不同發(fā)育階段椰心葉甲對(duì)3種殺蟲(chóng)劑的敏感性
2015-10-21 16:47:39林玉英金濤王謹(jǐn)金啟安溫海波彭正強(qiáng)
熱帶作物學(xué)報(bào) 2015年11期
林玉英 金濤 王謹(jǐn) 金啟安 溫海波 彭正強(qiáng)
摘 要 測(cè)定不同發(fā)育階段椰心葉甲的藥劑敏感性差異。結(jié)果表明,低齡幼蟲(chóng)和成蟲(chóng)對(duì)高效氯氰菊酯和阿維菌素的敏感性高于高齡幼蟲(chóng)、蛹和卵。幼蟲(chóng)和成蟲(chóng)對(duì)啶蟲(chóng)脒的敏感性高于蛹和卵。卵的藥劑敏感性最低。低齡幼蟲(chóng)期為最佳防治適期。椰心葉甲對(duì)3種殺蟲(chóng)劑的敏感性大小為高效氯氰菊酯>阿維菌素>啶蟲(chóng)脒。隨著幼蟲(chóng)的發(fā)育,其體重由小變大,對(duì)高效氯氰菊酯和啶蟲(chóng)脒的敏感性降低,而幼蟲(chóng)對(duì)阿維菌素的敏感性與體重相關(guān)不顯著。
關(guān)鍵詞 椰心葉甲;發(fā)育階段;藥劑敏感性
中圖分類(lèi)號(hào) TQ450.2 文獻(xiàn)標(biāo)識(shí)碼 A
Abstract The differences of insecticide sensitivities of various developmental stages of Brontispa longissima (Gestro)were studied. The result showed that the insecticide sensitivities of low instar larvae and adults to beta-cypermethrin and avermectin were higher than those of higher instar larvae, pupae and eggs. The insecticide sensitivities of larvae and adults to acetamiprid were higher than those of pupae and eggs. The insecticide sensitivities of eggs were the lowest. The low instar larvae stages were the optimum control period. The susceptibility of B. longissima to 3 kinds of insecticides was the highest to beta-cypermethrin, followed by avermectin and acetamiprid. The insecticide sensitivities to beta-cypermethrin and acetamiprid decreased, as the weight increased from small to large with the development of lavae, however the insecticide sensitivities of larvae to avermectin were not significantly related to weight.
Key words Brontispa longissima (Gestro);Developmental stage;Insecticide sensitivity
doi 10.3969/j.issn.1000-2561.2015.11.025
椰心葉甲[Brontispa longissima(Gestro)], 屬鞘翅目(Coleoptera)鐵甲科(Hispidae), 危害以椰子為主的棕櫚科植物,是一種重大危險(xiǎn)性外來(lái)有害生物。其成蟲(chóng)和幼蟲(chóng)均取食棕櫚科植株未展開(kāi)的心葉表皮組織,形成與葉脈平行的狹長(zhǎng)褐色條斑,心葉展開(kāi)后呈大型褐色壞死條斑[1],嚴(yán)重時(shí)導(dǎo)致植株大面積死亡,以殺蟲(chóng)劑為主的化學(xué)防治仍是當(dāng)前主要的應(yīng)急防控措施之一。金濤等[2]報(bào)道了東南亞地區(qū)的椰心葉甲大部分已對(duì)高效氯氰菊酯和阿維菌素產(chǎn)生了抗藥性。
殺蟲(chóng)劑的毒力與害蟲(chóng)本身(特別是不同蟲(chóng)態(tài)中的卵和蛹)的生物學(xué)特性有密切關(guān)系。黃建國(guó)等[3]報(bào)道了不同倉(cāng)庫(kù)害蟲(chóng)的成、幼蟲(chóng)對(duì)磷化氫氣體較為敏感,而卵和蛹則最不敏感。殺蟲(chóng)劑以特定的途徑進(jìn)入靶標(biāo)體內(nèi),通過(guò)干擾和破壞其正常的生長(zhǎng)發(fā)育控制害蟲(chóng),害蟲(chóng)的不同發(fā)育階段,將會(huì)帶來(lái)生理生化的差異,在藥劑防治過(guò)程中表現(xiàn)為敏感性的差異[4]。郝蕙玲等[5]報(bào)道了淡色庫(kù)蚊低齡幼蟲(chóng)對(duì)印楝素的敏感性遠(yuǎn)高于高齡幼蟲(chóng)。
化學(xué)防治一直是世界上控制害蟲(chóng)的主要方法,但由于化學(xué)殺蟲(chóng)劑的連年、大量、廣泛應(yīng)用,致使害蟲(chóng)抗藥性不斷發(fā)展,并加重了環(huán)境污染和對(duì)生態(tài)平衡的影響。抗性綜合治理的措施之一是選擇害蟲(chóng)最敏感的發(fā)育階段施藥,以提高防治效果[6]。謝圣華等[7]報(bào)道了啶蟲(chóng)脒、高效氯氰菊酯、阿維菌素對(duì)椰心葉甲具有優(yōu)良的毒殺效果,為科學(xué)合理地進(jìn)行椰心葉甲化學(xué)防治,本文研究了不同發(fā)育階段椰心葉甲對(duì)這3種不同類(lèi)型殺蟲(chóng)劑的藥劑敏感性的變化。
1 材料與方法
1.1 材料
1.1.1 供試?yán)ハx(chóng) 椰心葉甲采集于海南儋州,在室內(nèi)用椰子葉飼養(yǎng)3代,作為試驗(yàn)蟲(chóng)源。
1.1.2 供試藥劑 95%高效氯氰菊酯原粉,廣東立威化工有限公司生產(chǎn);90%阿維菌素原粉,深圳市瑞德豐農(nóng)資有限公司生產(chǎn);97.1%啶蟲(chóng)脒原粉,海南正業(yè)中農(nóng)高科股份有限公司生產(chǎn)。用丙酮將藥劑配制成母液,再稀釋至不同濃度。
1.2 方法
1.2.1 生物測(cè)定方法 采用浸漬法,使用丙酮 :雙蒸水=1 ∶ 9的藥液稀釋原藥母液5~6個(gè)濃度。每個(gè)梯度藥液量50 mL,置于燒杯中。挑選15頭1~5齡椰心葉甲幼蟲(chóng)、蛹、成蟲(chóng),或15粒卵置于指形浸蟲(chóng)器中,浸入到含有藥液的燒杯,輕震3 s取出,倒出試蟲(chóng),放置于卷紙上吸干藥液。將試蟲(chóng)放入2~4片椰子葉中,置于直徑9 cm培養(yǎng)皿中。對(duì)照組使用丙酮 ∶ 雙蒸水=1 ∶ 9的處理液浸漬。每個(gè)濃度重復(fù)3次。根據(jù)藥理的差異,高效氯氰菊酯處理組和啶蟲(chóng)脒處理組在處理后24 h,阿維菌素處理組在處理后72 h,檢查記錄幼蟲(chóng)或成蟲(chóng)死亡蟲(chóng)數(shù),計(jì)算死亡率;檢查記錄各處理組的卵孵化蟲(chóng)數(shù)和蛹羽化蟲(chóng)數(shù),計(jì)算孵化率和羽化率。幼蟲(chóng)和成蟲(chóng)以輕觸蟲(chóng)體不能產(chǎn)生明顯爬行視為死亡。
1.2.2 體重的測(cè)定 從同一批供試?yán)ハx(chóng)中選取1、2、3、4、5齡幼蟲(chóng)30頭,稱(chēng)量體重,重復(fù)10次。
1.3 數(shù)據(jù)處理
所有試驗(yàn)數(shù)據(jù)均用SPSS處理,計(jì)算出致死中濃度(LC50)、致死90%濃度(LC90)、毒力回歸式的斜率及卡方值 2,將殺蟲(chóng)劑對(duì)不同齡期椰心葉甲幼蟲(chóng)的LC50值與體重進(jìn)行Pearson相關(guān)性分析。
2 結(jié)果與分析
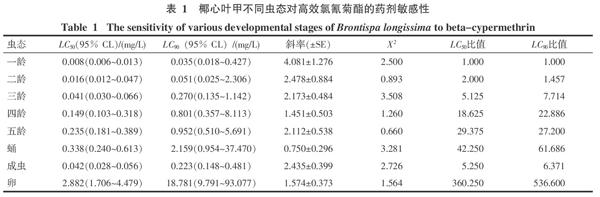
2.1 椰心葉甲不同蟲(chóng)態(tài)對(duì)高效氯氰菊酯的藥劑敏感性
由表1可知,高效氯氰菊酯對(duì)各蟲(chóng)態(tài)的LC50和LC90從小到大依次為一齡、二齡、三齡幼蟲(chóng)、成蟲(chóng)、四齡、五齡幼蟲(chóng)、蛹、卵。高效氯氰菊酯對(duì)一齡、二齡、三齡幼蟲(chóng)、成蟲(chóng)、四齡、五齡幼蟲(chóng)、蛹、卵的LC50值分別為0.008、0.016、0.041、0.042、0.149、0.235、0.338、2.882 mg/L,以一齡幼蟲(chóng)的LC50為參照,一齡幼蟲(chóng)的藥劑敏感性為二齡、三齡幼蟲(chóng)、成蟲(chóng)、四齡、五齡幼蟲(chóng)、蛹和卵藥劑敏感性的2.000、5.125、5.250、18.625、29.375、42.250和360.25倍,表明不同蟲(chóng)態(tài)個(gè)體對(duì)高效氯氰菊酯的敏感性差異很大,毒力可以相差360倍之多。高效氯氰菊酯對(duì)一齡、二齡、三齡幼蟲(chóng)和成蟲(chóng)的毒力比對(duì)四齡、五齡幼蟲(chóng)、蛹和卵的毒力高很多。一齡幼蟲(chóng)的藥劑敏感性最高;卵的藥劑敏感性最低。
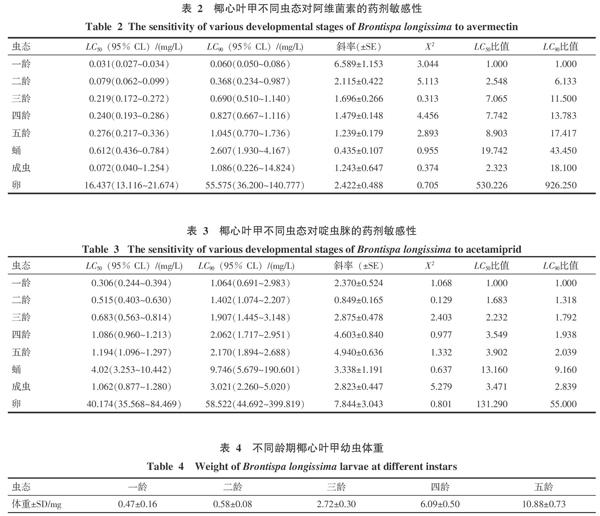
2.2 椰心葉甲不同蟲(chóng)態(tài)對(duì)阿維菌素的藥劑敏感性
椰心葉甲各蟲(chóng)態(tài)對(duì)阿維菌素的敏感性變化見(jiàn)表2。阿維菌素對(duì)椰心葉甲的LC50和LC90值一齡幼蟲(chóng)<成蟲(chóng)<二齡幼蟲(chóng)<三齡幼蟲(chóng)<四齡幼蟲(chóng)<五齡幼蟲(chóng)<蛹<卵,椰心葉甲幼蟲(chóng)從一齡到五齡對(duì)阿維菌素的敏感性逐漸降低。一齡幼蟲(chóng)的藥劑敏感性最高,LC50為0.031 mg/L;以一齡幼蟲(chóng)的LC50為參照,一齡幼蟲(chóng)的藥劑敏感性為成蟲(chóng)、二齡、三齡、四齡、五齡幼蟲(chóng)、蛹和卵藥劑敏感性的2.323、2.548、7.065、7.742、8.903、19.742和530.226倍,阿維菌素對(duì)一齡幼蟲(chóng)、成蟲(chóng)和二齡幼蟲(chóng)的毒力比對(duì)三齡、四齡、五齡幼蟲(chóng)、蛹和卵的毒力高很多。卵對(duì)阿維菌素的藥劑敏感性最低,LC50為16.437 mg/L,說(shuō)明不同蟲(chóng)態(tài)的椰心葉甲對(duì)阿維菌素的藥劑敏感性不同。
2.3 椰心葉甲不同蟲(chóng)態(tài)對(duì)啶蟲(chóng)脒的藥劑敏感性
啶蟲(chóng)脒對(duì)椰心葉甲各蟲(chóng)態(tài)的毒力見(jiàn)表3。啶蟲(chóng)脒對(duì)各蟲(chóng)態(tài)的LC50和LC90一齡幼蟲(chóng)<二齡幼蟲(chóng)<三齡幼蟲(chóng)<成蟲(chóng)<四齡幼蟲(chóng)<五齡幼蟲(chóng)<蛹<卵。幼蟲(chóng)隨著齡期的增加,其LC50值上升,對(duì)啶蟲(chóng)脒的敏感性降低,一齡幼蟲(chóng)的藥劑敏感性為五齡幼蟲(chóng)藥劑敏感性的3.902倍。一齡幼蟲(chóng)的LC50為0.306 mg/L,以一齡幼蟲(chóng)的LC50為參照,一齡幼蟲(chóng)的藥劑敏感性為二齡、三齡、成蟲(chóng)、四齡、五齡幼蟲(chóng)、蛹和卵藥劑敏感性的1.683、2.232、3.471、3.549、3.902、13.160、131.29倍,啶蟲(chóng)脒對(duì)一齡、二齡、三齡、成蟲(chóng)、四齡、五齡幼蟲(chóng)的毒力比對(duì)蛹和卵的毒力高很多,說(shuō)明不同蟲(chóng)態(tài)的椰心葉甲對(duì)啶蟲(chóng)脒的藥劑敏感性不同。
2.4 不同齡期椰心葉甲體重與LC50值的相關(guān)性
將高效氯氰菊酯處理組和啶蟲(chóng)脒處理組不同齡期椰心葉甲幼蟲(chóng)的LC50值(表1、3)與體重(表4)進(jìn)行Pearson相關(guān)性分析,得出相關(guān)系數(shù)>0呈正相關(guān),p<0.05,說(shuō)明高效氯氰菊酯和啶蟲(chóng)脒對(duì)該蟲(chóng)的LC50值受體重顯著正影響,隨著幼蟲(chóng)的發(fā)育其體重由小變大,其LC50值也由小變大,即對(duì)高效氯氰菊酯和啶蟲(chóng)脒的敏感性由大變小。將阿維菌素處理組不同齡期椰心葉甲幼蟲(chóng)的LC50值(表2)與體重(表4)進(jìn)行Pearson相關(guān)性分析,得出p>0.05,說(shuō)明幼蟲(chóng)對(duì)阿維菌素的敏感性與體重相關(guān)不顯著。
3 討論與結(jié)論
不同蟲(chóng)態(tài)椰心葉甲個(gè)體對(duì)藥劑的敏感性差異很大,幼蟲(chóng)從一齡到五齡的發(fā)育階段,其藥劑敏感性逐漸降低。幼蟲(chóng)從一齡發(fā)育到五齡,其表皮增厚,幾丁質(zhì)含量增多,可降低殺蟲(chóng)劑的穿透性,延緩了殺蟲(chóng)劑到達(dá)靶標(biāo)的時(shí)間,增加了殺蟲(chóng)劑在幼蟲(chóng)體內(nèi)的降解,減少了殺蟲(chóng)劑到達(dá)靶標(biāo)的量,從而增強(qiáng)對(duì)殺蟲(chóng)劑的耐受性。已有研究報(bào)道螺旋粉虱若蟲(chóng)、蚊蟲(chóng)幼蟲(chóng)、斜紋夜蛾幼蟲(chóng)和甜菜夜蛾幼蟲(chóng)藥劑敏感性隨著齡期的增大而降低[8-12],本研究結(jié)果再次證實(shí),椰心葉甲幼蟲(chóng)隨著齡期的增加,藥劑敏感性逐漸下降。廖永林等[8]報(bào)道了螺旋粉虱1~4齡若蟲(chóng)對(duì)殺蟲(chóng)劑的敏感性隨著蟲(chóng)齡的增加而降低,提出若蟲(chóng)隨著蟲(chóng)齡的增大而體內(nèi)的解毒功能增強(qiáng)可能是其原因之一。李士根等[9]報(bào)道了蚊蟲(chóng)低齡幼蟲(chóng)解毒酶基因尚未完全表達(dá)而不能合成足夠的解毒酶,是其對(duì)殺蟲(chóng)劑較敏感的原因之一。姚洪渭等[13]和Yu等[14-15]認(rèn)為高齡幼蟲(chóng)體內(nèi)解毒酶活性水平往往較高,且易被誘導(dǎo),同時(shí)表皮結(jié)構(gòu)更加完善,從而對(duì)藥劑更具耐性。不同齡期椰心葉甲幼蟲(chóng)解毒酶活性對(duì)其藥劑敏感性的影響還有待進(jìn)一步研究。椰心葉甲低齡幼蟲(chóng)和成蟲(chóng)對(duì)高效氯氰菊酯和阿維菌素的敏感性高于高齡幼蟲(chóng)、蛹和卵,幼蟲(chóng)和成蟲(chóng)對(duì)啶蟲(chóng)脒的敏感性高于蛹和卵,說(shuō)明椰心葉甲成蟲(chóng)期的藥劑敏感性較高,由于成蟲(chóng)跗節(jié)的表皮極薄,并常帶有化學(xué)感受器,爪間膜有通向外部的腺細(xì)胞,這些腺細(xì)胞的分泌能使藥劑溶化,因而更易穿透進(jìn)入[16]。椰心葉甲蛹的藥劑敏感性較低,卵的藥劑敏感性最低,蛹?xì)ず吐褮び捎诤袔锥≠|(zhì)而能夠減緩藥劑穿透,降低藥劑對(duì)蛹和卵的毒力。本研究結(jié)果顯示椰心葉甲卵的藥劑敏感性最低,而二斑葉螨抗性種群成螨對(duì)蟲(chóng)螨腈、噠螨靈、氟蟲(chóng)脲、三氯殺螨醇、三唑錫、三氟氯氰菊酯、雙甲脒、氧樂(lè)果、甲氰菊酯藥劑敏感性低于卵的敏感性[17],這可能與抗性成螨可以爬行,具有行為抗性,或者抗性螨體內(nèi)各種代謝酶的活性比卵內(nèi)的活性高,從而比卵更易產(chǎn)生抗藥性。
3種殺蟲(chóng)劑對(duì)椰心葉甲的LC50和LC90從低到高為高效氯氰菊酯<阿維菌素<啶蟲(chóng)脒,表明椰心葉甲對(duì)高效氯氰菊酯最為敏感;對(duì)阿維菌素較敏感;對(duì)啶蟲(chóng)脒的藥劑敏感性較低。該蟲(chóng)對(duì)不同類(lèi)型藥劑的耐受劑量有差異,表明了在田間使用多種殺蟲(chóng)劑防治椰心葉甲過(guò)程中,應(yīng)該根據(jù)椰心葉甲各發(fā)育時(shí)期的耐藥性來(lái)調(diào)整殺蟲(chóng)劑的施用劑量,建議不同類(lèi)型的殺蟲(chóng)劑隔代輪換使用或把作用機(jī)制不同的殺蟲(chóng)劑混用,可以達(dá)到增效的目的。同時(shí),防治該蟲(chóng)應(yīng)抓住低齡幼蟲(chóng)期,以降低農(nóng)藥使用劑量,避免害蟲(chóng)產(chǎn)生抗性,以達(dá)到理想的防治效果。
參考文獻(xiàn)
[1] 呂寶乾, 陳義群, 包 炎,等. 引進(jìn)天敵椰甲截脈姬小蜂防治椰心葉甲的可行性探討[J]. 昆蟲(chóng)知識(shí), 2005, 42(3): 254-258.
[2] Lin Y Y, Jin T, Jin Q A, et al. Differential susceptibilities of Brontispa longissima(Coleoptera: Hispidae)to insecticides in Southeast Asia[J]. Journal of Economic Entomology, 2012, 105(3): 988-993.
[3] 黃建國(guó), 關(guān)永嘉. 探討米象不同蟲(chóng)態(tài)對(duì)磷化氫的敏感性[J]. 鄭州糧食學(xué)院學(xué)報(bào), 1982(3): 12-16.
[4] 徐 菲, 孟鳳霞, 劉起勇,等. 家蠅雄成蟲(chóng)對(duì)殺蟲(chóng)劑的敏感性測(cè)定[J]. 中國(guó)媒介生物學(xué)及控制雜志, 2005, 16(3): 187-189.
[5] 郝蕙玲, 孫錦程. 印楝素對(duì)淡色庫(kù)蚊的生物活性研究[J]. 中華衛(wèi)生殺蟲(chóng)藥械, 2008, 14(1): 24-26.
[6] 唐振華. 昆蟲(chóng)抗藥性及其治理[M]. 北京: 農(nóng)業(yè)出版社, 1993, 208-332.
[7] 謝圣華, 陳綿才, 芮 凱,等. 36種殺蟲(chóng)制劑對(duì)椰心葉甲的毒力測(cè)定[J]. 海南大學(xué)學(xué)報(bào)自然科學(xué)版, 2007, 25(1): 57-61.
[8] 廖永林, 張 揚(yáng), 曾東強(qiáng),等. 多種藥劑對(duì)螺旋粉虱不同蟲(chóng)態(tài)的毒力測(cè)定[J]. 熱帶作物學(xué)報(bào), 2011, 32(1): 163-167.
[9] 李士根, 公茂慶, 王新國(guó),等. 不同品系淡色庫(kù)蚊各發(fā)育階段的非特異性酯酶和乙酰膽堿酯酶活力變化[J]. 地方病通報(bào),2002, 17(4): 15-18.
[10] 張 鵬, 李 慧, 王秋紅, 等. 齡期、 飼養(yǎng)條件和測(cè)定方法對(duì)韭菜遲眼蕈蚊藥劑敏感性的影響[J]. 環(huán)境昆蟲(chóng)學(xué)報(bào), 2014, 36(5): 730-736.
[11] 蘭亦全, 黃福特,陳海華. 寄主植物和試蟲(chóng)齡期對(duì)斜紋夜蛾藥劑敏感性的影響[J]. 武夷科學(xué), 2013, 29: 203-206.
[12] 彭 梅,鄧新平. 甜菜夜蛾不同齡期幼蟲(chóng)藥劑敏感性及酶活性差異[J]. 西南農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版), 2005, 27(2): 173-175, 183.
[13] 姚洪渭, 葉恭銀,程家安. 寄主植物影響害蟲(chóng)藥劑敏感性的研究進(jìn)展[J]. 昆蟲(chóng)學(xué)報(bào), 2002, 45(2): 253-264.
[14] Yu S J. Induction of detoxifying enzymes by allelochemicals and host plants in the fall armyworm[J]. Pesticide Biochemistry and Physiology, 1983, 19: 330-336.
[15] Yu S J, Hsu E L. Induction of detoxification enzymes in phytophagous insects: roles of insecticide synergists,larval age,and species[J]. Archives of Insect Biochemistry and Physiology,1993, 24(1): 21-23.
[16] 唐振華, 畢 強(qiáng). 殺蟲(chóng)劑作用的分子行為[M]. 上海遠(yuǎn)東出版社,2003:57.
[17] 王開(kāi)運(yùn), 趙衛(wèi)東, 姜興印,等. 十種殺螨劑對(duì)二斑葉螨抗性種群不同發(fā)育階段的毒力比較[J]. 農(nóng)藥, 2002, 41(3): 29-31.
