狂犬病病毒單質粒拯救系統的建立及應用
2015-07-21 03:13:32孫玉章許運斌叢彥龍KarlKlausConzelmann
畜牧獸醫學報 2015年11期
孫玉章,何 彪,許運斌,叢彥龍,Karl-Klaus Conzelmann
(1.中國動物衛生與流行病學中心,青島 266032;2.軍事醫學科學院軍事獸醫研究所,長春 130122;3.浙江大學動物科學學院,杭州 310020;4.吉林大學動物醫學學院,長春 130062;5.慕尼黑大學基因中心,慕尼黑81377)
狂犬病(rabies)是由狂犬病病毒(rabies virus,RABV)引起的以侵犯中樞神經系統為主的急性人獸共患傳染病,病死率幾乎高達100%。據世界衛生組織(World Health Organization,WHO)報道全球每年有約55000人死于狂犬病,其中絕大多數發生在亞洲、非洲等發展中國家[1]。狂犬病病毒是彈狀病毒科(Rhabdoviridae)狂犬病病毒屬(Lyssavirus)成員,其基因組為不分節段的負鏈RNA病毒。RABV基因組編碼5種結構蛋白,依次為核蛋白(nucleoprotein,N)、磷酸化蛋白 (phosphoprotein,P)、基質蛋白(matrix protein,M)、糖蛋白(glycoprotein,G)和RNA依賴的RNA聚合酶(RNA dependent RNA polymerase,RdRp/L)[2]。目前,學術界一般將狂犬病病毒屬劃分為7個基因型,其中6個基因型的RABV均以蝙蝠為宿主和媒介生物(基因III型,即MOKV的確切宿主尚未確定),另外還有一些新近分離出的RABV毒株尚待分型[3]。
狂犬病病毒基因組RNA只有與核蛋白形成核糖核蛋白復合物(ribonucleoprotein,RNP)才能在磷蛋白的催化下被RNA依賴的RNA聚合酶蛋白識別并作為轉錄和復制的模板[4]。因此,經典的RABV的反向遺傳操作系統除了構建具有精確末端序列的全長基因組cDNA克隆外,還需要構建表達核蛋白、磷蛋白和聚合酶大蛋白的真核表達質粒共同轉染適當的細胞系才可能獲得具有感染性的病毒顆粒[5]。
本研究在已建立SAD L16株高效拯救的反向遺傳操作平臺基礎上[6],以SAD L16株全長基因組cDNA克隆為骨架,利用內部核糖體進入位點(internal ribosome entry site,IRES)分別構建了5個全長cDNA克隆[7]并直接轉染BSR T7/5細胞系,獲得了2株重組病毒。通過進一步鑒定和分析重組狂犬病病毒株的生物學特性,本研究首次建立起RABV的單質粒拯救系統,并以此為平臺開展了一系列研究。
1 材料與方法
1.1 病毒與細胞
BSR T7/5細胞系由本室保存,以含10%胎牛血清的GMEM培養。狂犬病病毒SAD L16株由本室保存,其全長cDNA克隆質粒pSAD-L16由本室構建并保存。分別克隆有腦心肌炎病毒(encephalomyocarditis virus,EMCV)IRES 序列、脊髓灰質炎病毒(polivirus,PV)IRES序列和明脈扁刺蛾病毒(Thosea asigna virus,TaV)2A序列的pRLCMV載體質粒均由本室構建并保存。
1.2 主要試劑
Phusion DNA聚合酶購自Finnzymes公司,反轉錄聚合酶購自Roche公司,T4DNA連接酶購自NEB公司,質粒快速抽提試劑盒和凝膠快速回收試劑盒均購自QIAGEN公司,轉染試劑Lipofectamine 2000購自Invitrogen公司,CaPO4轉染試劑盒購自Stratagene公司,其他限制性內切酶主要購自NEB或Fermentas公司,細胞培養血清及相關試劑均購自Invitrogen公司,其他試劑或化學藥品均購自Invitrogen、Roche或Sigma公司。
1.3 重組全長cDNA克隆的構建
根據全長cDNA感染性克隆pSAD-L16 N、P和L基因上游序列的不同分別設計多對引物擴增EMCVIRES 序列(e)、PVIRES 序列(p)和 TaV 2A序列(t),將其分別克隆入pSAD-L16載體的對應位置中,構建成5個全長cDNA克隆:pSAD-eNePeL(Ⅰ)、pSAD-eNtPeL(Ⅱ)、pSAD-NeNePeL(Ⅲ )、pSAD-NeNpPeL (Ⅳ )和 pSAD-NeNtPeL(Ⅴ)。所有構建的cDNA克隆經測序正確后大量制備并純化。5個全長cDNA克隆的構建策略如圖1所示。
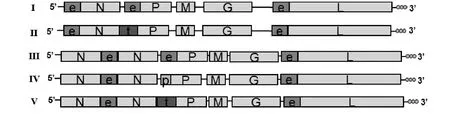
圖1 重組全長cDNA克隆構建模式Fig.1 Construction of 5full-length cDNA clones based on pSAD-L16
1.4 重組病毒的拯救
轉染前16h將BSR T7/5細胞分鋪到6孔板,用含10%胎牛血清的GMEM培養。轉染前1h更換為不含血清的GMEM培養。CaPO4法分別轉染10μg全長cDNA克隆質粒至每孔細胞中,輕晃混勻。同時設單質粒組(分別單獨輔助轉染pTiT-N、pTiT-P或pTiT-L質粒)和常規組(同時轉染pTiTN、pTiT-P和pTiT-L 3個輔助質粒)4個對照組。37℃培養6h,PBS清洗3次后更換含10%胎牛血清的GMEM繼續培養。轉染后每3d取培養液感染新的BSR T7/5細胞并以FITC標記的鼠抗狂犬病病毒N蛋白單克隆抗體進行檢測,直到第12天。獲救的每一代次的重組狂犬病病毒首先進行RTPCR并測序,序列完全正確的病毒測定滴度后以MOI=0.01接種于T75細胞培養瓶,每72h收獲一次培養液,連續收獲3次,并儲存于-80℃超低溫冰箱。
1.5 重組狂犬病病毒生物學特性的鑒定
重組狂犬病病毒的滴度測定:將收獲的細胞培養上清液以不含胎牛血清的GMEM連續做10倍梯度稀釋后取100μL接種于96孔板中的單層BSR T7/5細胞,于37℃培養48h,棄細胞上清液,PBS清洗后用80%丙酮冷藏固定20min,洗棄冷丙酮后以FITC標記的鼠抗狂犬病病毒N蛋白單克隆抗體37℃染色2h,PBS清洗3次后置于熒光顯微鏡下觀察并計數,計算病毒終釋滴度。
生長動力學曲線的測定:拯救重組病毒以MOI=0.01接種于T25細胞培養瓶,6h后收集細胞培養液并更換新鮮GMEM培養液,每24h收獲一次,直到96h。收獲的細胞培養液梯度稀釋后接種于96孔板,37℃培養48h后以FITC標記的鼠抗狂犬病病毒N蛋白單克隆抗體進行檢測,計算終釋病毒滴度并繪制多步生長曲線。
重組狂犬病病毒的遺傳穩定性:拯救重組病毒以MOI=1接種于BSR T7/5細胞,每96~120h收獲細胞培養上清液,分別進行轉移到新BSR T7/5細胞中繼續傳代和轉染96孔板測定病毒滴度,連續傳代20次,每5代收集一次細胞培養上清液進行RT-PCR并測序。
2 結 果
2.1 重組狂犬病病毒的拯救
在完全不依靠轉染輔助質粒的情況下,只有pSAD-NeNePeL(Ⅲ)和pSAD-NeNtPeL(Ⅴ)能夠成功拯救出重組狂犬病病毒,pSAD-NeNePeL直接轉染成功率較高(12/12),pSAD-NeNtPeL 成功率較低(8/14),單獨轉染任何一種輔助質粒pTiT-N、pTiT-P或pTiT-L對拯救效率均無顯著影響,轉染后第6天均可在細胞培養上清液中檢測到病毒顆粒(圖2)。
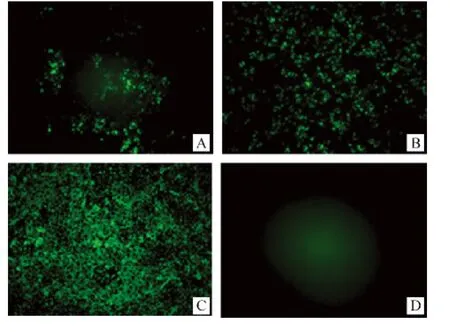
圖2 重組狂犬病病毒的DFA檢測Fig.2 Identification of recombinant RABVs with FITC-antiN-Mab
2.2 重組狂犬病病毒的增殖動態
初代獲救的重組狂犬病病毒的多步生長曲線如圖3所示,重組狂犬病病毒SAD-NeNePeL和SADNeNtPeL均高度致弱且增殖緩慢。
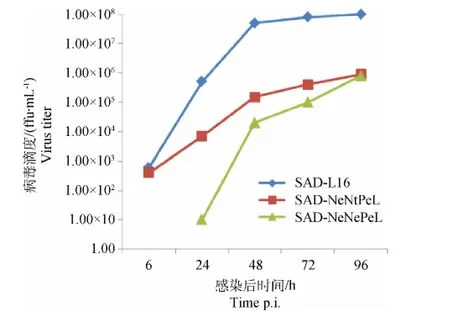
圖3 重組狂犬病病毒多步生長曲線Fig.3 Multistep growth curves of recombinant RABVs
SAD-NeNePeL和 SAD-NeNtPeL 2株重組狂犬病病毒在終釋滴度均能形成與野生型親本毒株SAD-L16株大小形態不同的熒光灶(圖4)。
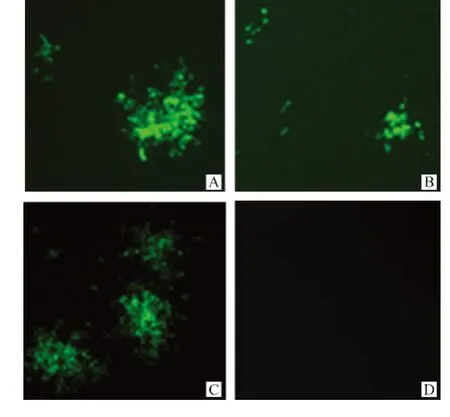
圖4 重組狂犬病病毒的熒光灶形態Fig.4 Different phenotypes in foci size of recombinant RABVs
2.3 重組狂犬病病毒的遺傳穩定性
重組狂犬病病毒SAD-NeNePeL在BSR T7/5細胞中僅能傳3~5代,RT-PCR結果顯示在第3代的細胞培養液上清中就已經存在不同水平的重組株,且表現出不同的生長動力學特性(未發表數據)。重組狂犬病病毒SAD-NeNtPeL在BSR T7/5細胞中則能穩定傳代20次,且RT-PCR結果顯示序列高度穩定。
3 討 論
經典狂犬病病毒反向遺傳系統的建立除需精確構建病毒基因組的全長cDNA克隆外,還需提供分別表達N、P和L 3種蛋白質的輔助質粒,才可能形成RNP復合物并起始病毒基因組的轉錄和翻譯。順式作用元件IRES序列的發現[8-9]給我們的研究提供了另外一種可能:通過在狂犬病病毒基因組全長cDNA克隆中N、P和L基因上游克隆入相應序列以實現單一質粒同時表達多種mRNA。因此,我們分別設計了包含EMCVIRES序列、PVIRES序列或TaV 2A序列的多個全長cDNA克隆,并先后建立起了三質粒拯救系統和二質粒拯救系統,最終有2個單質粒拯救系統成功拯救出重組狂犬病病毒株。
單質粒拯救系統在負鏈分節段的流感病毒已有報道[10],但其采用的方法并不適用于負鏈不分節段的狂犬病病毒。通過引入順式作用元件IRES序列建立的單質粒拯救系統獲救的SAD-NeNePeL株具有較高的拯救成功率,但基因組間的重配概率較高,進一步的研究發現在細胞培養上清液中至少存在3種基因重組株且呈現不同的生長動力學特性;而SAD-NeNtPeL則可以穩定的表達多順反子mRNA且具有相當的遺傳穩定性。值得注意的是在N基因上游引入IRES序列構建的多個全長cDNA克隆無一拯救成功,進一步的研究發現導致拯救失敗的原因是在N基因氨基端存在一個此前未曾報道過的順式調控元件(數據未發表),因此本研究通過引入表達更高效的EMCVIRES序列串聯起2個N基因,最終無需提供輔助質粒pTiT-N即可拯救出重組狂犬病病毒。
利用建立的狂犬病病毒單質粒拯救系統,本室先后開展了基于Pol-II聚合酶拯救系統的建立以及更高效的二質粒拯救系統的建立等研究。這些反向遺傳操作系統的建立為深入研究狂犬病病毒基因組結構與功能、狂犬病病毒分子致病機制、狂犬病病毒與宿主相互作用以及新型嗜神經性病毒載體和轉基因動物模型等提供了一個有力的研究工具和基礎的技術平臺。
(References):
[1]KNOBEL D L,CLEAVELAND S,COLEMAN P G,et al.Re-evaluating the burden of rabies in Africa and Asia[J].Bull World Health Organ,2005,83(5):360-368.
[2]TAKAYAMA-ITO M,INOUE K,SHOJI Y,et al.A highly attenuated rabies virus HEP-Flury strain reverts to virulent by single amino acid substitution to arginine at position 333in glycoprotein[J].Virus Res,2006,119(2):208-215.
[3]DIETZGEN R D,KUZMIN I V.Rhabdoviruses:molecular taxonomy,evolution,genomics,ecology,hostvector interactions,cytopathology and control[M].London:Caister Academic Press,2012:37-57.
[4]SHOJI Y,INOUE S,NAKAMICHI K,et al.Generation and characterization of P gene-deficient rabies virus[J].Virology,2004,318(1):295-305.
[5]SCHNELL M J,MEBATSION T,CONZELMANN K K.Infectious rabies viruses from cloned cDNA[J].EMBO J,1994,13(18):4195-4203.
[6]GHANEM A,KERN A,CONZELMANN K K.Significantly improved rescue of rabies virus from cDNA plasmids[J].Eur J Cell Biol,2012,91(1):10-16.
[7]MARSCHALEK A,FINKE S,SCHWEMMLE M,et al.Attenuation of rabies virus replication and virulence by picornavirus internal ribosome entry site elements[J].J Virol,2009,83(4):1911-1919.
[8]PIJLMAN G P,ROODE E C,FAN X,et al.Stabilized baculovirus vector expressing a heterologous gene and GP64from a single bicistronic transcript[J].J Bio-technol,2006,123(1):13-21.
[9]WAGSTAFF M J,LILLEY C E,SMITH J,et al.Gene transfer using a disabled herpes virus vector containing the EMCV IRES allows multiple gene expression in vitro and in vivo[J].Gene Ther,1998,5(11):1566-1570.
[10]ZHANG X,KONG W,ASHRAF S,et al.A one-plasmid system to generate influenza virus in cultured chicken cells for potential use in influenza vaccine[J].J Virol,2009,83(18):9296-9303.
