順鉑誘發大鼠睪丸毒性的機制及葡萄籽原花青素的防護作用
2015-11-02 13:00:53連燕娜張海蓮趙江燕高麗萍
食品科學 2015年15期
連燕娜,張海蓮,趙江燕,高麗萍
(北京聯合大學 生物活性物質與功能食品北京市重點實驗室,北京 100191)
順鉑誘發大鼠睪丸毒性的機制及葡萄籽原花青素的防護作用
連燕娜,張海蓮,趙江燕,高麗萍*
(北京聯合大學 生物活性物質與功能食品北京市重點實驗室,北京 100191)
目的:探究葡萄籽原花青素(grape seed proanthocyanidin extracts,GSPE)對順鉑(cisdiamminedichloroplatinum,CDDP)誘發大鼠睪丸氧化損傷和細胞凋亡的防護作用。方法:將40 只大鼠隨機分為空白對照組、CDDP模型組、GSPE低劑量組(200 mg/(kg·d))、GSPE高劑量組(400 mg/(kg·d))。分別以蒸餾水和相應劑量的GSPE對大鼠連續灌胃15 d,灌胃10 d后一次性腹腔注射CDDP,劑量為7 mg/kg,空白對照組注射相同劑量的生理鹽水。注射CDDP后第5天處死大鼠,檢測精子指標、大鼠睪丸組織中超氧化物歧化酶(superoxide dismutase,SOD)與谷胱甘肽過氧化物酶(glutathione peroxidase,GSH-Px)活力、還原型谷胱甘肽(glutathione,GSH)與丙二醛(malondialdehyde,MDA)含量,流式細胞儀檢測大鼠睪丸細胞凋亡率,并觀察睪丸組織在光鏡下形態的變化。結果:CDDP模型組大鼠睪丸和附睪的質量、附睪精子的濃度和活動度、GSH-Px、SOD活力以及GSH含量均顯著降低,而MDA水平和睪丸細胞凋亡率顯著升高。預防性口服GSPE顯著改善了CDDP引起的大鼠睪丸質量減小、功能減弱以及氧化應激、脂質過氧化和細胞凋亡這些不利影響。結論:GSPE對CDDP誘發的大鼠睪丸毒性有劑量依賴性防護作用,其機制很可能與抑制體內氧化應激及細胞凋亡有密切關系。
順鉑;葡萄籽原花青素;睪丸毒性;氧化應激;細胞凋亡
順鉑(cis-diamminedichloroplatinum,CDDP)是一種有效的抗腫瘤DNA烷化劑,可以治療多種實體腫瘤,如膀胱癌、卵巢癌、宮頸癌、子宮癌、腦癌、頸癌和肺癌等[1-2]。同時,CDDP在睪丸癌的治療中也廣泛應用,且治愈率>90%[3]。但CDDP對生殖系統產生的副作用限制了其治療效果[4-5]。CDDP誘發睪丸損傷的機理還不清楚,但許多研究[6-9]已經證明,CDDP的副作用與產生自由基和活性氧(reactive oxygen species,ROS),從而引發氧化應激作用有關。ROS通常都是在睪丸亞細胞結構中產生,尤其是線粒體,這些ROS緊接著會被相應細胞器的抗氧化系統清除。然而,CDDP會破壞抗氧化系統的平衡,導致細胞功能紊亂[7-9]。當活性氧大量產生時,會激發DNA斷裂和精子功能喪失,這與線粒體和精細胞膜的過氧化損傷有關。而且,由于精細胞多不飽和脂肪酸含量高并且抗氧化能力低,因此對過氧化損傷更敏感[10]。
鑒于ROS和氧化應激在CDDP誘發毒性途徑中的重要作用,在大多數抗CDDP毒性的研究中有一種方法為使用天然或人工合成的抗氧化劑。目前所報道的具有這種保護作用的一些天然化合物[11-19]有VC、VE、VA,白藜蘆醇、槲皮素、柚皮素和番茄紅素,還有一些人工合成的化合物,如卡維地洛、依達拉奉等。這些抗氧化劑對CDDP毒性的防護主要是通過其自由基清除作用實現的。葡萄籽原花青素(grape seed proanthocyanidin extracts,GSPE)是具有多種藥理活性的黃酮類化合物。已有報道稱由于原花青素有抗氧化、抗炎、清除自由基、保護腎臟以及抗癌的作用,使其具有治療病痛的潛能[20-22]。由于其具有很強的抗氧化活性、低毒性以及無遺傳毒性,GSPE在美國市場作為一種膳食補充劑被推廣[23]。本實驗旨在檢驗大鼠經CDDP給藥后造成的生殖功能損傷狀況,并探究GSPE對該損傷可能的保護作用。
1 材料與方法
1.1動物、材料與試劑
成年健康雄性Sprague-Dawley(SD)大鼠40 只(SPF級),體質量140~160 g,由中國人民解放軍軍事科學院實驗動物中心提供。
順鉑(CDDP,批號001001DF),注射用凍干型粉劑,每支10 mg,山東齊魯制藥廠生產,用時以生理鹽水配制;葡萄籽原花青素(GSPE,純度>95%),購于天津市尖峰天然產物研究開發有限公司,用時以雙蒸水配制。
考馬斯亮藍蛋白試劑盒、還原型谷胱甘肽(glutathione,GSH)試劑盒、谷胱甘肽過氧化物酶(glutathione peroxidase,GSH-Px)試劑盒、超氧化物歧化酶(superoxide dismutase,SOD)試劑盒、丙二醛(malondialdehyde,MDA)試劑盒 南京建成生物工程研究所;碘化丙啶(propidium iodide,PI) 上海佳和生物科技有限公司。
1.2儀器與設備
紫外-可見分光光度計 尤尼柯上海儀器有限公司;石蠟包埋機、半自動轉輪式切片機 德國Laica公司;三目攝像生物顯微鏡系 日本Olympus公司;流式細胞儀 美國Beckman Coulter公司。
1.3方法
1.3.1動物分組與模型建立
雄性SD大鼠在標準實驗條件下飼養,保持12 h光照/12 h黑暗,溫度(24±3) ℃。用普通鼠糧適應性飼養1 周后,將40 只大鼠隨機分為4 組,每組10 只。實驗第1天起,空白對照組、CDDP模型組每天灌胃蒸餾水(0.5 mL/100 g,以體質量計,下同),GSPE低、高劑量組分別以200、400 mg/(kg·d)的劑量連續灌胃GSPE(0.5 mL/100 g)15 d。本實驗中使用的GSPE的灌胃劑量基于前期的研究報道[24-25]。此外,在實驗的第10天CDDP模型組和GSPE低、高劑量組大鼠一次性腹腔注射CDDP 7 mg/kg,這是已知的能夠引起大鼠睪丸毒性的劑量[26]。空白對照組注射等量的生理鹽水。
1.3.2樣本采集
所有大鼠在注射CDDP后第5天通過股動脈放血法處死,在冰浴條件下迅速取出兩側睪丸及附睪,去除黏連的結締組織并稱質量。1 個睪丸固定在體積分數10%福爾馬林中用于組織切片,另1 個睪丸沿橫切面一分為二,一半放入液氮中迅速冷凍后轉移至-80 ℃條件下保存用于生化分析,另一半取一部分組織浸泡于70%的乙醇中,用于凋亡的檢測,剩余部分稱量質量后按1 mL/mg加入DMEM培養基勻漿,用于精子計數。
將睪丸組織從深度冷凍狀態取出并稱質量,轉移到預冷的玻璃管中,用9 倍體積(V/m)的磷酸鹽緩沖液(pH 7.4)稀釋。進行酶活性分析時,將睪丸組織切碎并用冷生理鹽水作為勻漿介質,冰浴條件下在聚四氟乙烯玻璃勻漿器中10 000 r/min勻漿3 min。制備好的組織勻漿4 ℃、825×g離心15 min,取上清液于-20 ℃條件下凍存,用于后續的生化指標檢測。
1.3.3指標檢測方法
1.3.3.1精子濃度和活動度
精子計數采用Yokoi等[27]的一種改良方法。
評估精子活動度的方法:取1~2 滴上述附睪精子懸液滴于清潔載玻片上,室溫((20±2)℃)條件下,于20 min內在400×光鏡下進行觀察,每只大鼠分析100 個精子,并對精子的活動度進行分級。
1.3.3.2生化指標測定
取凍存于-20 ℃的組織勻漿上清液,按照試劑盒說明書方法,采用硫代巴比妥酸法測定勻漿上清液中MDA含量;二硫代二硝基苯甲酸法測定GSH含量;黃嘌呤氧化酶法測定SOD活性;二硫代二硝基苯甲酸法測定GSH-Px活性;考馬斯亮藍法測定蛋白質含量。
1.3.3.3細胞凋亡的測定
將70%乙醇固定的組織放在120 目不銹鋼網上,下置一平皿,用眼科剪將組織剪碎,用鑷子輕輕揉搓組織塊,并用生理鹽水沖洗,直至將組織搓碎為止。去除平皿中混懸液的細胞團塊,收集細胞懸液,500~800 r/min離心2 min。取上述細胞懸液,調整其濃度為1×106個/mL,取0.1 mL該溶液(加入10%雞紅細胞作為內參標準,與樣品同步染色),加入PI染液1 mL,在4 ℃冰箱中避光染色30 min,以500 目銅網過濾,使樣品成為合格的單細胞懸液。流式細胞儀激發波長為488 nm,檢測前以flowcheckTMFluorpheres(10 μm)熒光微球作為標準樣品,調整儀器變異系數在2%以內。
1.3.3.4組織病理學檢測
睪丸樣本取出后放入10%的中性緩沖福爾馬林中進行固定,經常規處理和蘇木精-伊紅(hematoxylineosin,HE)染色后,用光學顯微鏡進行檢查。運用顯微測微器在光學顯微鏡下測每個睪丸中5 個不同位置的曲細精管的直徑和生發層細胞厚度,計算曲細精管的平均直徑和生發層細胞的平均厚度。
1.4數據統計分析
用SPSS 12.0統計軟件進行統計學處理,實驗數據采用±s表示,顯著水平設定為P<0.05,多個樣本均數的比較采用單因素方差分析,Duncan's多重比較法用來確定組間差異的顯著性。
2 結果與分析
2.1GSPE對大鼠睪丸及附睪質量的影響
表1 各組大鼠睪丸及附睪質量Table 1 Testes and epididymis weights of rats in all groups

表1 各組大鼠睪丸及附睪質量Table 1 Testes and epididymis weights of rats in all groups
注:a.與空白對照組相比,差異極顯著(P<0.01);b.與CDDP模型組相比,差異顯著(P<0.05);c.與CDDP模型組相比,差異極顯著(P<0.01)。
組別睪丸質量/g附睪質量/g空白對照組2.533 1±0.119 50.536 2±0.064 8 CDDP模型組2.151 5±0.226 7a0.360 0±0.051 7aGSPE低劑量組2.297 7±0.091 3a0.422 5±0.080 8abGSPE高劑量組2.411 5±0.102 2c0.488 5±0.047 8c
由表1可知,CDDP模型組大鼠的睪丸、附睪質量與空白對照組相比極顯著降低(P<0.01)。與CDDP模型組相比,GSPE高劑量組大鼠的睪丸質量和附睪質量極顯著增加(P<0.01),GSPE低劑量組大鼠的附睪質量顯著增加(P<0.05)。以上結果提示預防性給藥GSPE對CDDP引起的生殖損傷有減緩效果。然而與CDDP模型組相比,GSPE低劑量組大鼠睪丸質量雖有所增加,但無顯著差異(P>0.05)。
2.2GSPE對大鼠附睪精子濃度、活動度、畸形率及睪丸精子濃度的影響
表2 各組大鼠的附睪精子濃度、活動度、畸形率和睪丸精子濃度Table 2 Epididymal sperm concentration, sperm motility, abnormal sperm rate and testicular sperm concentration in rats from all groups

表2 各組大鼠的附睪精子濃度、活動度、畸形率和睪丸精子濃度Table 2 Epididymal sperm concentration, sperm motility, abnormal sperm rate and testicular sperm concentration in rats from all groups
注:a. 與空白對照組相比,差異顯著(P<0.05);b. 與空白對照組相比,差異極顯著(P<0.01);c. 與CDDP模型組相比,差異顯著(P<0.05);d. 與CDDP模型組相比,差異極顯著(P<0.01)。下同。
睪丸精子濃度/(105個/mL)空白對照組17.23±7.4770.12±7.202.14±0.8218.10±3.94 CDDP模型組8.67±4.16b49.06±14.40b3.56±0.44b9.64±2.39bGSPE低劑量組9.04±5.35b58.48±7.60bc2.96±0.43ad13.38±3.39aGSPE高劑量組15.46±3.99d63.45±7.60d2.69±0.41d17.33±3.46d組別附睪精子濃度/(105個/mL)附睪精子活動度/%附睪精子畸形率/%
由表2可知,與空白對照組相比,CDDP模型組大鼠的精子濃度和附睪精子活動度極顯著降低(P<0.01),附睪精子畸形率極顯著升高(P<0.01)。與CDDP模型組相比,GSPE高劑量組大鼠的附睪精子濃度、附睪精子活動度極顯著增加(P<0.01),GSPE低劑量組大鼠的附睪精子活動度顯著增加(P<0.05),GSPE低、高劑量組大鼠的附睪精子畸形率極顯著降低(P<0.01),以上結果表明GSPE可有效抑制CDDP誘導的大鼠精子質量方面的副作用,包括附睪精子濃度、活動度和畸形率。
2.3GSPE對大鼠睪丸組織中MDA和GSH含量、GSH-Px和SOD活性的影響
表3 各組大鼠睪丸組織中MDA、GSH含量及SOD、GSH-Px活性Table 3 MDA and GSH levels and SOD and GSH-Px activities in testes tissue in rates from all groups

表3 各組大鼠睪丸組織中MDA、GSH含量及SOD、GSH-Px活性Table 3 MDA and GSH levels and SOD and GSH-Px activities in testes tissue in rates from all groups
GSH-Px活力/(U/g pro)空白對照組2.04±0.54156.64±8.59520.80±84.23642.10±28.07 CDDP模型組3.52±1.04b126.60±11.31b401.97±79.24b544.07±63.00bGSPE低劑量組2.97±0.69a153.61±7.38d501.93±73.59c624.55±57.56dGSPE高劑量組2.54±0.78d166.30±13.82d527.55±37.48d637.21±59.83c組別MDA含量/(nmol/g pro)SOD活力/(U/g pro)GSH含量/(?mol/g pro)
由表3可知,與空白對照組相比,CDDP模型組大鼠的睪丸組織中MDA含量極顯著升高(P<0.01),而GSH含量、GSH-Px和SOD活力極顯著降低(P<0.01)。與CDDP模型組相比,GSPE高劑量組大鼠的睪丸組織中MDA含量極顯著降低(P<0.01),GSPE低劑量組大鼠的睪丸組織中MDA含量降低,但無顯著差異(P>0.05)。此外,與CDDP模型組相比,GSPE低劑量組大鼠睪丸組織中的GSH-Px、SOD活力及GSH含量增加(P<0.05或P<0.01)。
2.4GSPE對大鼠睪丸細胞凋亡的影響

圖1 各組大鼠睪丸細胞凋亡情況Fig.1 Testicular cell apoptosis of rats in all groups
由圖1可知,與空白對照組大鼠睪丸細胞凋亡率((1.02±0.09)%)相比,CDDP模型組大鼠的睪丸細胞凋亡率((3.15±0.68)%)顯著升高(P<0.01)。與CDDP模型組相比,GSPE高劑量組大鼠睪丸細胞凋亡率((2.57±0.43)%)顯著降低(P<0.05),GSPE低劑量組大鼠睪丸細胞凋亡率((2.88±0.55)%)降低,但差異不顯著(P>0.05)。
2.5各組大鼠睪丸組織病理學的變化
由表4可知,與空白對照組相比,CDDP模型組大鼠的睪丸曲細精管直徑和生發層細胞厚度極顯著降低(P<0.01),GSPE低劑量組未改善這種損害,然而給藥高劑量的GSPE顯著或極顯著地緩解了這種損害(P<0.05或P<0.01)。如圖2所示,空白對照組大鼠睪丸曲細精管排列規則,管腔內有大量成熟精子。CDDP模型組大鼠的睪丸曲細精管排列不規則、生精上皮細胞層次減少、管腔內成熟精子減少、且有空泡形成。GSPE低劑量組大鼠與空白對照組相比,雖然出現了曲細精管排列較不規則、管腔內成熟精子較少的現象,但與CDDP模型組相比已有改善。GSPE高劑量組與空白對照組相比,大鼠睪丸曲細精管排列基本規則,各級生精細胞未見明顯改變,較CDDP模型組明顯改善。
表4 各組大鼠曲細精管直徑及生發層細胞的厚度Table 4 Diameter of seminiferous tubules and germinal layer thickness of rats in all groups
組別曲細精管直徑/μm生發層細胞厚度/μm空白對照組318.52±30.84106.69±12.02 CDDP模型組279.07±32.48b80.43±8.76bGSPE低劑量組281.90±16.07a89.36±13.67bGSPE高劑量組310.52±36.89c94.54±12.99ad
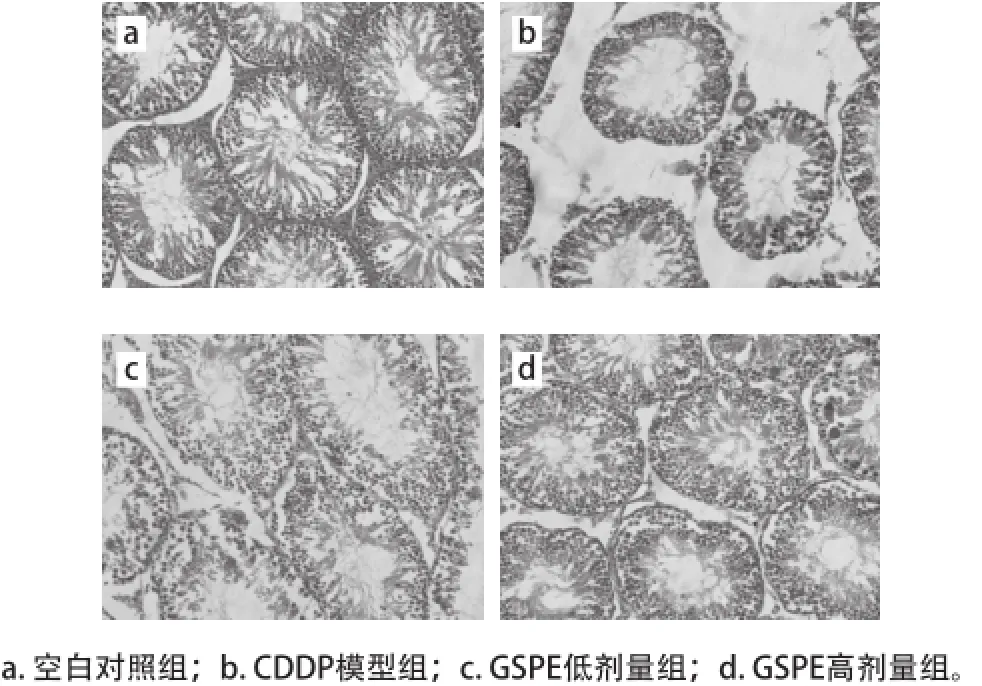
圖2 各組大鼠睪丸組織病理變化(HE,×40000)Fig.2 Pathological changes of testis tissue in rats (HE, ×400)
3 討 論
睪丸惡性腫瘤是常見的腫瘤病之一,腫瘤化療藥物在很多情況下已經提高了腫瘤患者的生存率。但是這種治療導致副作用的發病率比較高,睪丸功能障礙就是這種治療最常見的長期副作用之一[28]。近年來,化療后睪丸功能損害已經引起了研究人員的廣泛關注。用于化療的許多藥物尤其是烷化劑都具有生殖腺毒性作用,其生殖毒性與抗腫瘤藥物基團、所用化療藥物的劑量、處理時間及個體敏感度的變化有關[28-29]。CDDP是一種用于治療睪丸癌、卵巢癌、頭頸部癌癥等有效的化療烷化劑。有研究者報道CDDP的使用引起了暫時性或永久性精子活力缺乏或精子減少[30-31]。本實驗的結果表明,7 mg/kg CDDP的使用能造成大鼠精子濃度、精子活動度顯著降低及精子畸形率顯著增加。
生發層上皮細胞損傷而造成的精子減少或精子活力缺乏,一直被公認為是化療藥物治療的后果之一[4,32]。化療誘導的睪丸損傷與特定的藥物種類及劑量有關[28]。本實驗結果顯示CDDP誘發了大鼠睪丸與附睪質量減輕。組織病理學切片也表明CDDP引起了睪丸曲細精管直徑和生發層細胞厚度的減少。以上結果,尤其是精子特性和組織病理學方面的損傷與一些學者的研究報道一致[7-9],這提示CDDP可誘發睪丸損傷,從而造成了精子減少或精子活力缺乏,對大鼠產生了明顯的生殖毒性。
CDDP誘發大鼠生殖毒性的分子機理尚不清楚。一些體內與體外研究[6-7,9]表明實驗動物暴露于CDDP會伴隨氧化應激的產生。氧化應激是一種與ROS的產生和消除有關的現象。根據藥物毒性的分子生物學機制,ROS和氧化性損傷可能通過減弱精子的功能而引起雄性不育。已經發現在低生育或不育男性的精漿和精子中有過量的ROS和自由基產生[33]。
本實驗中,與空白對照組相比,注射CDDP顯著地增加了大鼠睪丸組織中MDA的含量,減少了GSH含量和GSH-PX、SOD活性。這些指標上的變化表明抗氧化酶的活性不足以平衡注射7 mg/kg CDDP后在指定時間內產生的自由基。此外,MDA含量增加也證實了這個結論。SOD活性的減弱導致超氧陰離子自由基含量的增加。另外,這些抗氧化酶活性的減小表明在自由基轉化成損傷更小或者無損傷的代謝物的過程中這些酶的消耗和反應,其次也表明CDDP對酶活性可能有直接抑制或刺激作用。Türk等[9]研究結果顯示CDDP在精液、精子和睪丸組織中引起MDA含量增加。此外,Salem等[7]也報道了CDDP引起Wistar大鼠睪丸組織中MDA含量顯著增加,同時抗氧化酶活性的降低易增加精子的自由基損傷。
有研究[34-35]表明,高劑量CDDP作用下細胞DNA損傷嚴重以致不能修復,促使細胞發生凋亡或死亡。本研究通過DNA含量的降低粗略定量凋亡細胞百分比,不能區分早期、晚期及壞死細胞,可作為凋亡細胞的初步篩選。本研究結果顯示,腹腔注射CDDP顯著增加了睪丸細胞凋亡率,證明了CDDP可促進細胞凋亡或死亡的發生。
GSPE包括低聚原花青素,是有生物活性的黃酮化合物,已被廣泛應用于生物、化療等領域。本實驗發現400 mg/kg劑量的GSPE能顯著的改善CDDP所引起的大鼠精子質量下降。用GSPE預處理能夠顯著的抑制大鼠睪丸組織中由CDDP引起的MDA和GSH的耗竭,并增加SOD和GSH-Px活性。用高劑量的GSPE預處理能夠比低劑量的GSPE更顯著地改善睪丸功能。GSPE抑制CDDP誘導過氧化脂質增加的原因可能是由于它能夠與氧代謝物反應。實驗結果發現CDDP引起大鼠精子濃度和活性顯著降低且精子畸形率顯著增加,但GSPE的使用能夠顯著改善這些狀況,尤其是高劑量的GSPE。由以上結果推測GSPE拮抗CDDP誘導的異常精子比率的保護作用是由于其具有抗氧化特性。以上結果或許可以說明GSPE對CDDP誘導的氧化應激有一定的緩解作用。
總之,本實驗結果表明,GSPE對CDDP引起的睪丸毒性有防護作用。GSPE的這一防護作用可能與氧化應激的抑制有密切關系。因此在化學療法中可以考慮將GSPE和CDDP同時使用進而改善CDDP誘導的精子質量下降和氧化應激損傷。
[1] COLPI G M, CONTALBI G F, NERVA F, et al. Testicular function following chemo-radio therapy[J]. European Journal of Obstetrics & Gynecology and Reproductive Biology, 2004, 113(Suppl 1): 2-6.
[2] HOWELL S J, SHALET S M. Spermatogenesis after cancer treatment:damage and recovery[J]. Journal of the National Cancer Institute. Monographs, 2005, 34: 12-17.
[3] WANG D, LIPPARD S J. Cellular processing of platinum anticancer drugs[J]. Nature Reviews Drug Discovery, 2005, 4(4): 307-320.
[4] CHERRY S M, HUNT P A, HASSOLD T J. Cisplatin disrupts mammalian spermatogenesis, but does not affect recombination or chromosome segregation[J]. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 2004, 564(2): 115-128.
[5] ISHIKAWA T, KAMIDONO S, FUJISAWA M. Fertility after high-dose chemotherapy for testicular cancer[J]. Urology, 2004,63(1): 137-140.
[6] AHMED E A, OMAR H M, RAGB S M M, et al. The antioxidant activity of vitamin C, DPPD and L-cysteine against cisplatin-induced testicular oxidative damage in rats[J]. Food and Chemical Toxicology,2011, 49(5): 1115-1121.
[7] SALEM E A, SALEM N A, MAAROUF A M, et al. Selenium and lycopene attenuate cisplatin-induced testicular toxicity associated with oxidative stress in wistar rats[J]. Urology, 2012, 79(5): 1184.e1-1184.e6. doi: 10.1016/j.urology.2011.12.006.
[8] ILBEY Y O, OZBEK E, SIMSEK A, et al. Potential chemoprotective effect of melatonin in cyclophosphamide-and cisplatin-induced testicular damage in rats[J]. Fertility and Sterility,2009, 92(3): 1124-1132.
[9] T?RK G, ATE??AHIN A, S?NMEZ M, et al. Improvement of cisplatin-induced injuries to sperm quality, the oxidant-antioxidant system, and the histologic structure of the rat testis by ellagic acid[J]. Fertility and Sterility, 2008, 89(5): 1474-1481.
[10] VERNET P, AITKEN R J, DREVET J R. Antioxidant strategies in the epididymis[J]. Molecular and Cellular Endocrinology, 2004, 216(1): 31-39.
[11] TARLADACALISIR Y T, KANTER M, UYGUN M. Protective effects of vitamin C on cisplatin-induced renal damage: a light and electron microscopic study[J]. Renal Failure, 2008, 30(1): 1-8.
[12] AJITH T A, ABHISHEK G, ROSHNY D, et al. Co-supplementation of single and multi doses of vitamins C and E ameliorates cisplatininduced acute renal failure in mice[J]. Experimental and Toxicologic Pathology, 2009, 61(6): 565-571.
[13] DILLIOGLUGIL M O, MARAL KIR H, GULKAC M D, et al. Protective effects of increasing vitamin E and A doses on cisplatininduced oxidative damage to kidney tissue in rats[J]. Urologia Internationalis, 2005, 75(4): 340-344.
[14] do AMARAL C L, FRANCESCATO H D C, COIMBRA T M, et al. Resveratrol attenuates cisplatin-induced nephrotoxicity in rats[J]. Archives of Toxicology, 2008, 82(6): 363-370.
[15] FRANCESCATO H D C, COIMBRA T M, COSTA R S, et al. Protective effect of quercetin on the evolution of cisplatin-induced acute tubular necrosis[J]. Kidney and Blood Pressure Research, 2004,27(3): 148-158.
[16] BADARY O A, ABDEL-MAKSOUD S, AHMED W A, et al. Naringenin attenuates cisplatin nephrotoxicity in rats[J]. Life Sciences,2005, 76(18): 2125-2135.
[17] ATESSAHIN A, YILMAZ S, KARAHAN I, et al. Effects of lycopene against cisplatin-induced nephrotoxicity and oxidative stress in rats[J]. Toxicology, 2005, 212(2): 116-123.
[18] RODRIGUES C, RODRIGUES J L, MARTINS N M, et al. Carvedilol protects against the renal mitochondrial toxicity induced by cisplatin in rats[J]. Mitochondrion, 2010, 10(1): 46-53.
[19] IGUCHI T, NISHIKAWA M, CHANG B J, et al. Edaravone inhibits acute renal injury and cyst formation in cisplatin-treated rat kidney[J]. Free Radical Research, 2004, 38(4): 333-341.
[20] HOUDE V, GRENIER D, CHANDAD F. Protective effects of grape seed proanthocyanidins against oxidative stress induced by lipopolysaccharides of periodontopathogens[J]. Journal of Periodontology, 2006, 77(8): 1371-1379.
[21] SHAO Zuohui, BECKER L B, VANDEN HOEK T L, et al. Grape seed proanthocyanidin extract attenuates oxidant injury in cardiomyocytes[J]. Pharmacological Research, 2003, 47(6): 463-469.
[22] VAYALIL P K, MITTAL A, KATIYAR S K. Proanthocyanidins from grape seeds inhibit expression of matrix metalloproteinases in human prostate carcinoma cells, which is associated with the inhibition of activation of MAPK and NFκB[J]. Carcinogenesis,2004, 25(6): 987-995.
[23] RAY S, BAGCHI D, LIM P M, et al. Acute and long-term safety evaluation of a novel IH636 grape seed proanthocyanidin extract[J]. Research Communications in Molecular Pathology and Pharmacology,2000, 109(3/4): 165-197.
[24] YOUSEF M I, SAAD A A, EL-SHENNAWY L K. Protective effect of grape seed proanthocyanidin extract against oxidative stress induced by cisplatin in rats[J]. Food and Chemical Toxicology, 2009, 47(6):1176-1183.
[25] SAAD A A, YOUSSEF M I, EL-SHENNAWY L K. Cisplatin induced damage in kidney genomic DNA and nephrotoxicity in male rats: the protective effect of grape seed proanthocyanidin extract[J]. Food and Chemical Toxicology, 2009, 47(7): 1499-1506.
[26] SILVA C ? R, GREGGI ANTUNES L M, BIANCHI M L P. Antioxidant action of bixin against cisplatin-induced chromosome aberrations and lipid peroxidation in rats[J]. Pharmacological Research, 2001, 43(6): 561-566.
[27] YOKOI K, UTHUS E O, NIELSEN F H. Nickel deficiency diminishes sperm quantity and movement in rats[J]. Biological Trace Element Research, 2003, 93(1/3): 141-153.
[28] HOWELL S J, SHALET S M. Testicular function following chemotherapy[J]. Human Reproduction Update, 2001, 7(4): 363-369.
[29] MARTIN R H, ERNST S, RADEMAKER A, et al. Analysis of sperm chromosome complements before, during, and after chemotherapy[J]. Cancer Genetics and Cytogenetics, 1999, 108(2): 133-136.
[30] CHERRY S M, HUNT P A, HASSOLD T J. Cisplatin disrupts mammalian spermatogenesis, but does not affect recombination or chromosome segregation[J]. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 2004, 564(2): 115-128.
[31] COLPI G M, CONTALBI G F, NERVA F, et al. Testicular function following chemo-radiotherapy[J]. European Journal of Obstetrics & Gynecology and Reproductive Biology, 2004, 113(Suppl 1): 2-6.
[32] ATE??AHIN A, KARAHAN ?, T?RK G, et al. Protective role of lycopene on cisplatin-induced changes in sperm characteristics,testicular damage and oxidative stress in rats[J]. Reproductive Toxicology, 2006, 21(1): 42-47.
[33] BAKER M A, AITKEN R J. Reactive oxygen species in spermatozoa:methods for monitoring and significance for the origins of genetic disease and infertility[J]. Reproductive Biology and Endocrinology,2005, 3(1): 67. doi: 10.1186/1477-7827-3-67.
[34] 劉玲, 張娟娟, 何蕊玲, 等. 黃芪注射液對順鉑所致小鼠腎損害的保護作用[J]. 中國中藥雜志, 2010, 35(20): 2736-2740.
[35] RYU H G, JEONG S J, KWON H Y, et al. Penta-O-galloyl-β-D-glucose attenuates cisplatin-induced nephrotoxicity via reactive oxygen species reduction in renal epithelial cells and enhances antitumor activity in Caki-2 renal cancer cells[J]. Toxicology in Vitro, 2012, 26:206-214.
Mechanism of cis-Diamminedichloroplatinum-Induced Testicular Toxicity and Protective Effect of Grape Seed Proanthocyanidin Extract in Rats
LIAN Yanna, ZHANG Hailian, ZHAO Jiangyan, GAO Liping*
(Beijing Municipal Key Laboratory of Biological Active Substance and Functional Food, Beijing Union University, Beijing 100191, China)
Objective: To investigate the possible protective role of grape seed proanthocyanidin extract (GSPE) on cisdiamminedichloroplatinum (CDDP)-induced spermiotoxicity. Methods: A total of 40 rats were randomly divided into 4 groups: blank control group, CDDP model group, and low-dose GSPE + CDDP group, high-dose GSPE + CDDP group(orally administered at doses of 200 mg/(kg·d) and 400 mg/(kg·d), respectively, daily for 15 days), each group containing ten rats. After 10 days of administration, the blank control group was given normal saline by intraperitoneal injection, while a single intraperitoneal i njection of CDDP (7 mg/kg) was carried out in the remaining groups. At the end of the administration period, all rats were sacrificed to determine testicular and epididymal weights, epididymal sperm count and motility, GSH-Px and SOD activities, and GSH contents. Apoptosis rates of testicular cells were tested by flow cytometer, and histopathological characteristics were observed under a microscope. Results: Testicular and epididymal weights, epididymal sperm count and motility, GSH-Px and SOD activities, and GSH levels were signi?cantly decreased whereas the level of MDA and apoptosis rates were signi?cantly increased in rats from the CDDP group. GSPE treatment signi?cantly attenuated the CDDP-induced loss of testicular and epididymal weights and the dysfunction of reproductive organs, oxidative stress, lipid peroxidation and cell apoptosis. Conclusion: GSPE has a dose-dependent protective effect against CDDP-induced testicular toxicity in rats. This protective effect seems to be closely associated with the suppression of oxidative stress and cell apoptosis.
cisplatin; grape seed proanthocyanidin extracts; testicular toxicity; oxidative stress; apoptosis
R318.0
A
1002-6630(2015)15-0189-06
10.7506/spkx1002-6630-201515035
2014-01-29
北京聯合大學校級科研項目重點實驗室開放課題(Zk70201502)
連燕娜(1987—),女,碩士研究生,研究方向為功能性食品生化作用。E-mail:l ianyanna2009@sina.com
高麗萍(1962—),女,教授,博士,研究方向為功能性食品生化作用。E-mail:gaolip62@163.com
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中國合理用藥探索(2022年1期)2022-11-26 00:22:32
鄉村科技(2021年33期)2021-03-16 02:26:54
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
