高等植物適應鹽逆境研究進展
2015-06-01 12:15:17張金林李惠茹郭姝媛王鎖民施華中韓慶慶包愛科馬清
草業學報 2015年12期
張金林,李惠茹,郭姝媛,王鎖民,施華中,韓慶慶,包愛科,馬清
(1.草地農業生態系統國家重點實驗室,蘭州大學草地農業科技學院,甘肅 蘭州 730020;2.美國德克薩斯理工大學化學與生物化學系,德克薩斯 拉伯克 TX79409,美國)
?
高等植物適應鹽逆境研究進展
張金林1*,李惠茹1,郭姝媛1,王鎖民1,施華中2,韓慶慶1,包愛科1,馬清1
(1.草地農業生態系統國家重點實驗室,蘭州大學草地農業科技學院,甘肅 蘭州 730020;2.美國德克薩斯理工大學化學與生物化學系,德克薩斯 拉伯克 TX79409,美國)
土壤鹽堿化已經成為制約農作物生長及產量的重要因子之一,尋求將鹽堿化對植物的危害降低到最小程度的策略勢在必行。關于植物對鹽逆境適應能力的研究已成為全球關注的熱點, 如何提高植物的耐鹽能力也已成為研究的重中之重。深入探究高等植物適應鹽逆境的機制,有助于提高植物耐鹽性,增加作物產量和保護生態環境。本文就高等植物適應鹽逆境的重點研究進展,綜述了鹽脅迫對植物的危害;植物耐鹽的生理機制,包括滲透調節、營養元素平衡和增強抗氧化脅迫等;植物耐鹽相關基因研究進展,包括離子轉運蛋白基因、滲透調節相關基因、信號傳導相關基因和細胞抗氧化相關基因等;提高植物耐鹽性的途徑。最后針對今后植物適應鹽逆境方面的研究方向進行了展望。
高等植物;鹽脅迫;耐鹽性;耐鹽基因
地球表面70%的面積被海洋覆蓋,海水中Na+濃度大約在500 mmol/L以上,而K+濃度僅為9 mmol/L[1];地殼當中鈉元素含量為2.8%,而鉀元素為2.6%[2]。因此,地球被稱為“咸行星”[1]。在中世紀,鹽(氯化鈉,NaCl)被稱為“白色金子”,因為它是和黃金一樣昂貴的商品[3]。鹽堿土在陸地生態系統上分布廣泛,全世界鹽漬土面積約l0億hm2[4]。在全球的干旱和半干旱地區,約有50%的灌溉土地受到鹽堿化的影響,區域內的非灌溉土地同樣會發生鹽堿化[4]。中國土地鹽漬化大約占到全球鹽漬土面積的1/10[5],從濱海到內陸,從低地到高原都分布著不同類型的鹽漬化土地[6]。在長期的自然進化過程中,海洋植物保留了對高濃度鹽分的耐受性,然而,大多數陸生高等植物在進化過程中喪失了這種耐受性而采取了一種甜土植物的生活方式[7]。土壤中可溶性鹽分過多,會對植物造成傷害[4,8]。因此,土壤鹽堿化已經成為影響農作物生長及產量的重要因子之一,關于植物對鹽逆境適應能力的研究已成為全球關注的熱點。如何提高植物的耐鹽能力已成為研究的重中之重。高等植物對鹽逆境的適應是一個綜合的生物調節過程,需要各種生理生化過程的協同作用,而非某種單一的過程就能夠使植物成功地抵御鹽逆境。本文綜述了鹽脅迫對植物的危害,植物適應鹽逆境的方式,提高植物耐鹽性的途徑和植物耐鹽的相關基因研究進展,最后針對植物適應鹽逆境方面的研究進行了展望。
1 鹽逆境對植物的危害
鹽逆境對植物造成的直接危害首先表現為滲透脅迫,并且持續存在;緊接著表現為離子失調引起的毒害和營養元素的虧缺;最后引起的氧化脅迫導致膜透性的改變、生理生化代謝的紊亂和有毒物質的積累,進而引起植物生長發育和形態建成的改變(圖1)[1,7-10]。
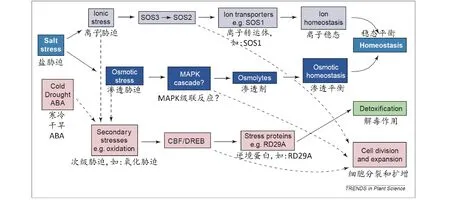
圖1 鹽脅迫對植物的危害以及植物耐鹽的主要生理機制[9]
1.1滲透脅迫
首先,在鹽脅迫下,植物種子的萌發會受到影響,一般分為滲透效應和離子效應。滲透效應引起溶液滲透勢降低而使種子吸水受阻,從而影響種子萌發;離子效應通過鹽離子(Na+、Cl-、SO42-等)直接毒害而抑制種子萌發[11]。在對一年生植物種子耐鹽機制進行研究時,發現高鹽濃度會對種子產生滲透抑制,只有脅迫減輕時才能消除這種抑制作用[12]。李昀等[13]通過對堿茅(Puccinelliadistans)、黑麥草(Loliumperenne)、野大麥(Hordeumbrevisubulatum)、鹽爪爪(Kalidiumcapsicum)、堿蓬(Suaedaglauca)五種牧草研究發現,鹽濃度升高,種子的發芽率會降低。其次,對于整株植物而言,若土壤中的鹽分過多,則會導致土壤中的水勢下降,植物細胞的水勢相對過高,導致植物吸水困難,嚴重的還會引起植物失水,引起生理干旱[7]。
1.2離子失調
土壤中的鹽分多以離子形式存在,雖然高等植物對土壤中的離子具有選擇吸收作用,但是在吸水的同時,也必定會吸收大量的鹽離子[1,7-10]。K+在細胞內可作為60多種酶的活化劑,能促進蛋白質的合成,促進糖類的合成與運輸;K+也是構成細胞滲透勢的重要成分[14]。而在根內K+可以從薄壁細胞轉運至導管,從而降低導管中的水勢,使水分能從根系表面轉運到木質部中去[15]。K+對氣孔開放有直接作用,有研究表明,K+可以維持細胞的滲透平衡,在氣孔的關閉中起作用,還可以作為許多酶的輔助因子行使生理功效[16]。土壤中K+的濃度范圍在0.2~10.0 mmol/L之間,而K+和Na+的水合半徑相似,因此高濃度的Na+會阻礙植物對K+的吸收,造成K+匱缺,從而抑制了以上提到的依賴于K+的生理生化反應的正常進行[4,7]。同時,大量Na+、Cl-進入細胞可以破壞Ca2+平衡,細胞質中游離Ca2+急劇增加,使Ca2+介導的鈣調蛋白(calmodulin,CaM)調節系統和磷酸肌醇調節系統失調,細胞代謝紊亂甚至傷害死亡[17]。此外,土壤中高濃度的鹽分還會抑制植物對NO3-和NH4+的吸收,而對NO3-吸收的抑制作用更大[18]。
1.3氧化脅迫
細胞膜是植物細胞重要的保護屏障,對物質運輸、能量傳遞、信號轉導有重要作用。細胞膜本身具有選擇透過性,因而可以調節細胞內的離子平衡,同時滿足植物生理活動的需要。然而,鹽脅迫導致的氧化脅迫會使膜的透性發生改變,一方面對離子的選擇性、流速、運輸等產生影響;另一方面,也造成了磷和有機物質的外滲,從而使得細胞的生命活動受到影響[7]。丙二醛(malondialdehyde,MDA)含量的高低和細胞質膜的透性變化是反映膜脂過氧化作用強弱和質膜破壞程度的重要指標,其含量可說明植物遭受逆境傷害的程度[19]。活性氧的增加還會破壞細胞中具有膜結構細胞器的結構,如引起線粒體DNA的突變,造成細胞衰老,導致內質網部分膨脹、線粒體數目減少而體積膨脹、液泡膜破碎、胞質降解等。
1.4光合作用受挫
鹽逆境會使得植物的光合速率下降,葉綠素是表征光合利用率的最重要的指標之一。隨著鹽濃度升高,葉綠素a、b和類胡蘿卜素含量均顯著降低,表明高鹽脅迫抑制或破壞了光系統Ⅱ的部分功能,光合作用的能量及電子傳遞受到抑制[20]。同時,鹽分過多可使磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase,PEPC)與1,5-二磷酸核酮糖羧化酶(ribulose-1,5-bisphosphate,Rubisco)活性降低,引起脅迫初期光合作用明顯下降[21]。
1.5有毒物質的積累
鹽脅迫下,由于植物細胞結構的損傷、活性氧的積累、生理代謝的破壞,植株體內蛋白質的合成速率降低,水解加速,造成植株體內氨基酸積累,會產生許多的有毒物質,如大量氮代謝的中間產物,包括NH3和某些游離氨基酸(異亮氨酸、鳥氨酸和精氨酸)轉化成有一定毒性的腐胺(如丁二胺、戊二胺等),而腐胺又可被氧化成NH3和H2O2,當它們達到一定濃度時,細胞會中毒死亡[22]。這些物質的積累,會抑制植物內相關物質的合成,使得植物生長受抑。
2 植物耐鹽的生理機制
植物受到鹽害脅迫時,會采取兩種方式來減輕鹽脅迫造成的危害:躲避鹽離子的傷害和增強對鹽脅迫的耐受性[6]。植物避鹽的方式主要分為4種:泌鹽、稀鹽、積鹽和拒鹽。泌鹽(salt excretion)是指植物在吸收鹽分后,不在體內存儲,而是通過體內的鹽腺等器官或機制排出體外,再通過雨水沖刷等方式脫鹽,從而維持植物體內的離子穩態。泌鹽被認為是一個有助于鹽生植物抗鹽的重要機制[5,23-24]。稀鹽(salt-dilution)是指植物通過吸收大量的水分、使細胞加速生長(增加薄壁細胞組織,使細胞質膨脹,增大細胞壁伸展度等)、改變形態等方法,稀釋體內鹽分,降低鹽濃度[7,23]。積鹽(salt-accumulation)是指植物體內的原生質特化后,將根部吸收的鹽分區隔化至液泡,同時,抑制鹽分從液泡內溢出,即將鹽分儲存在液泡內,作為廉價的滲透調節劑[7,23]。拒鹽(salt-exclusion)是指植物本身排斥鹽離子進入細胞,依靠其對鹽的不透性,阻止鹽分進入植物體;另一方面,植物根部通過阻止Na+向木質部的裝載和加大對Na+外排來減少植株對Na+積累。絕大多數栽培植物都屬于拒鹽能力弱的甜土植物,拒鹽能力強的鹽生植物有蘆葦、堿茅[7,23,25]。除了通過以上4種方式來躲避鹽離子的傷害,所有高等植物都可以通過以下幾個方面的生理調節過程來增強對鹽脅迫的耐受性,即滲透調節、營養元素平衡和增強抗氧化脅迫(圖1)。
2.1滲透調節
在鹽逆境脅迫下,高等植物通常會采用兩種滲透調節方式,一是在植物體內合成有機調節物質;二是積累更多的無機離子。
通常有機溶質大體可分為3類:1) 游離氨基酸(如脯氨酸),具有很大的水溶性,其疏水端可和蛋白質結合,親水端可與水分子結合,蛋白質可借助脯氨酸束縛更多的水,從而防止滲透脅迫條件下蛋白質的脫水變性;它可以維持細胞內外滲透平衡,防止水分散失[26]。2)甜菜堿,作為一種無毒的滲透調節劑和酶的保護劑,它的積累使植物細胞在鹽脅迫下保持膜的完整性,在滲透脅迫下仍能保持正常的功能。許多高等植物,尤其是藜科和禾本科植物,在受到水/鹽脅迫時積累大量甜菜堿[27]。甜菜堿對類囊體膜也有穩定作用,并顯著提高光系統Ⅱ光合放氧的穩定性;甜菜堿能保護抗氧化酶系統[超氧化物歧化酶(superoxide dismutase,SOD)、抗壞血酸過氧化物酶(ascorbate peroxidase,APX)、過氧化氫酶(catalase,CAT)等]的活性[28];甜菜堿還可以穩定生物大分子的結構與功能,解除高濃度鹽對酶的毒害作用,防止脫水誘導的蛋白質熱動力學干擾[29]。3)可溶性糖和多元醇,也可以作為滲透調節物質,調節植物細胞的滲透勢,從而增強植物的耐鹽機制[30]。
無機離子(K+,Na+和Cl-)作為滲透調節劑具有很多優點,也日益受到重視。首先,由于離子的大量存在,無需消耗物質和能量來大量合成,所以較為廉價;第二,無機離子的調節作用可以在短時間內迅速完成;第三,它們的作用也顯著高效[31]。無機離子在整株植物滲透調節中的作用可以很簡易地通過直接測量莖或根的滲透壓而容易的驗證,并且可以與它們在組織液中的濃度進行比較。無機離子主要是 K+、Na+、Cl-,無論是在鹽生植物還是在非鹽生雙子葉植物中,這3種離子提供了80%至95%的細胞液滲透壓[31]。K+是循環豐富的離子,同時K+是植物生長的必需元素,在維持細胞的基本功能中扮演了重要角色,并且在保持低水平的蛋白酶和核酸內切酶活性以及防止植物在鹽脅迫下細胞損傷和死亡中的作用也是不言而喻的[32]。有研究表明,堿茅與更敏感的長穗偃麥草(Elytrigiaelongata)相比,在嚴重的高鹽或缺氧條件下,長穗偃麥草將更嚴重地丟失K+,從而證明了K+保留在根中對高鹽缺氧土地耐受性的作用[33]。而對積鹽型鹽生植物而言,對Na+的吸收遠遠大于K+的吸收[34]。絕大部分被植物細胞吸收的Na+并非存在于細胞質中,而是區隔化在液泡中作為廉價的滲透調節物質來維持正常細胞膨壓,從而增強植物抵御鹽逆境的能力[35-37]。研究表明,在介質當中添加適量的NaCl可以增強荒漠植物霸王(Zygophyllumxanthoxylum)的抗旱性,從而促進其生長[38-39];進一步分析表明,干旱脅迫下霸王葉中Na+濃度顯著增加了64%,Na+對葉滲透勢的貢獻由8%增至13%;50 mmol/L NaCl處理使干旱脅迫下霸王葉中Na+顯著增加了2.3倍,Na+對葉滲透勢的貢獻增至28%,從而提高了植株的滲透調節能力[38]。
2.2營養元素平衡
在正常生理狀態下,植物細胞內的離子保持均衡狀態,而在鹽脅迫下,細胞質中過多的離子尤其是Na+對植物細胞的代謝活動會有傷害。大多數植物在鹽脅迫下,組織內的K+含量會降低[4]。王鎖民等[40]在對植物Na+、K+的選擇性吸收及運輸的研究基礎上,首次提出植物根系對土壤中Na+、K+的選擇性吸收(selective absorption,SA)能力以及植株不同部位對Na+、K+選擇性運輸(selective transport,ST)能力的計算公式。研究表明,小花堿茅(Puccinelliatenuiflora)可以在高Na+環境下生存,且其體內維持很低的Na+濃度,主要依靠其對K+/Na+強大的選擇性吸收能力和限制Na+的吸收[25]。這一結果在對鹽敏感和耐鹽品種的長穗偃麥草的比較研究中得到了進一步證實,即耐鹽品種的SA和ST值均顯著大于鹽敏感品種[41]。海濱堿蓬(Suaedamaritima)體內的K+含量變化是隨NaCl濃度的升高而呈升高趨勢,這樣既可以保持一定的K+營養,還可以保持相對穩定的K+/Na+,這對植物本身生長有利[42]。在鹽脅迫的條件下,提供某些微量元素,可有效地提高植物的含水量,促進植物的光合作用,有利于植物生長。研究表明,在含有Na+的土壤中添加硅時,植株的含水量提高了7.3%,同時,植物的光合效率、CO2同化效率等也得到了顯著地提高;在黃瓜(Cucumissativus)[43]、大麥(Hordeumvulgare)[44]、水稻(Oryzasativa)[45-46]、紫花苜蓿(Medicagosativa)[47]、小麥[48]和草地早熟禾(Poapratensis)[49]等植物研究中也得到了同樣的結果。
2.3增強抗氧化脅迫
在正常的生理條件下,植物體內的活性氧自由基和自身的抗氧化系統對活性氧的清除是動態平衡的,可以保持體內正常的代謝過程。在干旱、鹽漬等脅迫下,膜脂過氧化作用加劇,植物體內活性氧含量上升,隨之超氧化物歧化酶、過氧化物酶(peroxidase,POD)、過氧化氫酶和抗壞血酸(ascorbic acid,ASA)等保護酶的活性也相應增加,從而防止膜脂的過氧化作用,以此來增強植物對逆境的耐受性[50]。管博等[51]在鹽地堿蓬(Suaedasalsa)的研究發現,隨著鹽脅迫的增強,SOD作為防御活性氧自由基(reactive oxygen species,ROS)的第一道防線,其活性顯著增加,CAT活性變化趨勢與SOD相似。此發現在對梭梭(Haloxylonammodendron)[52]和溝葉結縷草(Zoysiamatrella)[53]的研究中也得到證實。有研究表明,高鹽和干旱處理下,西藏野生大麥(Hordeumvulgarevar.trifurcatum)體內的ROS水平會顯著升高來抵御傷害[54]。然而在重度鹽脅迫下,植物體內的這些活性氧去除劑的結構發生破壞,植物清除活性氧的防御能力下降,使膜脂的過氧化作用加劇,破壞細胞膜的透性[4,7,55]。
3 高等植物耐鹽相關基因研究進展
隨著分子生物學的發展,人們能夠在基因組成、表達調控及信號轉導等分子水平上認識植物的耐鹽機理;并且已經通過對植物耐鹽相關基因的研究,來進一步提高植物的耐鹽性。目前,高等植物耐鹽相關基因的研究主要集中在離子轉運蛋白基因、滲透調節相關基因、信號傳導相關基因、細胞抗氧化相關基因等。
3.1離子轉運蛋白基因
3.1.1質膜上離子轉運蛋白基因 有研究表明,環核苷酸調控通道(cyclic-nucleotide-gated channel,CNGCs)和非選擇性陽離子通道/電壓非依賴型通道(non-selective cation channel/voltage-independent channel,NSCCs/VICs)是不同類型的通道蛋白,在一些植物中CNGCs涉及Ca2+的信號轉導,而NSCCs/VICs涉及Na+的攝入[56]。而NSCCs/VICs[57]是根部Na+進入細胞膜的通道蛋白,當NSCCs通道被抑制時,就可緩解細胞的鹽脅迫(圖2)。Tyerman和Skerrett[58]在研究小麥根部的NSCCs通道時發現,涌入的Na+主要分布于根部的各個區室及表皮的原生質內。低親和性陽離子轉運體(low-affinity cation transporter,LCT1)是從小麥中發現的一類能夠介導低親和性陽離子吸收的蛋白[59]。研究表明,小麥中的LCT1不僅可以維持Rb+與Na+的濃度的平衡,吸收少量的Ca2+,也可維持Na+/K+的濃度[60];但由于受到土壤中Ca2+濃度的影響,LCT1并不是Na+流入的主要通道[4,61]。
高親和K+轉運載體(high-affinity K+transporter,HKT)是一種與植物耐鹽性密切相關的Na+或Na+-K+轉運蛋白,能將植物木質部中過多的Na+卸載到其周圍薄壁細胞中,降低地上部Na+含量,并維持體內K+穩態平衡[4,62](圖2)。根據在異源表達系統中對Na+和K+運輸的不同,HKT蛋白可被分成兩種[63]:HKT1作用于外部K+缺乏時,主要用于特異性Na+的運輸和介導Na+的吸收[64];HKT2則是有K+-Na+協同轉運蛋白及其同系物的功能[4]。Ren等[65]繪制了一個水稻SKC1基因的QTL圖譜,并發現SKC1基因編碼一種HKT型K+/Na+通道蛋白,在植物鹽脅迫下調節K+/Na+離子平衡,維持細胞內外的滲透壓,從而提高植物耐鹽能力,后來證明SKC1為水稻HKT1;5基因。Laurie等[66]研究表明HKT2基因的低表達可以抑制Na+的進入,從而降低組織內Na+的濃度。作為高親和性Na+轉運的轉運體,HKT轉運蛋白在草類以及其他植物中可能都在控制整體植株中Na+的轉運起重要作用[2]。Kader等[67]在研究水稻時發現,轉運蛋白基因OsHKT1、OsHKT2和OsVHA在鹽脅迫下誘導表達,可以通過調節Na+與K+的比例而降低Na+的濃度。研究表明,SsHKT1在鹽地堿蓬的離子平衡和耐鹽方面有重要作用[68]。
眾多研究表明,高親和K+轉運載體(high-affinity K+transporter,HAK)基因對Na+的吸收也有重要作用[69]。鹽對HvHAK1積累影響的分析結果表明,在轉錄水平上,一個強而短暫NaCl脅迫觸發HvHAK1的上調,說明HvHAK1是一個高鹽應答的快速反應基因。還有研究表明,在鹽逆境脅迫下,HvHAK1 mRNA的積累和對K+攝取的控制是高度相關的[70],對Na+吸收的控制則對維持細胞內的pH重要[71]。研究表明,PhaHAK5、PhaHAK2-n、PhaHAK2-e、PhaHAK2-u等基因均可調節鹽生植物中Na+的濃度[72]。
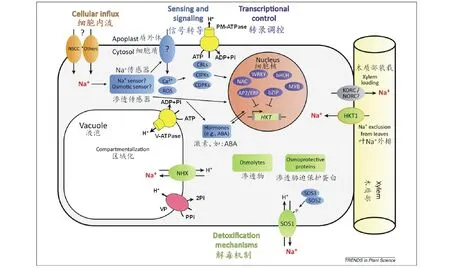
圖2 植物細胞Na+運輸機制及響應鹽逆境的重要調控網絡(在Deinlein等[62]基礎上修訂)
擬南芥K+轉運體(ArabidopsisK+transporter,AKT1)類的通道蛋白是在植物中廣泛表達的一類內整流K+通道。作為一條對NH4+不敏感的K+吸收途徑,AKT1的主要功能是與對NH4+敏感的K+吸收途徑并行存在,使植物能夠適應多樣的外界環境。研究表明,小麥中K+缺乏時,TaAKT1通道會增強對Na+的吸收。眾多研究顯示,OsAKT1的表達并不影響植物中K+的濃度,但是Na+的積累卻依靠此基因的表達[73]。KUP基因家族對細胞內K+濃度的調節有多種類型[74],它們的表達可以表現出對K+的高吸收性,但是在擬南芥(Arabidopsisthaliana)中,NaCl的加入會抑制K+的吸收[75]。Wang等[34]研究表明,積鹽型鹽生植物海濱堿蓬存在兩條低親和性Na+吸收途徑,即在低濃度NaCl(25 mmol/L)條件下主要由HKT介導Na+的吸收,在高濃度NaCl(150 mmol/L)條件下主要由AKT1介導Na+的吸收。Zhang等[76]進一步研究發現兩條途徑的外界Na+濃度介于95~100 mmol/L NaCl之間。
鹽敏感(salt overly sensitive,SOS)基因是從一類超鹽敏感突變體中發現的,遺傳分析表明,這些突變體可分為5種,即sos1、sos2、sos3、sos4和sos5[77-78]。目前篩選出的一些SOS基因家族編碼重要的植物離子載體蛋白、信號轉導蛋白等,它們的存在對提高植物的耐鹽性有重要作用。質膜Na+/H+逆向轉運蛋白就是由SOS1編碼的,該蛋白的主要功能就是將Na+排到細胞外部,從而減少細胞內Na+的積累[62,79-80](圖2)。Guo等[81]研究表明,SOS1在小花堿茅拒Na+當中發揮著重要作用。Ma等[82]對霸王ZxSOS1的研究表明,ZxSOS1參與調控霸王體內Na+、K+轉運和空間分配,進而影響植株生長。Feki等[83]將硬質小麥(Triticumdurum)TdSOS1轉入擬南芥顯著提高了轉基因植物的耐鹽性。Nie等[84]通過將大豆(Glycinemax)GmsSOS1基因轉到缺失SOS1的擬南芥中,證明了其互補性,即驗證了SOS1的功能。SOS2和SOS3都是編碼信號轉導途徑中的蛋白,在信號轉導過程中,彼此有著千絲萬縷的聯系。同時,有研究表明二者功能的行使與Ca2+有密切的關系[85]。目前研究的SOS信號通路就是:高濃度的Na+引起細胞內Ca2+濃度上升,Ca2+與SOS3結合,結合Ca2+后的SOS3與SOS2直接互作而激活了SOS2,之后通過SOS3氨基末端的酰化(amino terminus myristoylation)使SOS3-SOS2復合體會鉚釘到細胞質薄膜上,該復合體中被激活的SOS2磷酸化質膜上的SOS1,從而提高SOS1對Na+的轉運活性而促進高濃度Na+的外排[7]。但在擬南芥和水稻之間SOS信號通路似乎具有保守型,在水稻中,功能性同系物SOS1,SOS2,SOS3也構成一個重要的耐鹽的信號通路[86]。SOS5則在細胞壁形成、細胞伸展、植物繁殖等過程中起重要作用[77]。這一系列研究表明,在鹽脅迫下SOS基因家族對植物的耐鹽性有重要作用。
3.1.2液泡膜上離子轉運蛋白基因 成熟植物細胞的液泡為Na+儲存提供弄了一個很大的空間。將Na+區域化在液泡中是減少細胞溶質之中Na+的有效方式,從而減少胞液中Na+的毒害作用。鈉和氯離子在細胞溶質中的濃度是由穿過質膜和液泡膜的凈通量決定的。液泡膜蛋白中,Na+/H+逆向轉運蛋白(Na+/H+transporters,NHX)參與運輸Na+,該轉運蛋白是由液泡膜H+-ATP酶和H+-焦磷酸酶的作用所形成的質子梯度來驅動的(圖2)[34,62]。NHX基因被證明可以將細胞質中的Na+區域化在液泡中。第一個從擬南芥克隆出的NHX型轉運體基因被命名為AtNHX1[87],它屬于AtNHX1-6亞科六成員之一。這組轉運體運用質子驅動力來運輸Na+和K+[88-89]。除了在耐鹽性中的作用,NHX轉運體還涉及許多其他細胞過程,包括細胞內pH值調節[90],囊泡運輸和蛋白定位[91]。從發現擬南芥中NHX轉運體后,已在許多其他植物中發現了此類型的轉運體,包括水稻OsNHX1[92]、細葉海濱藜(Atriplexgmelini)AgNHX1[93]、大麥HvNHX1[94]、棉花(Gossypiumspp.)GhNHX1[95]、玉米ZmNHX[96]、小麥TaNHX1[97]和TaNHX2[98]、苜蓿MsNHX1[99]、大豆(Glycinemax)GmNHX1[100]、長穗偃麥草(Elytrigiaelongata)AeNHX1[101]、珍珠狼尾草(Pennisetumglaucum)PgNHX1[102]、灰綠藜(Chenopodiumglaucum)CgNHX1[103]、胡楊(Populuseuphratica)PeNHX1-6[104]、鹽穗木(Halostachyscaspica)HcNHX1[105]、霸王ZxNHX1[37]、海蓬子(Salicorniabigelovii)SbNHX1[106]和花花柴(Kareliniacaspia)KcNHX1和KcNHX2[107]等。 Wu等[37]發現在鹽或干旱條件下,霸王ZxNHX的上調與Na+的積累呈正相關關系。進一步研究表明,霸王ZxNHX在控制Na+、K+的吸收,長距離運輸和整株內離子穩態方面具有重要作用[108]。自從報道AtNHX基因超表達可以使擬南芥具有耐鹽性后[87],AtNHX或其他NHX基因在植物中超表達以使之具有抗逆性的成功例子也越來越多。例如將擬南芥AtNHX1分別導入玉米[109]、小麥[110]、棉花[111]和花生(Arachishypogaea)[112],將細葉海濱藜AgNHX1導入水稻[93],將棉花GhNHX1導入煙草[95],將短芒大麥草(Hordeumbrevisubulatum)HbNHX1導入煙草[98],將狼尾草(Pennisetumalopecuroides)PgNHX1導入芥菜(Brassicajuncea)[113]和水稻[102],將長穗冰草(Agropyronelongatum)AeNHX1導入擬南芥和高羊茅(Festucaelata)[101],將擬南芥AtNHX5導入蘭豬耳(Toreniafournieri)[114],將獐茅(Aeluropussinensis)AlNHX導入煙草[115],將小麥TaNHX1導入紫花苜蓿[116],將北美海蓬子(SalicorniaBigelovii)SbNHX1導入麻瘋樹(Jatrophacurcas)[117],將綠豆(Vignaradiata)VrNHX1導入擬南芥[118],均顯著提高了轉基因植物的耐鹽性。
NHX轉運體通過跨液泡膜H+梯度驅動來轉運Na+至液泡,因此通過H+-ATP酶和H+-焦磷酸酶的超表達增加H+驅動力,從而有可能提高抗鹽性。然而,由于其多亞基的特征,液泡H-ATP酶并不是超表達的理想候選者,因為不大可能將所有的亞基在相似的水平過量表達而去創建一個功能性的超表達轉基因植物。液泡膜H+-焦磷酸酶(vacuolar pyrophosphatase,VP)是用于此目的的一個更好的選擇,因為它是一個單一的多肽蛋白[119-120],并且,它產生H+梯度的能力可與H-ATP酶相媲美(圖2)[62,121]。H-焦磷酸酶基因也首先在擬南芥中被克隆出來,命名為AVP1[119]。VP基因也已從其他許多高等植物中克隆得到,如鹽芥TsVP[121]和鹽地堿蓬SsVP等[122]。擬南芥AVP1的過量表達賦予轉基因植物耐旱和耐鹽性,在許多植物中得到了證實,如擬南芥[120,123]、番茄[124]、紫花苜蓿[125]、匍匐翦股穎(Agrostisstolonifera)[126]、大麥[127]、甘蔗(Saccharumofficinarum)[128]。同時,其他VP基因導入植物后,植物抗逆性也得到了提高,例如將鹽地堿蓬SsVP1轉入擬南芥[122],將鹽芥(Thellungiellahalophila)TsVP轉入煙草[129]、棉花[130]和玉米[130],將鹽爪爪KfVP1轉入擬南芥[131],將小麥TaVP1轉入煙草[132],將小麥TaVP基因轉入煙草[133]。
近年來,一些學者將NHX和VP兩種轉運蛋白基因構建雙價表達載體同時超表達進行了轉基因研究,如在水稻中超表達鹽地堿蓬SsNHX1和擬南芥AVP1[134],水稻OsNHX1和OsVP1[135];在擬南芥中超表達小麥TaNHX1和TaTVP1[136];在西紅柿中超表達狼尾草PgNHX1和擬南芥AVP1[137];在煙草中超表達小麥TaNHX1和TaTVP1[138]以及在百脈根(Lotuscorniculatus)中超表達霸王ZxNHX和ZxVP1-1[139]。
3.2滲透調節相關基因
滲透脅迫是植物鹽脅迫的主要方面,滲透調節相關基因在植物耐鹽性中的作用也進行了較多研究。多元醇、脯氨酸、海藻糖、甜菜堿等均是植物耐鹽的重要滲透調節物質,因而合成這些滲透調節物質的一些關鍵基因在耐鹽中起到重要作用。如mtlD基因和gutD基因,它們分別編碼合成甘露醇和山梨醇的酶。超表達mtlD基因增加了甘露醇在毛白楊(Populustomentosa)體內的積累從而增強了其耐鹽性[140];轉gutD基因可使轉基因植株產生山梨醇,轉基因玉米可耐受1.17% NaCl的濃度[141];在水稻中超表達mtlD和gutD基因提高了其耐鹽性[142]。Kishor等[143]對P5CS基因進行研究,獲得P5CS轉基因煙草,其中脯氨酸的含量明顯提高,與對照相比,耐鹽性有提高。晚期胚胎發生富集蛋白(late embriogenesis abundant protein,LEA)是Dure等[144]首次在棉花種子發育晚期胚胎中發現的一類蛋白。Xu 等[145]將LEA基因轉入水稻懸浮細胞中,得到的轉基因植株經過繁殖,在第二代中表現出耐鹽的能力。張寧等[146]從菠菜(Spinaciaoleracea)葉片中分離了甜菜堿醛脫氫酶(betaine-aldehyde dehydrogenase,BADH)基因,并將該基因與其他植物的BADH序列作了同源性分析,同時,證實了菠菜BADH基因的轉錄與表達受干旱和鹽脅迫的誘導。Jia等[147]將山菠菜(Atriplexhortensis)的BADH基因轉入煙草,提高了煙草的耐鹽性。在水稻中過量表達缺失D結構域的組成型活性突變形式OsbZIP46CA1可以顯著增強耐旱性和抵抗滲透脅迫的能力,并可顯著上調已知的逆境應答基因(包括ABF或AREB類成員的下游基因)的表達[148]。
3.3信號傳導相關基因
在鹽脅迫下,植物可以關閉或開啟某些基因的表達,從而維持自身的營養平衡。與植物耐鹽性相關的信號轉導途徑包括:SOS途徑(見3.1.1)、脫落酸(abscisic acid,ABA)信號和蛋白激酶途徑。
ABA應答基因可分為ABA依賴型和ABA非依賴型。目前,已發現多種轉錄因子與鹽脅迫有關,這些基因的表達分為4條途徑:兩條依賴ABA途徑,兩條不依賴ABA途徑[149]。十字花科植物鹽芥是擬南芥的近親,它耐鹽,耐低溫和氧化脅迫,鹽芥基因組相對較小,因此成為研究植物耐受逆境分子機理的理想材料[150]。在鹽芥中研究發現,所有涉及ABA生物合成途徑的基因家族在鹽脅迫下基因數目增加,這種增加會調節復雜的ABA生物合成過程[45,151]。
蛋白激酶在調節植物對非生物脅迫的響應中發揮了重要作用。蛋白激酶是信號轉導的重要元件,信號轉導中絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)途徑是重要途徑之一。目前已分離到多種能被逆境誘導的MAPK基因:擬南芥AtMEKK1、AtMEK1/AtMEK2、ATMPK3、AtMPK4、ANP1、AtMKK2以及煙草NPK1、AtMPK3和AtMPK6等[152-153]。III型亞家族SnRK2蛋白(SnRK2.6/2.3/2.2)是擬南芥脫落酸信號轉導過程中關鍵的正調控因子。這些激酶受脫落酸或滲透脅迫激活后,可磷酸化與脅迫相關的轉錄因子和離子通道,最終使植物免受高鹽的危害[154]。有研究表明,野大麥的CIPK蛋白(CBL-interacting protein kinase,CIPK)HbCIPK2對鹽和滲透脅迫耐受性起正調節作用,HbCIPK2有助于阻止根中K+的丟失和Na+的積累,以便維持K+/Na+的動態平衡和細胞免于死亡[155]。有研究結果表明,PLDα1(磷脂酶)D衍生的磷脂酸(phosphatidic acid,PA)與微管結合蛋白(microtubule-associated protein,MAP)MAP65-1結合后,可調控微管的穩定及對鹽脅迫的耐受性。MAP65-1是PA的一個靶蛋白,該研究揭示了在環境脅迫引起的信號轉導過程中膜脂與細胞骨架之間的功能性聯系[156]。這一研究結果提出了植物中磷脂酸通過調控微管結合蛋白活性參與鹽脅迫反應的新機制。
3.4細胞抗氧化相關基因
膜脂的過氧化導致細胞膜透性的增加是鹽脅迫的主要危害之一。細胞膜本身具有選擇透過性,表面具有多種離子運輸與傳遞的蛋白,內部有多種抗氧化機制,因而可以抵制膜內不飽和脂肪酸的過氧化作用。目前發現,SOD、POD、GSH等是細胞內主要的抗氧化物質。它們本身可抑制氧自由基的產生,清除體內的活性氧。眾多研究表明,植物體內某些基因的存在,對植物的抗氧化有重要作用。如:NtGST/GPX基因編碼的酶既有谷胱苷肽-S-轉移酶活性,又有谷光苷肽過氧化物酶活性,將該基因在煙草中過量表達,可以增強植物的耐鹽性和耐寒性[157]。Kovtun等[158]將一種MAPKKK基因——ANP1基因轉入煙草中發現,它可通過MAPK級聯反應激活谷胱苷肽-S-轉移酶基因(GST6)的表達,從而使植株表現出耐鹽性。S-腺苷甲硫氨酸脫羧酶(S-adenosylmethionine decarboxylase,SAMDC)是多胺合成中的一個關鍵酶,SAMDC基因在煙草中過量表達可以增強植物對鹽和其他非生物脅迫的抗性。維生素C是植物體必需的維生素之一,在光合電子傳遞、活性氧清除、氧化還原平衡調控、細胞分裂與生長、衰老與凋亡等生理過程中具有重要作用。在對擬南芥的研究中表明,擬南芥AtERF98是維生素C合成的關鍵轉錄調控因子,鹽可以誘導AtERF98表達,而AtERF98突變顯著抑制鹽誘導的維生素C合成,并降低清除活性氧的能力,表現為鹽敏感。這些結果表明,ERF類轉錄因子可通過調控抗氧化物質的合成來調節植物對鹽脅迫的應答反應[159]。
4 提高植物耐鹽性的途徑
提高植物耐鹽性的途徑有很多,其中包括抗鹽鍛煉,加入生長調節劑和培育耐鹽新品種等。
4.1抗鹽鍛煉
植物耐鹽能力常隨生長發育時期的不同而異,且對鹽分的抵抗力有一個適應鍛煉過程。種子在一定濃度的鹽溶液中吸水膨脹,然后再播種萌發,可提高作物生育期的耐鹽能力。因此,在對植物進行抗鹽鍛煉時,可逐漸升高Na+濃度,從而提高植物的耐鹽能力。用CaCl2浸種的玉米在鹽脅迫下的葉綠素含量、細胞膜透性和根系活力的變化程度均小于水浸種,脯氨酸含量、干物質重高于水浸種,水勢低于水浸種,提高了三葉期玉米的耐鹽能力[160]。在低濃度鈉鹽處理下,夏枯草(Prunellavulgaris)種子的發芽率、發芽勢、活力指數、根長、苗高以及鮮重都得到了顯著提高[161]。
4.2加入生長調節劑
在很多情況下,植物對逆境脅迫的響應是通過改變內源激素的水平來實現的。如植物處于非生物逆境脅迫條件下會產生大量的ABA和乙烯(ethylene,Eth)等。生長素不僅調控植物的生長發育,也廣泛參與逆境脅迫反應[162]。植物使用不同的策略來應對高鹽土壤,植物可以調整自身的根系結構和根生長的方向來避免局部鹽濃度過高的情況[163]。有一種模式就是通過鹽度影響了植物激素在根系的分布,從而影響根生長。除了植物生長素(indole-3-acetic acid,IAA),細胞分裂素(cytokinin,CTK),乙烯和脫落酸也有相應的作用。植物生長素的運用可以通過促進側根形成從而緩解滲透脅迫并不受ABA的影響,這表明這兩種激素的行為獨立地確定側根原基的命運[164]。用植物激素處理植株,是常用的提高植物耐鹽性的途徑之一。例如,在含0.15% Na2SO4土壤中的小麥生長不良,但在播前用IAA浸種,可以抵消Na2SO4抑制小麥根系生長的作用,使小麥生長良好[165]。ABA是一種逆境激素,能誘導氣孔關閉,減少蒸騰作用、減少鹽的吸收,提高作物的耐鹽能力,它普遍存在于高等植物中,在植物對逆境的適應中有著重要的作用[166]。此外,NO在植物耐受逆境中的作用尤其引人注意。如NO能顯著緩解鹽對鹽地堿蓬種子萌發的抑制,外源NO供體硝普鈉(sodium nitroprusside,SNP)能通過促進種子吸水以緩解鹽的滲透脅迫來緩解鹽脅迫下鹽地堿蓬種子萌發[167]。并且SNP還可以緩解鹽脅迫對蒺藜苜蓿(Medicagotruncatula)種子萌發的抑制作用[168]。
4.3培育耐鹽品種
隨著植物分子生物學研究的深入、植物基因工程的發展、育種學的進步,培育耐鹽的植物品種已經從普通的生理生化等表面的研究深入到了分子領域。現今,人們可以通過改造植物內部的基因、構建耐鹽基因的載體和遺傳轉化等方法進一步改良植株。如上文所述,利用高等植物耐鹽相關基因培育耐鹽作物品種將會成為未來研究的主要方向。此外有研究表明,植物的多倍體化同樣可以使植物具有耐鹽性,同時增強植物鉀的積累[169]。
5 展望
近年來對于作物的耐鹽性研究發展迅速,但在很大程度上仍然停留在通過鑒定和利用擬南芥中已知的相應基因來進行耐鹽性調控的水平。對水稻[64,170]和小麥[171-172]的研究已經指出了一條闡明作物特別是禾谷類作物耐鹽機制的研究方向,但對作物的系統性研究仍然需要使用正向和反向遺傳工具以及植物生理學、生物化學和分子生物學等手段。用正向遺傳篩選作物鹽敏感突變體從而確定作物中重要的耐鹽基因仍然沒有廣泛開展,這種方法將有助于找到新基因或特定的作物耐鹽機制。可以相信,擬南芥中未確定的某些機制,可能存在于作物中,特別是具有不同體系結構和組織構架的單子葉植物組織中。
由3種類型的Na+轉運體調控的3種細胞水平耐鹽機制(控制Na+吸收,增強Na+外排,提高Na+區域化)可以共同協調來提高植物的抗鹽能力。然而,雖然在細胞水平上這3種Na+轉運機制易于理解,但在整體植物中這些轉運體在特定位置上的協調作用機制仍然沒有完全搞清楚。因此,還需要在生理和分子水平上進一步研究來確定這些轉運體的組織和細胞定位以闡明其在抗鹽中的相互協調功能。除此之外,這些轉運體在其他生物過程中的作用也有待于進一步研究。顯而易見的是,這些轉運體不僅僅對植物耐鹽性很重要,而且還涉及其他的細胞生理學過程,這在擬南芥中已經得到證實;然而,除了耐鹽性以外,這些轉運體在作物中的其他作用還不是很清楚。對這些方面研究將會有助于我們估計過量表達這些轉運體來增強植物耐鹽性后所產生的潛在副作用。
鹽生植物為闡明高等植物耐鹽機制的研究提供了天然資源。利用親緣關系密切的鹽生植物和甜土植物以及比較基因組學研究方法將有助于闡明鹽生植物耐鹽性的遺傳基礎。Wu等[150]的研究證明了這一結論,即利用比較基因組學方法來確定鹽芥基因組和擬南芥基因組之間的差異以及其與鹽脅迫反應和抗鹽的關系,最終鑒定其耐鹽的分子機制。雖然比較基因組學的研究需要完整的基因組序列,但可以預見的是,隨著測序技術的迅速發展和相關成本的減少,可供利用的基因組序列將會更加豐富。這種方法也可以用于鑒定自然變異種群中耐鹽變種中的耐鹽基因的變化,這會在分子育種方面為作物的耐鹽性遺傳改良提供有價值的信息。此外,轉錄組學、蛋白質組學和表觀遺傳學等手段將加速鹽生植物耐鹽機制的研究和耐鹽基因的大量挖掘。
[1] Flowers T J.Improving crop salt tolerance.Journal of Experimental Botany, 2004, 55:307-319.
[2] Kronzucker H J, Coskun D, Schulze L M,etal.Sodium as nutrient and toxicant.Plant and Soil, 2013, 369:1-23.
[3] Janz D, Polle A.Harnessing salt for woody biomass production.Tree Physiology, 2012, 32:1-3.
[4] Zhang J L, Flowers T J, Wang S M.Mechanisms of sodium uptake by roots of higher plants.Plant and Soil, 2010, 326:45-60.
[5] Zhao K F, Li F Z, Fan S J,etal.Halophytes in China.Chinese Bulletin of Botany, 1999, 16(3):201-207.
[6] Zhao K F.Plants adapt to salt adversity.Bulletin of Biology, 2002, 37(6):7-10.
[7] Zhang J L, Shi H Z.Physiological and molecular mechanisms of plant salt tolerance.Photosynthesis Research, 2013, 115:1-22.
[8] Munns R, Tester M.Mechanisms of salinity tolerance.Annual Review of Plant Biology, 2008, 59:651-681.
[9] Zhu J K.Plant salt tolerance.Trends in Plant Science, 2001, 6:66-71.
[10] Kronzucker H J, Britto D T.Sodium transport in plants:a critical review.New Phytologist, 2011, 189:54-81.
[11] Gorai M, El A W, Yang X,etal.Toward understanding the ecological role of mucilage in seed germination of a desert shrubHenophytondeserti:interactive effects of temperature, salinity and osmotic stress.Plant and Soil, 2014, 374:727-738.
[12] Wei Y, Dong M, Huang Z Y,etal.Factors influencingseed germination ofSalsolaaffinis(Chenopodiaceae), a dominant annual halophyte inhabiting the deserts of Xinjiang.Flora of China, 2008, 203:134-140.
[13] Li Y, Shen Y Y, Yan S G.Comparative studies of effect of NaCl stress on the seed germination of 5 forage species.Pratacultural Science, 1997, 14(2):50-53.
[14] Liang Y C.Effects of silicon on enzyme activity and sodium, potassium and calcium concentration in barley under salt stress.Plant and Soil, 1999, 209:217-224.
[15] Trono D, Flagella Z, Laus M N,etal.The uncoupling protein and the potassium channel are activated by hyperosmotic stress in mitochondria from durum wheat seedlings.Plant Cell and Environment, 2004, 27:437-448.
[16] Becker D, Hoth S, Ache P,etal.Regulation of the ABA-sensitiveArabidopsispotassium channel gene GORK in response to water stress.Febs Letters, 2003, 554:119-126.
[17] Ottow E A, Brinker M, Teichmann T,etal.Populuseuphraticadisplays apoplastic sodium accumulation, osmotic adjustment by decreases in calcium and soluble carbohydrates, and develops leaf succulence under salt stress.Plant Physiology, 2005, 139:1762-1772.
[18] Song J, Ding X D, Feng G,etal.Nutritional and osmotic roles of nitrate in a euhalophyte and a xerophyte in saline conditions.New Phytologist, 2006, 171:357-366.
[19] Liu J, Cai H, Liu Y,etal.A study on physiological characteristics and cmparison of salt resistance of twoMedicagosativaat the seeding stage.Acta Prataculturae Sinica, 2013, 22(2):250-256.
[20] Jarunee J, Kenjiusui, Hiroshi M.Differences in physiological responses to NaCl between salt-tolerantSesbaniarostrataBrem.and Obem.And non-tolerantPhaseolusvulgarisL.Weed Biology and Management, 2003, 3:21-27.
[21] Makela P, Karkkainen J, Somersalo S.Effect of glycinebetaine on chloroplast ultrastructure, chlorophyll and protein content, and RuBPCO activities in tomato grown under drought or salinity.Biologia Plantarum, 2000, 43:471-475.
[22] Leshem Y, Melamed B N, Cagnac O,etal.Suppression ofArabidopsisvesicle-SNARE expression inhibited fusion of H2O2containing vesicles with tonoplast and increased salt tolerance.Proceedings of the National Academy of Sciences of the United States of America, 2006, 103:18008-18013.
[23] Flowers T J, Colmer T D.Salinity tolerance in halophytes.New Phytologist, 2008, 179:945-963.
[24] Shabala S, Bose J, Hedrich R.Salt bladders:do they matter.Trends in Plant Science, 2014, 19(11):687-691.
[25] Wang C M, Zhang J L, Liu X S,etal.Puccinelliatenuifloramaintains a low Na+level under salinity by limiting unidirectional Na+influx resulting in a high selectivity for K+over Na+.Plant, Cell and Environment, 2009, 32:486-496.
[26] Ueda A, Yamamoto-Yamane Y, Takabe T.Salt stress enhances proline utilization in the apical region of barley roots.Biochemical and Biophysical Research Communications, 2007, 355:61-66.
[27] Wang C Q, Zhao J Q, Chen M,etal.Identification of betacyanin and effects of environmental factors on its accumulation in halophyteSuaedasalsa.Journal of Plant Physiology and Molecular Biology, 2006, 32(2):195-201.
[28] Md A H, Mst N A B, Yoshimasa N,etal.Proline and glycinebetaine enhance antioxidant defense and methylglyoxal detoxification systems and reduce NaCl-induced damage in cultured tobacco cells.Journal of Plant Physiology, 2008, 165:813-824.
[29] Incharoensakd A, Takabe T, A kazawa T.Effect of bteaine on enzyme activity and subunit internation of ribulose-1,5 -bisphosphate carboxylase/oxygenas fromAphnothecehalophytica.Plant Physiology, 1986, 81:1044-1049.
[30] Dubey R S, Singh A K.Salinity induces accumulation of soluble sugars and alters the activity of sugar metabolising enzymes in rice plants.Biologia Plantarum, 1999, 42:233-239.
[31] Shabala S, Shabala L.Ion transport and osmotic adjustment in plants and bacteria.Biomolecular Concepts, 2011, 2:407-419.
[32] Demidchik V, Cuin T A, Svistunenko D,etal.Arabidopsisroot K+-efflux conductance activated by hydroxyl radicals:single-channel properties, genetic basis and involvement in stress-induced cell death.Journal of Cell Science, 2010, 123:1468-1479.
[33] Teaklea N L, Bazihizina N, Shabala S,etal.Differential tolerance to combined salinity and O2deficiency in the halophytic grassesPuccinelliaciliataandThinopyrumponticum:The importance of K+retention in roots.Environmental and Experimental Botany, 2013, 87:69-78.
[34] Wang S M, Zhang J L, Flowers T J.Low-affinity Na+uptake in the halophyteSuaedamaritima.Plant Physiology, 2007, 145(2):559-571.
[35] Cuin T A, Bose J, Stefano G,etal.Assessing the role of root plasma membrane and tonoplast Na+/H+exchangers in salinity tolerance in wheat:in planta quantification methods.Plant, Cell and Environment, 2009, 34:947-961.
[36] Schmidt U G, Endler A, Schelbert S,etal.Novel tonoplast transporters identified using a proteomic approach with vacuoles isolated from cauliflower buds.Plant Physiology, 2007, 145:216-229.
[37] Wu G Q, Xi J J, Wang Q,etal.TheZxNHXgene encoding tonoplast Na+/H+antiporter from the xerophyteZygophyllumxanthoxylumplays important roles in response to salt and drought.Journal of Plant Physiology, 2011, 168:758-767.
[38] Ma Q, Yue L J, Zhang J L,etal.Sodium chloride improves photosynthesis and water status in the succulent xerophyteZygophyllumxanthoxylum.Tree Physiology, 2012, 32(1):4-13.
[39] Yue L J, Li S X, Ma Q,etal.NaCl stimulates growth and alleviates water stress in the xerophyteZygophyllumxanthoxylum.Journal of Arid Environments, 2012, 87:153-160.
[40] Wang S M, Zhu X Y, Shu X X.Studies on the characteristics of ion absorption anddistribution inPuccinelliatenuiflora.Acta Prataculturae Sinica, 1994, 3(1):39-43.
[41] Guo Q, Meng L, Mao P C,etal.Salt tolerance in two tall wheatgrass species is associated with selective capacity for K+over Na+.Acta Physiologiae Plantarum, 2014, 37:1708.
[42] Zhang J L, Flowers T J, Wang S M.Differentiation of low-affinity Na+uptake pathways and kinetics of the effects of K+on Na+uptake in the halophyteSuaedamaritime.Plant and Soil, 2013, 368(1-2):629-640.
[43] Zhu Z J, Wei G Q, Li J,etal.Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (CucumissativusL.).Plant Science, 2004, 167:527-533.
[44] Liang Y C, Zhang W H, Chen Q,etal.Effects of silicon on H+-ATPase and H+-PPase activity, fatty acid composition and fluidity of tonoplast vesicles from roots of salt-stressed barley (HordeumvulgareL.).Environmental and Experimental Botany, 2005, 53:29-37.
[45] Gong H J, Randall D P, Flowers T J.Silicon deposition in the root reduces sodium uptake in rice (OryzasativaL.) seedlings by reducing bypass flow.Plant Cell and Environment, 2006, 29:1970-1979.
[46] Ma J F, Yamaji N, Mitani N,etal.An efflux transporter of silicon in rice.Nature, 2007, 448:209-212.
[47] Wang X S, Han J G.Effects of NaCl and silicon on ion distribution in the roots, shoots and leaves of two alfalfa cultivars with different salt tolerance.Soil Science and Plant Nutrition, 2007, 53:278-285.
[48] Tuna A L, Kaya C, Higgs D,etal.Silicon improves salinity tolerance in wheat plants.Environmental and Experimental Botany, 2008, 62:10-16.
[49] Chai Q, Shao X, Zhang J.Silicon effects onPoapratensisresponses to salinity.HortScience, 2010, 45:1876-1881.
[50] Bose J, Rodrigo-Moreno A, Shabala S.ROS homeostasis in halophytes in the context of salinity stress tolerance.Journal of Experimental Botany, 2014, 65(5):1241-1257.
[51] Guan B, Yv J B, Lu Z H,etal.Effects of water-salt stresses on seeding growth and activities of antioxidative enzyme ofSuaedasalsain coastal wetlands of the yellow river delta.Environmental Science, 2011, 32(8):2422-2429.
[52] Lu Y, Lei J Q, Zeng F J,etal.Effects of salt treatments on the growth and ecophysiological characteristies.Acta Prataculturae Sinica, 2014, 23(3):152-159.
[53] Xue X D, Dong X Y, Duan Y X,etal.A comparison of salt resistance of three kinds ofZoysiaat different salt concentrations.Acta Prataculturae Sinica, 2013, 22(6):315-320.
[54] Ahmed I M, Nadira U A, Bibi N,etal.Secondary metabolism and antioxidants are involved in the tolerance to drought and salinity, separately and combined, in Tibetan wild barley.Environmental and Experimental Botany, 2015, 111:1-12.
[55] Gao H J, Yang H Y, Bai J P,etal.Ultrastructural and physiological responses of potato (SolanumtuberosumL.) plantlets to gradient saline stress.Frontiers in Plant Science, 2014, 5:787.
[56] Talke I N, Blaudez D, Maathuis F J M,etal.CNGCs:prime targets of plant cyclic nucleotide signalling.Trends in Plant Science, 2003, 8:286-293.
[57] Amtmann A, Sanders D.Mechanism of Na+uptake by plant cells.Advances in Botanical Research, 1999, 29:75-112.
[58] Tyerman S D, Skerrett I M.Root ion channels and salinity.Scientia Horticulturae, 1999, 78:175-235.
[59] Zhang H F, Wang S M.Advances in study of Na+uptake and transport in higher plants and Na+homeostasis in the cell.Chinese Bulletin of Botany, 2007, 24(5):561-571.
[60] Amtmann A, Fischer M, Marsh E L,etal.The wheat cDNA LCT1 generates hypersensitivity to sodium in a salt-sensitive yeast strain.Plant Physiology, 2001, 126:1061-1071.
[61] Kronzucker H J, Szczerba M W, Schulze L M,etal.Non-reciprocal interactions between K+and Na+ions in barley (HordeumvulgareL.).Journal of Experimental Botany, 2008, 59:2793-2801.
[62] Deinlein U, Stephan A B, Horie T,etal.Plant salt-tolerance mechanisms.Trends in Plant Science, 2014, 19(6):371-379.
[63] Zamani B M, Ebrahimie E, Niazi A.In silico analysis of high affinity potassium transporter (HKT) isoforms in different plants.Aquatic Biosystems, 2014, 10:9.
[64] Wang Q, Guan C, Wang P,etal.AtHKT1;1 andAtHAK5 mediate low-affinity Na+uptake inArabidopsisthalianaunder mild salt stress.Plant Growth Regulation, 2015, 75(3):615-623.
[65] Ren Z H, Gao J P, Li L G,etal.A rice quantitative trait locus for salt tolerance encodes a sodium transporter.Nature Genetics, 2005, 37:1141-1146.
[66] Laurie S, Feeney K A, Maathuis F J M,etal.A role forHKT1 in sodium uptake by wheat roots.Plant Journal, 2002, 32:139-149.
[67] Kader M A, Seidel T, Golldack D,etal.Expressions ofOsHKT1,OsHKT2, andOsVHAare differentially regulated under NaCl stress in salt-sensitive and salt-tolerant rice (OryzasativaL.) cultivars.Journal of Experimental Botany, 2006, 57(15):4257-4268.
[68] Shao Q, Zhao C, Han N,etal.Cloning and expression pattern ofSsHKT1 encoding a putative cation transporter from halophyteSuaedasalsa.DNA sequence, 2008, 19(2):106-114.
[69] Senn M E, Rubio F, Banuelos M A,etal.Comparative functional features of plant potassiumHvHAK1 andHvHAK2 transporters.Journal of Biological Chemistry, 2001, 30:44563-44569.
[70] Fulgenzi F R, Peralta M L, Mangano S,etal.The ionic environment controls the contribution of the barleyHvHAK1 transporter to potassium acquisition.Plant Physiology, 2008, 147:252-262.
[71] Carden D E, Walker D J, Flowers T J,etal.Single-cell measurements of the contributions of cytosolic Na+and K+to salt tolerance.Plant Physiology, 2003, 131:676-683.
[72] Takahashi R, Nishio T, Ichizen N,etal.Cloning and functional analysis of the K+transporter PhaHAK2 from salt-sensitive and salt-tolerant reed plants.Biotechnol Letters, 2007, 29:501-506.
[73] Golldack D, Quigley F, Michalowski C B,etal.Salinity stress-tolerant and -sensitive rice (OryzasativaL.) regulate AKT1-type potassium channel transcripts differently.Plant Molecular Biology, 2003, 51:71-81.
[74] Kim E J, Kwak J M, Uozumi N,etal.AtKUP1:AnArabidopsisgene encoding high-affinity potassium transport activity.Plant Cell, 1998, 10:51-62.
[75] Fu H H, Luan S.AtKUP1:a dual-affinity K+transporter fromArabidopsis.Plant Cell, 1998, 10:63-73.
[76] Zhang J L.Low-Affinity Na+Uptake and Accumulation in the Halophyte Suaeda maritina[D].Lanzhou: Lanzhou University, 2008.
[77] Shi H, Kim Y, Guo Y,etal.TheArabidopsisSOS5 locus encodes a putative cell surface adhesion protein and is required for normal cell expansion.Plant Cell, 2003, 15(1):19-32.
[78] Shi H Z, Quintero F J, Pardo J M,etal.The putative plasma membrane Na+/H+antiporterSOS1 controls long-distance Na+transport in plants.Plant Cell, 2002, 14:465-477.
[79] Wu G Q, Wang P, Ma Q,etal.Selective transport capacity for K+over Na+is linked to the expression levels ofPtSOS1 in halophytePuccinelliatenuiflora.Functional Plant Biology, 2012, 39:1047-1057.
[80] Liu M, Wang T Z, Zhang W H.Sodium extrusion associated with enhanced expression ofSOS1 underlies different salt tolerance betweenMedicagofalcataandMedicagotruncatulaseedlings.Environmental and Experimental Botany, 2015, 110:46-55.
[81] Guo Q, Wang P, Ma Q,etal.Selective transport capacity for K+over Na+is linked to the expression levels ofPtSOS1 in halophytePuccinelliatenuiflora.Functional Plant Biology, 2012, 39:1047-1057.
[82] Ma Q, Li Y X, Yuan H J,etal.ZxSOS1 is essential for long-distance transport and spatial distribution of Na+and K+in the xerophyteZygophyllumxanthoxylum.Plant and Soil, 2014, 374:661-676.
[83] Feki K, Quintero F J, Khoudi H,etal.A constitutively active form of a durum wheat Na+/H+antiporterSOS1 confers high salt tolerance to transgenicArabidopsis.Plant Cell Reports, 2014, 33(2):277-288.
[84] Nie W X, Xu L, Yu B J.A putative soybeanGmsSOS1 confers enhanced salt tolerance to transgenicArabidopsissos1-1 mutant.Protoplasma, 2015, 252(1):127-134.
[85] Ishitani M, Liu J, Halfter U,etal.SOS3 function in plant salt tolerance requires N-myristoylation and calcium binding.Plant Cell, 2000, 12(9):1667-1678.
[86] Martinez-Atienza J, Jiang X, Garciadeblas B,etal.Conservation of the salt overly sensitive pathway in rice.Plant Physiology, 2007, 143:1001-1012.
[87] Gaxiola R A, Rao R, Sherman A,etal.TheArabidopsisthalianaproton transporters, AtNHX1 and AVP1, can function in cation detoxification in yeast.Proceedings of the National Academy of Sciences of the United States of America, 1999, 96:1480-1485.
[88] Apse M P, Aharon G S, Snedden W A,etal.Salt tolerance confermi by over expression of a vacuolar NaCMC antiport inArabidopsis.Science, 1999, 285(12):1256-1258.
[89] Venema K, Quintero F J, Pardo J M,etal.TheArabidopsisNa+/H+exchanger catalyzes low affinity Na+and K+transport in reconstituted vesicles.Journal of Biological Chemistry, 2002, 277:2413-2418.
[90] Yamaguchi T, Fukuda-Tanaka S, Inagaki Y,etal.Genes encoding the vacuolar Na+/H+exchanger and flower coloration.Plant Cell Physiology, 2001, 142:451-461.
[91] Sottosanto J B, Gelli A, Blumwald E.DNA array analyses ofArabidopsisthalianalacking a vacuolar Na+/H+antiporter:impact of AtNHX1 on gene expression.Plant Journal, 2004, 40:752-771.
[92] Fukuda A, Nakamura A, Tanaka Y.Molecular cloning and expression of the Na+/H+exchanger gene inOryzasativa.BBA-Gene Structure and Expression, 1999, 1446:149-155.
[93] Ohta M, Hayashi Y, Nakashima A,etal.Introduction of a Na+/H+antiporter gene fromAtriplexgmeliniconfers salt tolerance to rice.FEBS Letters, 2002, 532:279-282.
[94] Fukuda A, Chiba K, Maeda M,etal.Effect of salt and osmotic stresses on the expression of genes for the vacuolar H+-pyrophosphatase, H+-ATPase subunitA, and Na+/H+antiporter from barley.Journal of Experimental Botany, 2004, 55:585-594.
[95] Wu C A, Yang G D, Meng Q W,etal.The cottonGhNHX1 gene encoding a novel putative tonoplast Na+/H+antiporter plays an important role in salt stress.Plant Cell Physiology, 2004, 45:600-607.
[96] Zorb C, Noll A, Karl S,etal.Molecular characterization of Na+/H+antiporters (ZmNHX) of maize (ZeamaysL.) and their expression under salt stress.Journal of Plant Physiology, 2005, 162:55-66.
[97] Brini F, Gaxiola R A, Berkowitz G A,etal.Cloning and characterization of a wheat vacuolar cation/proton antiporter and pyrophosphatase proton pump.Plant Physiology and Biochemistry, 2005, 43:347-354.
[98] Yu J N, Huang J, Wang Z M,etal.An Na+/H+antiporter gene from wheat plays an important role in stress tolerance.Journal of Biosciences, 2007, 32:1153-1161.
[99] Yang Q C, Wu M S, Wang P Q,etal.Cloning and expression analysis of a vacuolar Na+/H+antiporter gene from alfalfa.DNA sequece, 2005, 16:352-357.
[100] Li W Y, Wong F L, Tsai S N,etal.Tonoplast-locatedGmCLC1 andGmNHX1 from soybean enhance NaCl tolerance in transgenic bright yellow (BY)-2cells.Plant, Cell and Environment, 2006, 29:1122-1137.
[101] Qiao W H, Zhao X Y, Li W,etal.Overexpression ofAeNHX1, a root-specific vacuolar Na+/H+antiporter fromAgropyronelongatum, confers salt tolerance toArabidopsisandFestucaplants.Plant Cell Reports, 2007, 26:1663-1672.
[102] Verma D, Singla-Pareek S L, Rajagopal D,etal.Functional validation of a novel isoform of Na+/H+antiporter fromPennisetumglaucumfor enhancing salinity tolerance in rice.Journal of Biosciences, 2007, 32:621-628.
[103] Li J Y, He X W, Xu L,etal.Molecular and functional comparisons of the vacuolar Na+/H+exchangers originated from glycophytic and halophytic species.Journal of Zhejiang University Science, 2008, 9:132-140.
[104] Ye C Y, Zhang H C, Chen J H,etal.Molecular characterization of putative vacuolar NHX-type Na+/H+exchanger genes from the salt-resistant treePopuluseuphratica.Physiologia Plantarum, 2009, 137:166-174.
[105] Guan B, Hu Y, Zeng Y,etal.Molecular characterization and functional analysis of a vacuolar Na+/H+antiporter gene (HcNHX1) fromHalostachyscaspica.Molecular Biology Reports, 2010, 38:1889-1899.
[106] Jha A, Joshi M, Yadav N,etal.Cloning and characterization of theSalicorniabrachiataNa+/H+antiporter geneSbNHX1 and its expression by abiotic stress.Molecular Biology Reports, 2011, 38:1965-1973.
[107] Liu L, Zeng Y, Pan X,etal.Isolation, molecular characterization, and functional analysis of the vacuolar Na+/ H+antiporter genes from the halophyteKareliniacaspica.Molecular Biology Reports, 2012, 39:7193-7202.
[108] Yuan H J, Ma Q, Wu G Q,etal.ZxNHXcontrols Na+and K+homeostasis at the whole-plant level inZygophyllumxanthoxylumthrough feedbackregulation of the expression of genes involved in their transport.Annals of Botany, 2015, 115(3):495-507.
[109] Yin X Y, Yang A F, Zhang K W,etal.Production and analysis of transgenic maize with improved salt tolerance by the introduction ofAtNHX1 gene.Acta Botanica Sinica, 2004, 46:854-861.
[110] Xue Z Y, Zhi D Y, Xue G,etal.Enhanced salt tolerance of transgenic wheat (TritivumaestivumL.) expressing a vacuolar Na+/H+antiporter gene with improved grain yields in saline soils in the field and a reduced level of leaf Na+.Plant Science, 2004, 167:849-859.
[111] He C, Yan J, Shen G,etal.Expression of anArabidopsisvacuolar sodium/proton antiporter gene in cotton improves photosynthetic performance under salt conditions and increases fiber yield in the field.Plant Cell Physiology, 2005, 46:1848-1854.
[112] Banjara M, Zhu L, Shen G,etal.Expression of anArabidopsissodium/proton antiporter gene (AtNHX1) in peanut to improve salt tolerance.Plant Biotechnology Reports, 2012, 6:59-67.
[113] Rajagopal D, Agarwal P, Tyagi W,etal.PennisetumglaucumNa+/H+antiporter confers high level of salinity tolerance in transgenicBrassicaJuncea.Molecular Breeding, 2007, 19:137-151.
[114] Shi L Y, Li H Q, Pan X P,etal.Improvement ofToreniafournierisalinity tolerance by expression ofArabidopsisAtNHX5.Functional Plant Biology, 2008, 35:185-192.
[115] Zhang G H, Su Q, An L J,etal.Characterization and expression of a vacuolar Na+/H+antiporter gene from the monocot halophyteAeluropuslittoralis.Plant Physiology and Biochemistry, 2008, 46:117-126.
[116] Zhang Y M, Liu Z H, Wen Z Y,etal.The vacuolar Na+-H+antiport geneTaNHX2 confers salt tolerance on transgenic alfalfa (Medicagosativa).Functional Plant Biology, 2012, 39:708-716.
[117] Joshi M, Jha A, Mishra A,etal.Developing transgenic Jatropha using theSbNHX1 gene from an extreme halophyte for cultivation in saline wasteland.PLoS One, 2013, 8(8):e71136.
[118] Mishra S, Alavilli H, Lee B H,etal.Cloning and functional characterization of a vacuolar Na+/H+antiporter gene from mungbean (VrNHX1) and its ectopic expression enhanced salt tolerance inArabidopsisthaliana.PLoS One, 2014, 9(10):e106678.
[119] Sarafian V, Kim Y, Poole R J,etal.Molecular cloning and sequence of cDNA encoding the pyrophosphate-energized vacuolar membrance proton pump ofArabidopsisthaliana.Proceedings of the National Academy of Sciences of the United States of America, 1992, 89:1775-1779.
[120] Gaxiola R A, Palmgren M G, Schumacher K.Plant proton pumps.FEBS Letters, 2007, 581:2204-2214.
[121] Gao F, Gao Q, Duan X G,etal.Cloning of an H+-PPase gene fromThellungiellahalophilaand its heterologous expression to improve tobacco salt tolerance.Journal of Experimental Botany, 2006, 57:3259-3270.
[122] Guo S L, Yin H B, Zhang X,etal.Molecular cloning and characterization of a vacuolar H+-pyrophosphatase gene, SsVP, from the halophyteSuaedasalsaand its overexpression increases salt and drought tolerance ofArabidopsis.Plant Molecular Biology, 2006, 60:41-50.
[123] Li J, Yang H, Peer W A,etal.ArabidopsisH+-PPaseAVP1 regulates auxin-mediated organ development.Science, 2005, 310:121-125.
[124] Park S, Li J, Pittman J K,etal.Up-regulation of a H+- pyrophosphatase (H+-PPase) as a strategy to engineer drought-resistant crop plants.Proceedings of the National Academy of Sciences of the United States of America, 2005, 102:18830-18835.
[125] Bao A K, Wang S M, Wu G Q,etal.Overexpression of theArabidopsisH+-PPase enhanced resistance to salt and drought stress in transgenic alfalfa (MedicagosativaL.).Plant Science, 2009, 176:232-240.
[126] Li Z G, Baldwin M, Hu Q,etal.Heterologous expression ofArabidopsisH+-pyrophosphatase enhances salt tolerance in transgenic creeping bentgrass (AgrostisstoloniferaL.).Plant Cell Environment, 2010, 33:272-289.
[127] Schilling R K, Marschner P, Shavrukov Y,etal.Expression of theArabidopsisvacuolar H+-pyrophosphatase gene (AVP1) improves the shoot biomass of transgenic barley and increases grain yield in a saline field.Plant Biotechnology Journal, 2014, 12(3):378-386.
[128] Kumar T, Uzma, Khan M R,etal.Genetic improvement of sugarcane for drought and salinity stress tolerance usingArabidopsisvacuolarpyrophosphatase (AVP1) gene.Molecular Biotechnology, 2014, 56(3):199-209.
[129] Lv S L, Lian L J, Tao P L,etal.Overexpression ofThellungiellahalophilaH+-PPase (TsVP) in cotton enhances drought stress resistance of plants.Planta, 2009, 229:899-910.
[130] Pei L, Wang J, Li K,etal.Overexpression ofThellungiellahalophilaH+-pyrophosphatase gene improves low phosphate tolerance in maize.PLOS One, 2012, 7(8):e43501.
[131] Yao M, Zeng Y, Liu L,etal.Overexpression of the halophyteKalidiumfoliatumH+-pyrophosphatase gene confers salt and drought tolerance inArabidopsisthaliana.Molecular Biology Reports, 2012, 39:7989-7996.
[132] Khoudi H, Maatar Y, Gouiaa S,etal.Transgenic tobacco plants expressing ectopically wheat H+-pyrophosphatase (H+-PPase) geneTaVP1 show enhanced accumulation and tolerance to cadmium.Journal of Plant Physiology, 2012, 169:98-103.
[133] Li X, Guo C, Gu J,etal.Overexpression of VP, a vacuolar H+-pyrophosphatase gene in wheat (TriticumaestivumL.), improves tobacco plant growth under Pi and N deprivation, high salinity, and drought.The Journal of Experimental Botany, 2014, 65(2):683-696.
[134] Zhao F Y, Zhang X J, Li P H,etal.Co-expression of theSuaedasalsaSsNHX1 andArabidopsisAVP1 confer greater salt tolerance to transgenic rice than the singleSsNHX1.Molecular Breeding, 2006, 17:341-353.
[135] Liu S P, Zheng L Q, Xue Y H,etal.Overexpression ofOsVP1 andOsNHX1 increases tolerance to drought and salinity in rice.Journal of Integrative Plant Biology, 2010, 53:444-452.
[136] Brini F, Hanin M, Mezghani I,etal.Overexpression of wheat Na+/H+antiporterTNHX1 and H+- pyrophosphataseTVP1 improve salt- and drought-stress tolerance inArabidopsisthalianaplants.Journal of Experimental Botany, 2007, 58:301-308.
[137] Bhaskaran S, Savithramma D L.Co-expression ofPennisetumglaucumvacuolar Na+/H+antiporter andArabidopsisH+- pyrophosphatase enhances salt tolerance in transgenic tomato.Journal of Experimental Botany, 2011, 62:5561-5570.
[138] Gouiaa S, Khoudi H, Leidi E O,etal.Expression of wheat Na+/H+antiporterTNHXS1 and H+- pyrophosphataseTVP1 genes in tobacco from a bicistronictranscriptional unit improves salt tolerance.Plant Molecular Biology, 2012, 79:137-155.
[139] Bao A K, Wang Y W, Xi J J,etal.Co-expression of xerophyteZygophyllumxanthoxylumZxNHXandZxVP1-1 enhances salt and drought tolerance in transgenicLotuscorniculatusby increasing cations accumulation.Functional Plant Biology, 2014, 41:203-214.
[140] Hu L, Lu H, Liu Q L,etal.Overexpression ofmtlDgene in transgenicPopulustomentosaimproves salt tolerance through accumulation of mannitol.Tree Physiology, 2005, 25:1273-1281.
[141] Liu Y, Wang G Y, Liu J J,etal.Transfer ofE.coligutDgene into maize and regeneration of salt-tolerant transgenic plants.Science in China Series C-Life Science, 1999, 42(1):90-95.
[142] Wang H Z, Huang D N, Lu R F,etal.Salt tolerance of transgenic rice (OryzasativaL.) withmtlDgene andgutDgene.Chinese Science Bulletin, 2000, 45:1685-1690.
[143] Kishor P B K, Hong Z, Mian G H.Over expression of △’-pyrroline-5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants.Plant Physiology, 1995, 108:1387-1394.
[144] Dure L, Greenway S C, Galau G A.Developmental biochemistry of cottonseed embryogenesis and germination-changing messenger ribonucleic-acid populations as shown byinvitroandinvivoprotein-synthesis.Biochemistry, 1981, 20(14):4162-4168.
[145] Xu D, Duan X, Wang B,etal.Expression of a late embryogenesis abundant protein geneHVA1, from barley confers tolerance to water deficit and salt stress in transgenic rice.Plant Physiology, 1996, 110:249-257.
[146] Zhang N, Wang D, Si H J.Isolation and induced expression of betaine aldehyde dehydrogenase genefrom spinach.Journal of Agricultural Biotechnology, 2004, 12(5):612-613.
[147] Jia G X, Zhu Z Q, Chang F Q,etal.Transformation of tomato with theBADHgene fromAtripleximproves salt tolerance.Plant Cell Reports, 2002, 21(2):141-146.
[148] Tang N, Zhang H, Li X H,etal.Constitutive activation of transcription factor OsbZIP46 improves drought tolerance in rice.Plant Physiology, 2012, 158:1755-1768.
[149] Shinozaki K, Yamaguchi-Shinozaki K.Gene expression and signal transduction in water stress response.Plant Physiology, 1997, 115:327-334.
[150] Wu H J, Zhang Z H, Wang J Y,etal.Insights into salt tolerance from the genome ofThellungiellasalsuginea.Proceedings of the National Academy of Sciences, 2012, 109(30):12219-12224.
[151] Taji T, Seki M, Satou M,etal.Comparative genomics in salt tolerance betweenArabidopsisand a Rabidopsis-related halophyte salt cress usingArabidopsismicroarray.Plant Physiology, 2004, 135:1697-1709.
[152] Sheen J.Signal transduction in maize andArabidopsismesophyll protoplasts.Plant Physiology, 2001, 127:1466-1475.
[153] Moon H, Lee B, Choi G,etal.NDP kinase 2 interacts with two oxidative stress-activated MAPKs to regulate cellular reduxstate and enhances multiple stress tolerance in transgenic plants.Proceedings of the National Academy of Sciences, 2003, 100(1):358-363.
[154] Xie T, Ren R, Zhang Y Y,etal.Molecular mechanism for inhibition of a critical component in theArabidopsisthalianaabscisic acid signal transduction pathways, SnRK2.6, by protein phosphataseABI1.Journal of Biological Chemistry, 2012, 287:794-802.
[155] Li R F, Zhang J W, Wu G Y,etal.HbCIPK2, a novel CBL-interacting protein kinase from halophyteHordeumbrevisubulatum, confers salt and osmotic stress tolerance.Plant, Cell and Environment, 2012, 35:1582-1600.
[156] Zhang Q, Lin F, Mao T,etal.Phosphatidic acid regulates microtubule organization by interacting with MAP65-1 in response to salt stress inArabidopsis.Plant Cell, 2012, 24:4555-4576.
[157] Roxas V P, Lodhi S A, Garrett D K,etal.Stress tolerance in transgenic tobacco seedlings that overexpress glutathione S-transferase/glutathione peroxidase.Plant & Cell Physiology, 2000, 41(11):1229-1234.
[158] Kovtun Y, Chiu W L, Tena G,etal.Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants.Proceedings of the National Academy of Sciences of the United States of America, 2000, 97:2940-2945.
[159] Zhang Z, Wang J, Zhang R X,etal.The ethylene response factor AtERF98 enhances tolerance to salt through the transcriptional activation of ascorbic acid synthesis inArabidopsis.The Plant Journal, 2012, 71:273-287.
[160] Ge Y, Gao P, Xia J Z,etal.The effects of calcium chloride on improving te salt resistance ofZeamaysL.Journal of Northeast Agicultural University, 2004, 35(3):281-284.
[161] Zhang L X, Chang Q S, Hou X G,etal.Effects of sodium salt stress on seed germination ofPrunellavulgaris.Acta Prataculturae Sinica, 2015, 24(3):177-186.
[162] Qian Q, Qu L J, Yuan M,etal.Research advances on plant science in China in 2012.Chinese Bulletin of Botany, 2013, 48:231-287.
[163] Galvan-Ampudia C S, Testerink C.Salt stress signals shape the plant root.Current Opinion in Plant Biology, 2011, 14:296-302.
[164] Brady S M, Sarkar S F, Bonetta D,etal.The abscisic acid insensitive 3(ABI3) gene is modulated by farnesylation and is involved in auxin signaling and lateral root development inArabidopsis.Plant Journal, 2003, 34:67-75.
[165] An J P, Chen K S.The relations between the injury of plasma membrane and the increase of aba content in wheat leaves.Journal of Lanzhou University (Natural Science), 1994, 3:127-131.
[166] Iraudat J, Parcy F, Gosti F.Current advances in abseisic acid action and signaling.Plant Molecular Biology, 1994, 26:1557-1577.
[167] Yuan F, Yang J C, Chen M,etal.Effect of no donor sodium nitroprusside (SNP) on seed germination ofSuaedasalsaL.under NaCl Stress.Plant Physiology Journa, 2010, 46(1):24-28.
[168] Liu W Y, Yang H W, Wei X H,etal.Effects of exogenous nitric oxide on seed germination, physiological characteristics and active oxygen metabolism ofMedicagotruncatula.Acta Prataculturae Sinica, 2015, 24(2):85-95.
[169] Chao D Y, Dilkes B, Luo H,etal.Polyploids exhibit higher potassium uptake and salinity tolerance inArabidopsis.Science, 2013, 341:658-659.
[170] Horie T, Costa A, Kim T H,etal.RiceOsHKT2;1 transporter mediates large Na+influx component into K+-starved roots for growth.EMBO Journal, 2007, 26(12):3013-3014.
[171] James R A, Blake C, Byrt C S,etal.Major genes for Na+exclusion,Nax1 andNax2 (wheatHKT1;4 andHKT1;5), decrease Na+accumulation in bread wheat leaves under saline and waterlogged conditions.Journal of Experimental Botany, 2011, 62:2939-2947.
[172] Byrt C S, Platten J D, Spielmeyer W,etal.HKT1; 5-like cation transporter linked to Na+exclusion loci in wheat,Nax2 andKna1.Plant Physiology, 2007, 143:1918-1928.
參考文獻:
[5] 趙可夫, 李法曾, 樊守金, 等.中國的鹽生植物.植物學通報, 1999, 16(3):201-207.
[6] 趙可夫.植物對鹽漬逆境的適應.生物學通報, 2002, 37(6):7-10.
[13] 李昀, 沈禹穎, 閻順國.NaCl脅迫下5種牧草種子萌發的比較研究.草業科學, 1997, 14(2):50-53.
[19] 劉晶, 才華, 劉瑩, 等.兩種紫花苜蓿苗期耐鹽生理特性的初步研究及其耐鹽性比較.草業學報, 2013, 22(2):250-256.
[40] 王鎖民, 朱興運, 舒孝喜.堿茅離子吸收與分配特性研究.草業學報, 1994, 3(1):39-43.
[51] 管博, 于君寶, 陸兆華, 等.黃河三角洲濱海濕地水鹽脅迫對鹽地堿蓬幼苗生長和抗氧化酶活性的影響.環境科學, 2011, 32(8):2422-2429.
[52] 魯艷, 雷加強, 曾凡江, 等.NaCl處理對梭梭生長及生理生態特征的影響.草業學報, 2014, 23(3):152-159.
[53] 薛秀棟, 董曉穎, 段艷欣, 等.不同鹽濃度下3種結縷草的耐鹽性比較研究.草業學報, 2013, 22(6):315-320.
[59] 張宏飛, 王鎖民.高等植物Na+吸收、轉運及細胞內Na+穩態平衡研究進展.植物學通報, 2007, 24(5):561-571.
[76] 張金林.鹽生植物海濱堿蓬Na+吸收和積累的研究[D].蘭州: 蘭州大學, 2008.
[146] 張寧, 王蒂, 司懷軍.菠菜甜菜堿醛脫氫酶基因的分離和誘導表達.農業生物技術學報, 2004, 12(5):612-613.
[160] 葛瑛, 高鵬, 夏激宗, 等.氯化鈣在提高玉米抗鹽性方面的作用.東北農業大學學報, 2004, 35(3):281-284.
[161] 張利霞, 常青山, 侯小改, 等.不同鈉鹽脅迫對夏枯草種子萌發特性的影響.草業學報, 2015, 24(3):177-186.
[162] 錢前, 瞿禮嘉, 袁明, 等.2012年中國植物科學若干領域重要研究進展.植物學報, 2013, 48:231-287.
[165] 安建平, 陳靠山.滲透脅迫下小麥的膜損傷與ABA增高的關系.蘭州大學學報, 1994, (3):127-131.
[167] 袁芳, 楊劍超, 陳敏, 等.NaCl 脅迫下外源NO供體硝普鈉(SNP)對鹽地堿蓬種子萌發的影響.植物生理學通訊, 2010, 46(1):24-28.
[168] 劉文瑜, 楊宏偉, 魏小紅, 等.外源NO調控鹽脅迫下蒺藜苜蓿種子萌發生理特性及抗氧化酶的研究.草業學報, 2015, 24(2):85-95.
Research advances in higher plant adaptation to salt stress
ZHANG Jin-Lin1*, LI Hui-Ru1, GUO Shu-Yuan1, WANG Suo-Min1, SHI Hua-Zhong2, HAN Qing-Qing1, BAO Ai-Ke1, MA Qing1
1.StateKeyLaboratoryofGrasslandAgro-ecosystems,CollegeofPastoralAgricultureScienceandTechnology,LanzhouUniversity,Lanzhou730020,China; 2.DepartmentofChemistryandBiochemistry,TexasTechUniversity,LubbockTX79409,USA
Soil salinity is a serious worldwide problem causing reduction in crop growth and agricultural output potential.Consequently, finding new ways to minimize the adverse effects of soil salinization on agriculture is globally important.Understanding the adaptation mechanisms of higher plants to salt stress is critical for enhancing salt tolerance and yields of crop plants as well as protecting ecological environments.In this paper, we reviewed the key progresses in salt stress adaptation of higher plants, including the effects of salt stress in plants; physiological mechanism of plant salt tolerance (osmotic adjustment, nutrient balance and the antioxidant system); the diversity of genes relevant to salt tolerance (ion transporting protein genes, osmotic regulation-related genes, signal transduction-related genes and cellular antioxidant-related genes and so on); and the approaches for crop improvement in salt tolerance.Prospects for developing crop plants tolerant to salinity are also discussed.
higher plants; salt stress; salt tolerance; salt tolerant genes
10.11686/cyxb2015233
http://cyxb.lzu.edu.cn
2015-05-07;改回日期:2015-07-14
國家自然科學基金項目(31222053,31170431和31172256),教育部“長江學者和創新團隊發展計劃”(IRT13019)和中央高校基本科研業務費項目(lzujbky-2014-m01和lzujbky-2015-194)資助。
張金林(1975-),男,甘肅涇川人,教授,博士生導師,博士。
*通信作者Corresponding author.E-mail:jlzhang@lzu.edu.cn
張金林, 李惠茹, 郭姝媛, 王鎖民, 施華中, 韓慶慶, 包愛科, 馬清.高等植物適應鹽逆境研究進展.草業學報, 2015, 24(12):220-236.
ZHANG Jin-Lin, LI Hui-Ru, GUO Shu-Yuan, WANG Suo-Min, SHI Hua-Zhong, HAN Qing-Qing, BAO Ai-Ke, MA Qing.Research advances in higher plant adaptation to salt stress.Acta Prataculturae Sinica, 2015, 24(12):220-236.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
