低鉀脅迫對(duì)煙草幼苗碳氮代謝基因表達(dá)譜的影響
2015-04-18 08:35:48魯黎明魯逸飛李立芹
中國(guó)煙草科學(xué) 2015年4期
魯黎明,陳 勇,魯逸飛,李立芹
(四川農(nóng)業(yè)大學(xué)農(nóng)學(xué)院,成都 611130)
低鉀脅迫對(duì)煙草幼苗碳氮代謝基因表達(dá)譜的影響
魯黎明,陳 勇,魯逸飛,李立芹*
(四川農(nóng)業(yè)大學(xué)農(nóng)學(xué)院,成都 611130)
為探索煙草響應(yīng)低鉀脅迫的分子機(jī)制,對(duì)K326煙草幼苗進(jìn)行了0、6、12和24 h的低鉀脅迫處理,并利用定制的煙草全基因組芯片,分析了煙草幼苗基因表達(dá)譜的變化。結(jié)果表明,與0 h相比,6、12和24 h的3個(gè)時(shí)間點(diǎn)差異表達(dá)在2倍以上(p<0.05)的基因總數(shù)為3790個(gè)。從功能方面看,這些差異表達(dá)基因廣泛參與了煙草的各種生物學(xué)過(guò)程,包括抗氧化活性、應(yīng)激反應(yīng)、運(yùn)輸活性、發(fā)育過(guò)程、催化活性、生物調(diào)節(jié)、代謝過(guò)程等。其中包括了硝酸還原酶基因NIA2等10個(gè)參與煙草氮代謝的基因,以及UGP等44個(gè)參與煙草碳水化合物與糖代謝的基因。本研究結(jié)果說(shuō)明,低鉀脅迫對(duì)煙草的基因表達(dá)產(chǎn)生了廣泛的影響,進(jìn)而可能影響到煙草的碳氮等基礎(chǔ)代謝。
煙草;低鉀脅迫;基因表達(dá)譜;基因芯片;碳氮代謝
鉀是煙草的一個(gè)重要品質(zhì)元素。然而,與國(guó)外優(yōu)質(zhì)煙葉相比,我國(guó)煙葉中的鉀含量較低[1-2]。研究表明,由于植物生長(zhǎng)對(duì)鉀的需求量較大,土壤中含鉀量往往不足以滿足植物生長(zhǎng)發(fā)育的需要[3]。為適應(yīng)這種低鉀環(huán)境,植物產(chǎn)生了一整套調(diào)節(jié)機(jī)制,如功能基因的誘導(dǎo)表達(dá)[4-9]、信號(hào)轉(zhuǎn)導(dǎo)與調(diào)節(jié)途徑的運(yùn)作[10-11]。近年來(lái),基因芯片技術(shù)以其高通量、全方位的特點(diǎn),在植物響應(yīng)非生物脅迫的機(jī)制研究方面得到了廣泛的應(yīng)用[12]。如擬南芥[13-15]及水稻[16-20]分別響應(yīng)低氮、低磷的研究均表明,在脅迫條件下,擬南芥及水稻的基因表達(dá)譜均發(fā)生了較大的變化。其中,Ma等[11]對(duì)水稻響應(yīng)低鉀脅迫的研究證實(shí),編碼碳氮代謝、信號(hào)蛋白、轉(zhuǎn)錄因子及礦質(zhì)元素轉(zhuǎn)運(yùn)蛋白的基因,均參與了這種脅迫響應(yīng)。在煙草的生長(zhǎng)發(fā)育后期,也經(jīng)常遇到土壤中有效鉀含量不足的問(wèn)題。所以,探索煙草響應(yīng)低鉀脅迫的分子機(jī)制,對(duì)于提高煙葉鉀含量具有重要的意義。因此,本研究利用基因芯片技術(shù),分析低鉀脅迫下煙草參與碳氮代謝的基因表達(dá)譜的變化。
1 材料與方法
1.1 試驗(yàn)材料及處理
試驗(yàn)材料為烤煙品種K326(Nicotiana tabacum cv. K326),基因芯片為美國(guó)安捷倫公司煙草全基因組芯片。
煙草種子表面消毒后,播種于MS培養(yǎng)基上,并放于光照培養(yǎng)室進(jìn)行全光照培養(yǎng),培養(yǎng)溫度為28 ℃。無(wú)菌苗培養(yǎng)4周后,進(jìn)行低鉀脅迫處理(0、6、12和24 h),重復(fù)3次。將煙草幼苗移到低鉀培養(yǎng)基[21]上,并在上述4個(gè)時(shí)間點(diǎn)進(jìn)行取樣,迅速于液氮中冷凍備用。
1.2 基因表達(dá)譜芯片分析
煙草樣品總RNA的抽提采用TRIZOL法,按照試劑盒說(shuō)明書的要求進(jìn)行。使用QIAGEN RNeasy? Kit 純化總RNA。然后,使用Nanodrop ND-1000及Agilent BioAnalyzer 2100進(jìn)行總RNA質(zhì)檢。
芯片雜交:cDNA的合成、cRNA的制備、Cy3單色熒光標(biāo)記、標(biāo)記產(chǎn)物的片段化等均按照美國(guó)安捷倫公司的基因芯片雜交標(biāo)準(zhǔn)流程進(jìn)行。芯片雜交過(guò)程與數(shù)據(jù)采集由上海伯豪生物技術(shù)有限公司芯片雜交平臺(tái)完成。
1.3 數(shù)據(jù)處理與分析
基因表達(dá)信號(hào)經(jīng)歸一化后,各處理與對(duì)照信號(hào)值進(jìn)行比對(duì),若foldchange變化的比值≥2為該基因表達(dá)上調(diào),比值≤-2.0則該基因表達(dá)下調(diào),以此篩選出每組對(duì)比中的差異表達(dá)基因。登陸上海伯豪生物公司的SAS分析系統(tǒng)(http://www. shanghaibiotech.com)進(jìn)行數(shù)據(jù)分析,并對(duì)差異表達(dá)基因進(jìn)行GO(Gene Ontology)功能分析。
Cluster聚類分析使用Cluster3.0軟件進(jìn)行。差異基因的表達(dá)趨勢(shì)分析采用STEM(Short Timeseries Expression Miner)軟件進(jìn)行。
1.4 RT-qPCR驗(yàn)證
隨機(jī)挑選出3個(gè)上調(diào)表達(dá)和3個(gè)下調(diào)表達(dá)基因作為驗(yàn)證基因,引物采用primer 3.0軟件進(jìn)行設(shè)計(jì),由生工生物工程有限公司合成。RT采用Invitrogen的反轉(zhuǎn)錄試劑盒,按照說(shuō)明書進(jìn)行。SYBR Green qPCR采用10 μL反應(yīng)體系(2×SYBR Green PCR buffer 5ul,正反向引物各0.2 μL,模板DNA 5 ng,用ddH2O加至10 μL),反應(yīng)條件為95 ℃,5 min,95 ℃,10 s,60 ℃,30 s,40個(gè)循環(huán)。
2 結(jié) 果
2.1 差異表達(dá)基因總體特征
利用上海伯豪生物公司的SAS分析系統(tǒng),對(duì)芯片數(shù)據(jù)進(jìn)行了分析。以P<0.05且變化倍數(shù)在2倍以上為條件,梳理了差異表達(dá)的基因。3個(gè)時(shí)間點(diǎn)共計(jì)富集到3790個(gè)差異表達(dá)基因(DEGs)。與0 h相比,6 h時(shí)間點(diǎn)差異表達(dá)基因達(dá)2277個(gè),其中,上調(diào)表達(dá)1270個(gè),下調(diào)表達(dá)1007個(gè);12 h為1954個(gè),其中,上調(diào)表達(dá)1159個(gè),下調(diào)表達(dá)795個(gè);24 h為1835個(gè),其中,上調(diào)表達(dá)1023個(gè),下調(diào)表達(dá)812個(gè)。6 h時(shí)間點(diǎn)差異表達(dá)基因數(shù)量較多,同時(shí),在3個(gè)時(shí)間點(diǎn),上調(diào)基因的數(shù)量總是比下調(diào)基因多。在上調(diào)表達(dá)的DEGs中,3個(gè)處理時(shí)間點(diǎn)一直都處于上調(diào)的基因有300個(gè);在下調(diào)表達(dá)的DEGs中,3個(gè)處理時(shí)間點(diǎn)一直都處于下調(diào)的基因有257個(gè),共計(jì)557個(gè)。該類基因在低鉀脅迫后一直處于活躍狀態(tài),可能與低鉀脅迫有著密切的聯(lián)系。
差異表達(dá)基因可以分為8種表達(dá)趨勢(shì)(圖1,A-H),趨勢(shì)中各基因的表達(dá)一致性較強(qiáng),變化動(dòng)態(tài)較一致。這些表達(dá)趨勢(shì)相似的基因,可能在某些生理生化過(guò)程中處于協(xié)同關(guān)系,或者具有類似的功能。
2.2 RT-qPCR驗(yàn)證
為檢驗(yàn)基因芯片檢測(cè)結(jié)果的準(zhǔn)確性,在差異表達(dá)基因中,隨機(jī)挑選了3個(gè)上調(diào)和3個(gè)下調(diào)基因作為驗(yàn)證基因,進(jìn)行RT-qPCR的驗(yàn)證。結(jié)果表明基因芯片技術(shù)對(duì)基因表達(dá)檢測(cè)的準(zhǔn)確性較高。

圖1 煙草響應(yīng)低鉀脅迫的基因表達(dá)模式分析Fig. 1 Expression profiles of tobacco seedlings under potassium deficiency
2.3 差異表達(dá)基因的功能分析
在P(0.05)顯著水平下,這些差異表達(dá)基因可以歸納為18種功能,如抗氧化活性、結(jié)構(gòu)分子活性、電子載體活性、應(yīng)激反應(yīng)等(圖2)。說(shuō)明低鉀脅迫引發(fā)了煙草幼苗多種生理生化反應(yīng)。

表1 RT-qPCR所用引物Table1 Primers for RT-qPCR
2.4 氮代謝過(guò)程相關(guān)基因
氮代謝是植物體內(nèi)的基礎(chǔ)代謝過(guò)程,直接影響著植物的生長(zhǎng)發(fā)育,也是植物產(chǎn)量形成的主要影響因素[22-24]。在本研究中,低鉀引起了氮代謝過(guò)程中的相關(guān)基因的變化(表2)。如硝酸還原酶NIA2、谷氨酸合成酶GLT1、谷氨酰胺合成酶AtGSR1等。這些基因均集中于植物氮代謝的起點(diǎn),即無(wú)機(jī)氮同化為有機(jī)氮的過(guò)程。
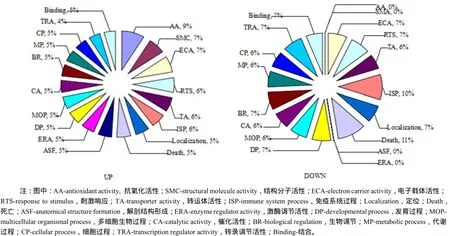
圖2 差異表達(dá)基因的功能分析Fig. 2 GO analysis of DEGs

表2 氮代謝途徑中的重要基因Table 2 Nitrogen metabolic genes
2.5 碳水化合物代謝相關(guān)基因
碳水化合物及糖類代謝是植物的最基本的初生代謝,直接影響著植物的基本生命活動(dòng)[25-26]。本研究發(fā)現(xiàn),在低鉀脅迫下,煙草幼苗參與碳水化合物代謝的基因中,有44個(gè)基因的表達(dá)發(fā)生了顯著變化,其中上調(diào)28個(gè),下調(diào)16個(gè)(表3)。這些差異表達(dá)基因主要參與了光合作用碳同化、呼吸作用之糖酵解與PPP途徑、蔗糖合成以及淀粉代謝等生化過(guò)程。本結(jié)果說(shuō)明,低鉀脅迫可能導(dǎo)致了煙草幼苗光合作用與呼吸作用發(fā)生改變。
3 討 論
3.1 低鉀對(duì)氮代謝的影響
本試驗(yàn)結(jié)果表明,低鉀脅迫導(dǎo)致了煙草編碼硝酸還原酶、谷氨酸合成酶與谷氨酰胺合成酶等基因的表達(dá)均發(fā)生了明顯變化。此結(jié)果與擬南芥及水稻的研究結(jié)果相一致[10-11]。在本研究中,硝酸還原酶基因的表達(dá)在3個(gè)時(shí)間點(diǎn)均處于下調(diào),而谷氨酰胺合成酶基因則均上調(diào)表達(dá)。由于這些基因在根部參與無(wú)機(jī)氮同化過(guò)程,因此,可以推測(cè),在外界環(huán)境缺鉀時(shí),煙草氮同化會(huì)受到抑制,從而可能會(huì)抑制煙草的生長(zhǎng)。
3.2 低鉀對(duì)碳代謝的影響
研究表明,低鉀環(huán)境中,擬南芥根中碳氮代謝酶基因的表達(dá)發(fā)生了明顯的改變,丙酮酸、蘋果酸及硝酸鹽等的含量也明顯下降[27]。供鉀恢復(fù)正常時(shí),其含量又回歸正常。MA等[11]在水稻的研究也得到了類似的結(jié)果,有兩個(gè)PEPCK基因表達(dá)顯著下調(diào)。本研究也發(fā)現(xiàn),低鉀條件下,煙草有44個(gè)差異表達(dá)基因參與了糖及碳水化合物的代謝。它們參與了碳固定、EMP、PPP、蔗糖合成以及淀粉代謝等途徑。其中,有5個(gè)丙酮酸激酶基因的表達(dá)上調(diào)。由此可見(jiàn),低鉀脅迫時(shí),煙草通過(guò)改變基礎(chǔ)碳代謝基因的表達(dá),調(diào)整基礎(chǔ)代謝,以適應(yīng)外界的脅迫環(huán)境,保證植株的正常生長(zhǎng)。
4 結(jié) 論
在低鉀脅迫下,煙草幼苗眾多基因的表達(dá)發(fā)生了顯著的改變。其中,包括了硝酸還原酶基因NIA2等參與無(wú)機(jī)氮同化的基因,以及UGP等參與碳水化合物代謝的基因。說(shuō)明煙草在低鉀脅迫時(shí),可能通過(guò)調(diào)節(jié)基因的表達(dá),影響到碳氮等基礎(chǔ)代謝。本研究的結(jié)果為煙草響應(yīng)低鉀脅迫的分子機(jī)制研究提供了參考。
[1] 郭清源,丁松爽,劉國(guó)順,等. 鉀用量與灌溉量對(duì)不同土層鉀素及煙葉鉀含量的積累效應(yīng)[J]. 中國(guó)煙草科學(xué),2015,36(1):61-67.
[2] 楊鐵釗,魯黎明,夏巍,等. 烤煙富鉀基因型鉀吸收積累與內(nèi)向鉀電流特性[J].中國(guó)農(nóng)業(yè)科學(xué),2008,41(8):2392-2399.
[3] Ashley M, Grant M, GRABOV A. Plant responses to potassium deficiencies: a role for potassium transport proteins [J]. Journal Experimental Botany, 2006, 57: 425-436.
[4] Gierth M, M?SER P, Schroeder J I.The potassium transporter AtHAK5 functions in K(+) deprivationinduced high-affinity K(+) uptake and AKT1 K(+)channel contribution to K(+) uptake kinetics inArabidopsis roots[J]. Plant Physiology, 2005, 137(3): 1105-1114.
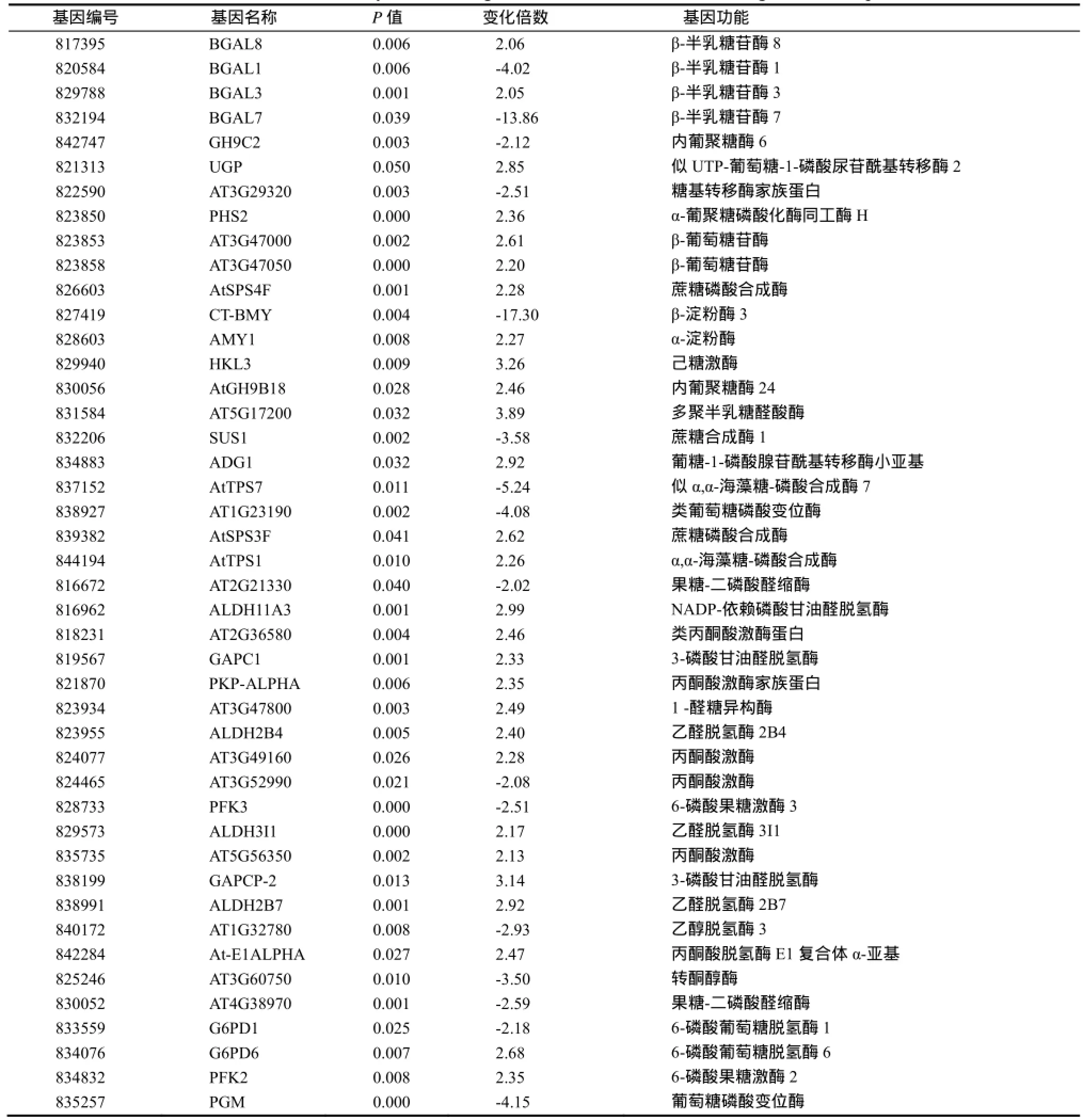
表3 參與碳水化合物及糖代謝相關(guān)差異表達(dá)基因Table 3 DEGs involved in carbohydrate and sugar metabolism of tobacco seedlings under low potassium stress
[5] SANTA-MARíA G E, RUBIO F, DUBCOVSKY J, et al. The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter[J]. Plant Cell, 1997, 9(12): 2281-2290.
[6] FULGENZI F R, PERALTA M L, MANGANO S, et al. The Ionic Environment Controls the Contribution of the Barley HvHAK1 Transporter to Potassium Acquisition[J]. Plant Physiology, 2008, 147(1): 252-262.[7] BA?UELOS M A, GARCIADEBLAS B, CUBERO B,et al. A.Inventory and functional characterization of the HAK potassium transporters of rice[J]. Plant Physiology, 2002, 130(2): 784-795.
[8] BUSCHMANN P H, VAIDYANATHAN R,GASSMANN W, et al. Enhancement of Na(+) uptake currents, time-dependent inward-rectifying K(+)channel currents, and K(+) channel transcripts by K(+)starvation in wheat root cells[J]. Plant Physiolgy, 2000,122(4): 1387-1397.
[9] CELLIER F, CONéJéRO G, RICAUD L, et al. Characterization of AtCHX17, a member of the cation/H+ exchangers, CHX family, from Arabidopsis thaliana suggests a role in K+ homeostasis[J]. The PlantJournal, 2004, 39(6): 834-846.
[10] Armengaud P, Breitling R, Amtmann A. The Potassium-Dependent Transcriptome of Arabidopsis Reveals a Prominent Role of Jasmonic Acid in Nutrient Signaling[J]. Plant Physiology, 2004, 136(1): 2556-2576.
[11] MA T L, WU W H, WANG Y. Transcriptome analysis of rice root responses to potassium deficiency[J]. BMC Plant Biology, 2012, 12: 161-173.
[12] 許志茹,李玉花. 基因芯片技術(shù)在植物研究中的應(yīng)用[J]. 生物技術(shù),2004,14(6):70-72.
[13] KRAPP A, BERTHOME R, ORSEL M, et al. Arabidopsis Roots and Shoots Show Distinct Temporal Adaptation Patterns toward Nitrogen Starvation[J]. Plant Physiology, 2011, 157(3): 1255-1282.
[14] MISSON J, RAGHOTHAMA K G, JAIN A, et al. A genome-wide transcriptional analysis using Arabidopsis thaliana Affymetrix gene chips determined plant responses to phosphate deprivation[J]. Proceeding National Academy Science USA, 2005, 102(33): 11934-11939.
[15] WOO J, MACPHERSON C R, LIU J, et al. The response and recovery of the Arabidopsis thaliana transcriptome to phosphate starvation[J]. BMC Plant Biology, 2012,12: 62-83.
[16] WANG D, PAN Y, ZHAO X, et al. Genome-wide temporal-spatial gene expression profiling of drought responsiveness in rice[J]. BMC Genomics, 2011, 12: 149-163.
[17] LIAN X, WANG S, ZHANG J, et al. Expression profiles of 10,422 genes at early stage of low nitrogen stress in rice assayed using a cDNA microarray[J]. Plant Molecular Biology, 2006, 60(5): 617-631.
[18] WASAKI J, YONETANI R, KURODA S, et al. Transcriptomic analysis of metabolic changes by phosphorus stress in rice plant roots[J]. Plant Cell & Environment, 2003, 26(9): 1515-1523.
[19] WASAKI J, SHINANO T, ONISHI K, et al. Transcriptomic analysis indicates putative metabolic changes caused by manipulation of phosphorus availability in rice leaves[J]. Journal Experimental Botany, 2006, 57(9): 2049-2059.
[20] LI L, LIU C, LIAN X. Gene expression profiles in rice roots under low phosphorus stress[J]. Plant Molecular Biology, 2010, 72(4-5): 423-432.
[21] XU J, LI H D, CHEN L Q, et al. A protein kinase,interacting with two calcineurin B-like proteins,regulates K+transporter AKT1 in Arabidopsis[J]. Cell,2006, 125: 1347-360
[22] 楊志曉,史躍偉,林世峰,等. 烤煙碳氮代謝關(guān)鍵酶活性動(dòng)態(tài)及其與類胡蘿卜素關(guān)系研究[J]. 中國(guó)煙草科學(xué),2014,35(2):59-63.
[23] 武云杰,楊鐵釗,張小全. 谷氨酰胺合成酶抑制劑對(duì)衰老期煙葉氮代謝的影響[J]. 中國(guó)煙草科學(xué),2014,35(1):37-42.
[24] 劉國(guó)順,何永秋,楊永鋒,等. 不同鉀肥配施對(duì)烤煙質(zhì)體色素和碳氮代謝及品質(zhì)的影響[J]. 中國(guó)煙草科學(xué),2013,34(6):49-55.
[25] 史宏志,韓錦峰,趙鵬,等. 不同氮量與氮源下烤煙淀粉酶和轉(zhuǎn)化酶活性動(dòng)態(tài)變化[J]. 中國(guó)煙草科學(xué),1999,20(3):5-8.
[26] 岳紅賓. 不同氮素水平對(duì)煙草碳氮代謝關(guān)鍵酶活性的影響[J]. 中國(guó)煙草科學(xué),2007,28(1):18-20.
[27] ARMENGAUD P, SULPICE R, MILLER A J, et al. Multilevel analysis of primary metabolism provides new insights into the role of potassium nutrition for glycolysis and nitrogen assimilation in Arabidopsis roots[J]. Plant Physiol, 2009, 150: 72-85.
The Impact of Low Potassium Stress on Tobacco Gene Expression Profiles of Carbon and Nitrogen Metabolism
LU Liming, CHEN Yong, LU Yifei, LI Liqin*
(Agronomy College of Sichuan Agriculture University, Chengdu 611130, China)
In order to explore the molecular mechanism of tobacco potassium nutrition, tobacco seedlings of K326 were treated with low potassium stress for 0, 6, 12 and 24 h. Gene expression profiles of tobacco seedlings at each time point were analyzed. The results showed that a total of 3790 genes were detected with a change of two folds or more in expression level (p<0.05). GO analysis showed that these differentially expressed genes can be divided into functional classifications including antioxidant activity, stress response,transport activity, development process, catalytic activities, biological regulation, metabolism, etc. Among them, 10 genes, including the nitrate reductase gene NIA2, were involved in nitrogen metabolism, and 44 genes, such as UGP, were involved in the metabolism of carbohydrate and sugar. The results of this study indicate that low potassium stress has a broad impact on gene expression of tobacco,and it may affect the metabolism of carbon and nitrogen in tobacco.
tobacco; low potassium stress; gene expression profiling; gene chips; carbon and nitrogen metabolism
S572.01
1007-5119(2015)04-0012-06
10.13496/j.issn.1007-5119.2015.04.002
國(guó)家自然科學(xué)基金“E3參與植物響應(yīng)低磷脅迫的分子機(jī)制”(31070244)
魯黎明,男,博士,副教授,研究方向?yàn)闊煵葩涍z傳育種。E-mail:luliming@sicau.edu.cn。*通信作者,E-mail:lilq88@126.com
2015-05-05
2015-07-14
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫(yī)結(jié)合雜志(2017年2期)2017-01-12 18:23:59
當(dāng)代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
