類飛向了藍天,但它們的飛行源自它們恐龍祖先的陸地奔跑(Birds took off, but their dinosaur ancestors had given them a running start.)”。
參考文獻:
季強,姬書安. 1996.中國最早鳥類化石的發現及鳥類的起源. 中國地質,(10):30-33.
季強,姬書安.1997.原始祖鳥(Protarchaeopteryxgen. nov.):中國的始祖鳥類化石. 中國地質,(3):38-41.
季強,姬書安,尤海魯,等.2002.中國首次發現真正會飛的“恐龍”:中華神州鳥(新屬新種).地質通報,21(7):363-369.
季強,袁崇喜,季鑫鑫,等.2003.論鳥類飛行的起源. 地質論評,49(1):1-3.
李志恒,張玉光,周忠和.2008.鳥類飛行起源的研究.自然雜志,30(1):32-38.
鄭光美.1995.鳥類學. 北京:北京師范大學出版社.
BOCK W J. 1969. The origin and radiation of birds. ANN N Y acad Sci, 167: 147-155.
BOCK W J. 1986. The arboreal origin of avian flight // PADIAN K. The origin of birds and the evolution of flight. San Francisco: Calif. Acad Sci,57-72.
BURGERS P, CHIAPPE L M.1999. The wing ofArchaeopteryxas a primary thrust generator . Nature, 399: 60-62.
CHATTERJEE S. 1997. The rise of birds . Baltimore, MD,USA:Johns Hopkins Univ Press.
CHIAPPE L M. 1997.ClimbingArchaeopteryx? A response to Yalden . Archaeopteryx, 15: 109-112.
DIAL K D. 2003.Wing-assisted incline running and the evolution of flight . Science, 299: 402-404.
FEDUCCIA A. 1993. Evidence from claw geometry indicatingarboreal habits ofArchaeopteryx. Science,259: 790-793.
FARLOW J O.2000.Theropod locomotion . American Zoologist,40: 640-663.
HEILMANN G. 1926. Origin of Birds .London,UK: Witherby.
JI QIANG.1998. Two feathered dinosaurs from northeasternChina . Nature, 393: 753-761.
JI QIANG. 2001. The distribution of integumentary structures in a feathered dinosaur . Nature, 410: 1084-1088.
LONGRICH N. 2006. Structure and function of hindlimb feathers inArchaeopteryxlithographica. Paleobiology,32 (3): 417-431.
MARSH O C. 1880. Odontornithes:A Monograph on the Extinct Toothed Birds of North America; With Thirty-four Plates and Forty Woodcuts. Washington, USA:Govt print off.
MAYE G, POHL B, PETERS D S A. 2005. A well-preservedArchaeopteryxspecimen with theropod features . Science, 310: 1483-1486.
NOPSCA F. 1907. Ideas on the origin of flight . Proc Zool Soc Lond,15: 223-236.
NOPSCA F. 1923. On the origin of flight in birds . Proc Zool Soc Lond, 31:463-477.
NORELL M A.2002.“Modern” feathers on a non-avian dinosaur. Nature, 416: 36-37.
OSTROM J.1974.Archaeopteryxand the origin of flight . Quart Rev Biol,49: 27-47.
OSTROM J.1976.Archaeopteryxand the origin of birds . Bio J Linnean Soc,8: 91-182.
OSTROM J. 1979. Bird flight: how did it begin? . American Scientist, 67: 46-56.
PADIAN K.1982.Running, Leaping, lifting off . The Sciences,10-15.
PADIAN K. 1985. The origin and aerodynamics of flight in extinct vertebrates . Palaeontology,28 (3): 413-433.
PADIAN K. 2003. Four winged dinosaurs, bird precursors, or neither? . Bio Science,53 (5): 450-452.
PADIAN K, DIAL K P. 2005. Origin of flight: could four-winged dinosaurs fly? . Nature,438: E3-E4.
WILLISTON S. 1879. Are birds derived from dinosaurs? .Kansas City Rev Sci, 3: 457-460.
XU XING, ZHOU ZHONGHE, WANG XIAOLIN. 2000. The smallest known non-avian theropod dinosaur . Nature, 408: 705-708.
YALDEN D W. 1997. ClimbingArchaeopteryx. Archaeopteryx, 15: 107-108.
A debate between the “arboreal” and “cursorial” hypotheses of the origin of bird flight
JI Yan-nan1, WANG Xu-ri2, Kurosu Mariko3, JI Qiang2
(1. China Geological Environmental Monitoring Institute, Beijing 100081, China; 2. Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China; 3. China University of Geosciences (Beijing), Beijing 100083, China)
Abstract:This paper briefly introduces two hypotheses of the origin of bird flight: the “arboreal” hypothesis and the “cursorial” (running)hypothesis, and discusses their theoretical bases and some problems in detail. It is considered that the ancestors and primitive birds could get their gliding ability and flight power with the help of favorable natural terrains, such as hill, hillock, slope, brae, cliff, steep ridge, huge boulder and inclined tree trunk, rather than only by the aid of erect tree trunk. It is suggested that the “running” hypothesis seems more reasonable, more close to, or, more coincident with the objective facts. Herein is given a new theoretical model of bird flight.
Keywords:bird flight; arboreal hypothesis; cursorial hypothesis; wing-assisted incline running hypothesis
