桑青枯雷爾氏菌gsp G和gsp K基因的獲取與生物信息學(xué)分析
2015-05-06 15:02:25禚蘇孔衛(wèi)青
山東農(nóng)業(yè)科學(xué) 2015年3期
禚蘇++孔衛(wèi)青

摘要: gsp G和gsp K參與青枯雷爾氏菌Ⅱ型分泌系統(tǒng)周質(zhì)復(fù)合體的形成。本研究以桑青枯雷爾氏菌MR111為材料,利用PCR擴(kuò)增和DNA測序技術(shù)成功獲得其gsp G和gsp K基因438 bp和843 bp的序列,在線BLAST分析兩個(gè)基因編碼蛋白與其它青枯雷爾氏菌同源區(qū)域的相似性在90%以上。在線CDD分析兩個(gè)蛋白除分別含有典型的T2SG和T2SK以及N端主要由亮氨酸和丙氨酸組成的分泌型信號(hào)肽外,gsp G還含Ⅳ型分泌系統(tǒng)的“Ⅳ_pilin_GFxxxE”結(jié)構(gòu),而在gsp K中則出現(xiàn)了重復(fù)的“GIQSTE”序列,該序列在雷爾氏菌屬中高度保守。
關(guān)鍵詞:桑樹;青枯雷爾氏菌;Ⅱ型分泌系統(tǒng);gsp基因
中圖分類號(hào):S432.4+2文獻(xiàn)標(biāo)識(shí)號(hào):A文章編號(hào):1001-4942(2015)03-0009-04
Acquisition and Bioinformatics Analysis of gsp G and gsp K Genes
of Ralstonia solanacearum from Mulberry
Zhuo Su, Kong Weiqing*
(College of Agriculture and Life Sciences, Ankang University/Key Sericultural Laboratory of Shaanxi, Ankang 725099, China)
Abstractgsp G and gsp K genes are involved in the formation of periplasmic complex of Ralstonia solanacearum type Ⅱ secretion system. The sequences of gsp G and gsp K genes of Ralstonia solanacearum strain MR111 with 438 bp and 843 bp in length respectively were obtained from mulberry by PCR amplication and DNA sequencing technology. By online BLAST analysis, the similarity of the 2 genes with the homologous genes from other R. solanacearum species were both over 90%. There were typical functional structure of T2SG and T2SK, and secretory signal peptide composed mainly by leucine and alanine in the N-end in the 2 genes, respectively. A type Ⅳ secretion system “Ⅳ_pilin_GFxxxE” structure was found in gsp G, and a repeat sequence “GIQSTE” was found in gsp K. This repeat sequence was highly conserved in R. solanacearum.
Key wordsMulberry; Ralstonia solanacearum; Type Ⅱ secretion system; gsp gene
植物病原細(xì)菌通過泌出毒性因子和各種酶類與寄主植物蛋白進(jìn)行互作完成其侵染過程,并使寄主植物產(chǎn)生病斑、枯萎等病癥[1]。目前病原細(xì)菌主要存在4種類型的分泌系統(tǒng),Ⅰ型分泌系統(tǒng)主要分泌蛋白酶、毒素、脂肪酶等[2];Ⅲ型分泌系統(tǒng)通過干擾抑制植物防御反應(yīng)、調(diào)節(jié)寄主細(xì)胞的生理過程,幫助其對寄主的侵染,這兩種分泌系統(tǒng)均是直接將底物蛋白從細(xì)胞質(zhì)分泌到胞外[3];Ⅳ型分泌系統(tǒng)被認(rèn)為是接觸依賴性系統(tǒng),類似一個(gè)大的菌毛結(jié)構(gòu),橫跨內(nèi)外膜,使蛋白和大分子輸出、靶定寄主細(xì)胞[4];Ⅱ型分泌系統(tǒng)則主要分泌淀粉酶、蛋白酶、纖維素酶以及細(xì)胞毒素等,降解植物細(xì)胞壁并產(chǎn)生大量的胞外多糖,使植株導(dǎo)管阻塞,植物體內(nèi)水分的運(yùn)輸受到阻礙,或引起過高的靜水力學(xué)壓力,導(dǎo)致導(dǎo)管破裂,最終引起植株萎蔫[5]。
青枯雷爾氏菌(Ralstonia solanacearum)能單獨(dú)存活于土壤并在其中繁殖,可以從次生根的根冠部位直接侵入植物,致病性、致病機(jī)理復(fù)雜[6]。由該菌引起的植物青枯病可危害大姜等多種植物,是一種危害嚴(yán)重的土傳性細(xì)菌維管束病害[7]。青枯雷爾氏菌Ⅱ型分泌系統(tǒng)的中心由分泌途徑轉(zhuǎn)膜蛋白(general secretory pathway transmembrane protein,gsp)基因簇共12個(gè)蛋白gsp C~N組成,呈“致病島”的形式排列在基因組上[8]。其中很多蛋白和Ⅳ型分泌系統(tǒng)的菌毛合成中的蛋白有明顯的序列相似性,尤其gsp G、H、I、J和K 5個(gè)蛋白與菌毛蛋白有高相似性,被稱為“類菌毛亞基”,這5個(gè)蛋白還可能在周質(zhì)中形成復(fù)合體,與細(xì)菌的外膜孔直接接觸,將蛋白泌出胞外[9]。本試驗(yàn)以桑青枯雷爾氏菌為研究對象,對其gsp G和gsp K基因及其特征進(jìn)行分析研究,為細(xì)菌Ⅱ型分泌系統(tǒng)的結(jié)構(gòu)和泌出機(jī)制的研究奠定基礎(chǔ)。
1材料與方法
1.1試驗(yàn)材料
供試菌株:桑青枯雷爾氏菌MR111,由本實(shí)驗(yàn)室保存。
1.2基因組DNA的提取和gsp基因的擴(kuò)增
桑青枯雷爾氏菌的培養(yǎng)在26℃下進(jìn)行,使用TTC固體培養(yǎng)基對保存菌株進(jìn)行活化,SPA液體培養(yǎng)基對挑取的單菌落過夜振蕩培養(yǎng)[10]。后離心收集菌體,按細(xì)菌基因組DNA提取試劑盒(天根生化科技北京有限公司產(chǎn)品)說明提取培養(yǎng)的桑青枯雷爾氏菌的基因組DNA,并以此為模板,使用設(shè)計(jì)的基因擴(kuò)增引物進(jìn)行PCR擴(kuò)增。引物序列分別為:gsp G的引物序列為GF:5′-ATGATGCAAGGCCAAC-3′;GR:5′- CATTGTCCCAGTTGCC-3′;gsp K的引物序列分別為KF:5′-CGGGGCTCTGACATGA-3′;KR:5′-CCGAGGGGTATTGTGCC-3′。PCR反應(yīng)程序?yàn)椋?4℃預(yù)變性5 min;94℃變性30 s,55℃退火45 s,72℃延伸1 min,30個(gè)循環(huán);72℃終延伸10 min。PCR擴(kuò)增產(chǎn)物進(jìn)行1.2%瓊脂糖凝膠電泳和回收純化,送上海生工武漢測序部進(jìn)行測序。endprint
1.3gsp G和gsp K基因序列的生物信息學(xué)分析
NCBI在線BLAST對MR111的gsp G和gsp K基因編碼蛋白序列進(jìn)行比對;在線CDD預(yù)測基因編碼蛋白的結(jié)構(gòu)域;Target P1.1預(yù)測蛋白信號(hào)肽類型[11];NPS (http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_compo.html)分析信號(hào)肽氨基酸組成。下載相關(guān)序列,Clustal X1.83軟件進(jìn)行蛋白多序列比對[12],BoxShade3.21(http://www.ch.embnet.org/software/BOX_form.html)對比對的序列文件進(jìn)行描影和印刷。
2結(jié)果與分析
2.1gsp G和gsp K基因的獲取
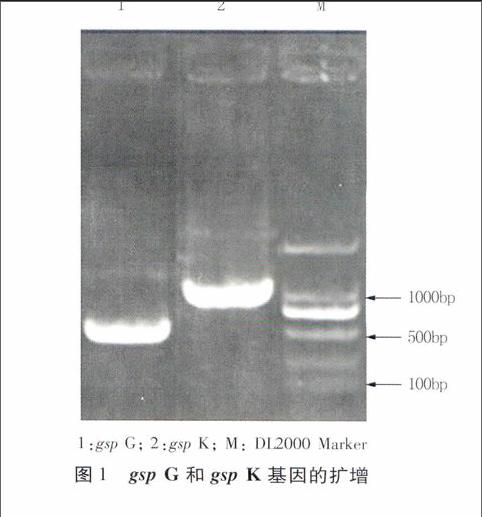
以提取的桑青枯雷爾氏菌MR111的基因組DNA為模板,2個(gè)引物對GF、GR和KF、KR分別進(jìn)行PCR擴(kuò)增gsp G和gsp K基因,1.2%瓊脂糖凝膠電泳檢測PCR擴(kuò)增產(chǎn)物,結(jié)果分別在約500 bp和1 000 bp下方獲得單一擴(kuò)增條帶(圖1)。回收兩條序列進(jìn)行測序,結(jié)果獲得gsp G基因自起始密碼子下游34 bp的長438 bp的序列,和gsp K基因位于25~867 bp間843 bp的序列(GenBank登錄號(hào)分別為KM115546、KM115547)。
2.2gsp G和gsp K基因編碼蛋白的多序列比對與結(jié)構(gòu)分析
NCBI在線BLASTP比對分析,gsp G基因編碼蛋白的保守性較高,與其他來源青枯雷爾氏菌的同源區(qū)域的相似性在98%以上,與皮氏羅爾斯頓氏菌(Ralstonia pickettii)和雷爾氏菌(Ralstonia sp.)的相似性在95%以上,與伯克氏菌(Cupriavidus sp.)和伯克霍爾氏菌(Burkholderia sp.)的相似性也在90%左右。相比之下,gsp K蛋白的保守性稍差,其與其他來源青枯雷爾氏菌同源區(qū)域的相似性在90%以上,與皮氏羅爾斯頓氏菌和雷爾氏菌的相似性在80%左右,與伯克氏菌的相似性在70%以下,與伯克霍爾氏菌的相似性小于60%。獲得的gsp G蛋白N末端含有N末端切割/甲基化域“Ⅳ_pilin_GFxxxE”結(jié)構(gòu),這與其它細(xì)菌中的研究一致[13],而對gsp K蛋白的比對發(fā)現(xiàn),173~178氨基酸處含有一“GIQSTE”結(jié)構(gòu),該結(jié)構(gòu)在雷爾氏菌屬中普遍存在,但在本研究的MR111菌種中發(fā)現(xiàn)有2個(gè)重復(fù)(圖2),是否對其功能有影響還需進(jìn)一步的研究證明。
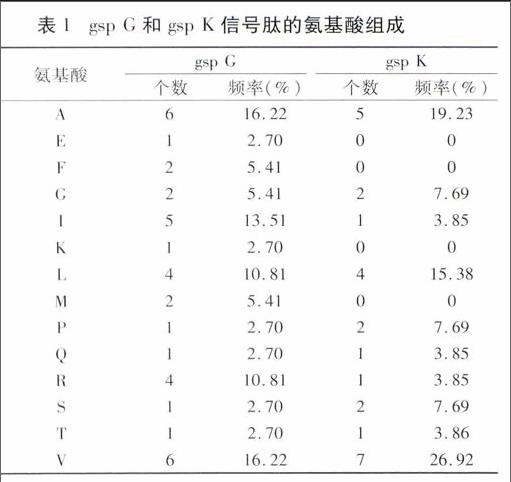
NCBI在線CDD預(yù)測基因編碼蛋白結(jié)構(gòu)域,結(jié)果見圖3。gsp G編碼蛋白含典型的、具有高度保守性的T2SG(pfam08334)結(jié)構(gòu),屬于T2SG超家族成員;gsp K編碼蛋白含T2SK(pfam03934)結(jié)構(gòu)。同時(shí)經(jīng)Target P1.1預(yù)測,gsp G和gsp K蛋白均含分泌型信號(hào)肽,切割位點(diǎn)分別位于37和26位氨基酸。NPS分析信號(hào)肽氨基酸組成(表1),結(jié)果顯示2個(gè)信號(hào)肽均以疏水氨基酸纈氨酸(V)和丙氨酸(A)最多。
3討論
Ⅰ型分泌系統(tǒng)廣泛存在于人類、動(dòng)物和植物的革蘭氏陰性病原細(xì)菌中,是革蘭氏陰性細(xì)菌的常規(guī)代謝途徑,參與寄主植物防御反應(yīng)的激活[5]。Ⅱ型分泌系統(tǒng)中基因的突變會(huì)造成許多蛋白如降解植物細(xì)胞壁不同成分的蛋白分泌的缺乏,也可使細(xì)菌的毒性減弱或喪失[8,14]。青枯雷爾氏菌是世界范圍內(nèi)重要的植物病原細(xì)菌,該菌通過Ⅱ型分泌系統(tǒng)分泌多種胞外毒性蛋白,降解植物細(xì)胞壁,并產(chǎn)生大量胞外多糖造成植物維管束阻塞,使寄主植物萎蔫,是研究植物病原細(xì)菌致病機(jī)制的一種模式菌[15]。本研究獲得了桑青枯雷爾氏菌MR111的T2S的gsp G和gsp K基因,其編碼蛋白均含有Ⅱ型分泌系統(tǒng)蛋白的功能結(jié)構(gòu)域。除此之外,gsp G蛋白還含有Ⅳ型分泌系統(tǒng)N末端切割/甲基化域“Ⅳ_pilin_GFxxxE”結(jié)構(gòu),該結(jié)構(gòu)又被稱為菌毛蛋白的甲基化位點(diǎn),首先在Ⅳ型分泌系統(tǒng)中發(fā)現(xiàn),主要存在于細(xì)菌的絲狀表面蛋白或菌毛的N末端,在疏水性苯丙氨酸或蛋氨酸發(fā)生甲基化后,形成成熟的親水前導(dǎo)肽,其中的谷氨酸(E)是該結(jié)構(gòu)必須的[13]。這也驗(yàn)證了Ⅱ型和Ⅳ型分泌系統(tǒng)存在結(jié)構(gòu)上的一致性[16]。本研究發(fā)現(xiàn)gsp K蛋白的“GIQSTE”序列在雷爾氏菌屬中非常保守,而在非雷爾氏菌屬中存在較大差異,且在桑青枯雷爾氏菌MR111中發(fā)現(xiàn)了“GIQSTE”的重復(fù)序列,這在gsp K的同源基因研究中未見報(bào)道,其形成原因和是否具有功能變化有待進(jìn)一步研究。
氨基酸組成是決定蛋白功能的重要指標(biāo)。分析gsp G和gsp K兩個(gè)基因編碼蛋白的信號(hào)肽序列的氨基酸組成,分別有37和26個(gè)氨基酸,非極性氨基酸中以V、A和L最多,gsp G帶正電荷的氨基酸以R最多,gsp K不帶電荷的極性氨基酸以S最多。這與黃俊麗等[18]根據(jù)青枯雷爾氏菌基因組分析的分泌型信號(hào)肽的氨基酸組成中,分泌型信號(hào)肽的長度在15~55個(gè)氨基酸,非極性氨基酸中,殘基W出現(xiàn)頻率最低,A和L出現(xiàn)頻率最高,帶正電荷氨基酸R出現(xiàn)頻率最高,不帶電荷的極性氨基酸中,S出現(xiàn)頻率最高等情況相似。BLAST在線分析和多序列比對顯示,gsp G和gsp K與青枯雷爾氏菌同源區(qū)域的一致性最高,與皮氏羅爾斯頓氏菌和雷爾氏菌屬的相似性次之,與伯克氏菌和伯克霍爾氏菌相似性最低,這與Bouley等[19]的研究認(rèn)為T2S具有細(xì)菌種的專一性相一致。
參考文獻(xiàn):
[1] Bogdanove A J. Protein-protein interactions in pathogen recognition by plants[J]. Plant Mol. Biol., 2002, 50(6): 981-989.
[2]Koster M, Bitter W, Tommassen J. Protein secretion mechanisms in Gram-negative bacteria[J]. Int. J. Med. Microbiol., 2000, 290(4/5): 325-331.endprint
[3]Jin Q, Thilmony R, Zwiesler-Vollick J, et al. Type Ⅲ protein secretion in Pseudomonas syringae[J]. Microbes Infect., 2003, 5(4): 301-310.
[4]Backert S, Meyer T F. Type IV secretion systems and their effectors in bacterial pathogenesis[J]. Curr. Opin. Microbiol., 2006, 9(2):207-217.
[5]Sandkvist M. Type II secretion and pathogenesis[J]. Infect. Immun., 2001, 69(6): 3523-3535.
[6]Hayward A C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum[J]. Annual Review of Phytopathology, 1991, 29(1): 65-87.
[7]嚴(yán)金平, 澤桑梓, 張火云, 等. 姜細(xì)菌性青枯病病原菌及其防治研究進(jìn)展[J]. 河南農(nóng)業(yè)科學(xué), 2004, 33(9): 63-65.
[8]Genin S, Boucher C. Ralstonia solanacearum: secrets of a major pathogen unveiled by analysis of its genome[J]. Mol. Plant Pathol., 2002, 3(3): 111-118.
[9]Douet V, Loiseau L, Barras F, et al. Systematic analysis by the yeast two-hybrid of protein interaction between components of the type II secretory machinery of Erwinia chrysanthemi[J]. Res. Microbiol., 2004, 155(2): 71-75.
[10]呂志強(qiáng), 王漢榮, 周勤, 等. 浙江省桑樹青枯病菌生理小種及生化型的測定[J]. 浙江農(nóng)業(yè)學(xué)報(bào), 2007, 19(4): 306-309.
[11]Emanuelsson O, Nielsen H, Brunak S, et al. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence[J]. J. Mol. Biol., 2000, 300(4): 1005-1016.
[12]Thompson J D, Gibson T J, Plewniak F, et al. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools[J]. Nucleic Acids Res., 1997, 25(24): 4876-4882.
[13]Peabody C R, Chung Y J, Yen M R, et al. Type II protein secretion and its relationship to bacterial type IV pili and archaeal flagella[J]. Microbiology, 2003, 149(11): 3051-3072.
[14]Genin S, Boucher C. Lessons learned from the genome analysis of Ralstonia solanacearum[J]. Annu. Rev. Phytopathol., 2004, 42(1): 107-134.
[15]劉波, 林營志, 朱育菁, 等. 青枯雷爾氏菌多態(tài)性研究[M]. 福州:福建科學(xué)技術(shù)出版社, 2005.
[16]Tosi T, Estrozi L F, Job V, et al. Structural similarity of secretins from type II and type III secretion systems[J]. Structure, 2014, 22(9): 1348-1355.
[17]劉雅婷, 李成云, 李永忠, 等. 應(yīng)用計(jì)算機(jī)手段分析植物病原細(xì)菌Ralstonia solanacearum的蛋白質(zhì)序列[J]. 云南農(nóng)業(yè)大學(xué)學(xué)報(bào), 2007, 22(4): 485-490.
[18]黃俊麗, 吳金鐘, 肖崇剛, 等. 植物病原細(xì)菌Ralstonia solanacearum GMll000中分泌蛋白信號(hào)肽分析[J].遺傳, 2007, 29(11): 1409-1416.
[19]Bouley J, Condemine G, Shevchik V E. The PDZ domain of OutC and the N-terminal region of OutD determine the secretion specificity of the type II out pathway of Erwinia chrysanthemi[J]. J. Mol. Biol., 2001, 308(2): 205-219.endprint
