三疣梭子蟹血清凝集素的生物學(xué)特性
2015-04-17 23:57:24郝貴杰王春琳林鋒等
江蘇農(nóng)業(yè)科學(xué) 2015年1期
郝貴杰 王春琳 林鋒 等
摘要:對(duì)三疣梭子蟹(Portunus trituberculatus)血清凝集素的凝集特性進(jìn)行了研究。結(jié)果表明: 三疣梭子蟹血清凝集素對(duì)鯽魚(yú)、中華鱉、草魚(yú)、雞和人類A、B、O型血細(xì)胞沒(méi)有凝集作用,但是對(duì)小鼠、兔的血細(xì)胞表現(xiàn)出較強(qiáng)的凝集活性,其中對(duì)小鼠血細(xì)胞的血凝活性達(dá)到210,對(duì)兔血細(xì)胞的血凝活性達(dá)到28;鹽度對(duì)其凝集活性有較大影響,NaCl濃度在0.6 mol/L以上時(shí)基本失活;凝集素活性最適pH 值為6.0~7.4;血清凝集素對(duì)Ca2+、Mg2+離子有明顯的依賴性,EDTA可以明顯抑制其凝集活性;糖抑制結(jié)果表明,血清凝集素活性能被N-乙酰葡萄糖胺及N-乙酰甘露糖胺特異性抑制;對(duì)血清凝集素進(jìn)行硫酸銨分級(jí)沉淀分離后,發(fā)現(xiàn)凝集素活性主要分布在25%飽和度硫酸銨沉淀區(qū);聚丙烯凝膠電泳(SDS-PAGE)結(jié)果表明,主要蛋白條帶在72~95 ku。
關(guān)鍵詞:三疣梭子蟹;血清凝集素;凝集活性;聚丙烯凝膠電泳
中圖分類號(hào): S966.16文獻(xiàn)標(biāo)志碼: A文章編號(hào):1002-1302(2015)01-0220-04
收稿日期:2014-08-14
基金項(xiàng)目:國(guó)家自然科學(xué)基金(編號(hào):41106123);國(guó)家“863”計(jì)劃(編號(hào):2012AA10A409);浙江省自然科學(xué)基金(編號(hào):LY12C19009);浙江省科技廳重大科技專項(xiàng)(編號(hào):2012C12907-3) 。
作者簡(jiǎn)介:郝貴杰(1979—),女,安徽六安人,碩士,副研究員,主要從事水生動(dòng)物病害及免疫學(xué)研究。 E-mail:melissa511@sina.com。
通信作者:沈錦玉,碩士,研究員,主要從事水生動(dòng)物病害及免疫學(xué)研究。E-mail: shenjinyu@126.com。三疣梭子蟹(Portunus trituberculatus)是一種重要的海洋經(jīng)濟(jì)動(dòng)物,隸屬甲殼綱(Crustacea)、十足目(Decapoda)、梭子蟹科(Portunidae)、梭子蟹屬(Portunus),廣泛分布于中國(guó)、日本等海域[1],因其具有生長(zhǎng)快、肉質(zhì)好的特點(diǎn),深受國(guó)內(nèi)外消費(fèi)者的喜愛(ài)。近年來(lái)隨著育苗和養(yǎng)殖技術(shù)的日臻成熟,梭子蟹養(yǎng)殖面積和養(yǎng)殖產(chǎn)量逐年增加,目前已成為沿海各省海水養(yǎng)殖的主導(dǎo)產(chǎn)品。然而由于梭子蟹養(yǎng)殖規(guī)模的逐年增加,養(yǎng)殖環(huán)境的變化等已對(duì)其免疫防御系統(tǒng)造成了影響,導(dǎo)致梭子蟹自身抗病力下降,對(duì)病害的易感性增加。例如近年來(lái)由弧菌引起的“牙膏病”“乳化病”給梭子蟹帶來(lái)較大危害,也造成了該產(chǎn)業(yè)重大經(jīng)濟(jì)損失,嚴(yán)重制約了梭子蟹養(yǎng)殖業(yè)的持續(xù)健康發(fā)展[2-3]。目前水產(chǎn)養(yǎng)殖戶多用抗生素防治病害,但從使用抗生素等藥物的安全性、抗藥性以及對(duì)水環(huán)境的不良影響等方面考慮,研究梭子蟹的免疫機(jī)制,有效提高蟹本身的抗病能力,顯得越來(lái)越重要。
三疣梭子蟹屬于甲殼類動(dòng)物,無(wú)特異性免疫應(yīng)答,非特異性免疫包括細(xì)胞免疫和體液免疫,凝集素是甲殼類動(dòng)物體液免疫中的一種重要免疫因子[4],其作用就相當(dāng)于脊椎動(dòng)物中的抗體[5-6],具有重要的免疫作用,例如可對(duì)外來(lái)病原菌具有選擇性凝集作用,可通過(guò)調(diào)理和介導(dǎo)血細(xì)胞發(fā)揮吞噬外來(lái)物質(zhì)的作用,能夠協(xié)同機(jī)體的其他免疫因子抵御外來(lái)病原入侵[7]。隨著甲殼類養(yǎng)殖的增加及病害的增多,對(duì)養(yǎng)殖對(duì)蝦、貝類以及鱟的凝集素研究已有較多報(bào)道[8-14],但對(duì)三疣梭子蟹凝集素的研究較少[15-16],而同屬蟹類的中華絨螯蟹(Eriocheir sinensis)、鋸緣青蟹(Scylla serrata)的報(bào)道很多[17-22]。本研究對(duì)三疣梭子蟹血清凝集素開(kāi)展相關(guān)研究,包括其血凝譜、凝集條件、影響因素及凝集素的鹽析純化等,旨在為深入研究三疣梭子蟹血清凝集素類型、分布、功能,闡明凝集素在三疣梭子蟹免疫中的作用機(jī)制提供基礎(chǔ)資料。
1材料與方法
1.1材料
1.1.1供試動(dòng)物及血液三疣梭子蟹取自浙江省寧波象山鑫億鮮活水產(chǎn)品有限公司,體質(zhì)量150 g左右;新西蘭大白兔、ICR小鼠購(gòu)自浙江省醫(yī)學(xué)院動(dòng)物中心;鯽魚(yú)、草魚(yú)、雞、中華鱉購(gòu)自浙江省湖州市農(nóng)貿(mào)市場(chǎng),人的A、B、O型血由湖州市中心血站提供。
1.1.2試驗(yàn)藥品及試劑N-乙酰基氨基葡萄糖(N-acetyl-D-glucosamine,GlcNAc)、N-乙酰氨基甘露糖(N-acetyl-D-mannosamine,ManNAc)、葡萄糖、木糖、蔗糖、海藻糖、甘露糖、乳糖、D-半乳糖、果糖均為Sigma進(jìn)口分裝。Alservers抗凝劑、pH值7.4 Tris-HCl(TBS-Ⅰ 含Ca2+,TBS-Ⅱ不含Ca2+)等按文獻(xiàn)[23]方法配制。
1.2方法
1.2.1三疣梭子蟹血清制備用剪刀剪斷三疣梭子蟹的步足關(guān)節(jié)柔軟處,收集血淋巴于玻璃平皿中,室溫靜置2~3 h,4 ℃ 冰箱過(guò)夜,將其分裝于離心管中,6 000 r/min 、4 ℃離心10 min,吸取上層血清,-80 ℃保存?zhèn)溆谩?/p>
1.2.2動(dòng)物血細(xì)胞制備以Alservers抗凝劑潤(rùn)洗注射器抽血,小鼠和中華鱉為心臟采血,兔為耳靜脈采血,雞為翅下靜脈采血,鯽魚(yú)和草魚(yú)為尾靜脈采血。抗凝血液保存于4 ℃冰箱中,用前3 000 r/min 離心3 min,取血細(xì)胞以0.85%生理鹽水洗3次后,配制成2%(體積比)血細(xì)胞懸液,用于凝集試驗(yàn)。
1.2.3血清凝集素凝集譜的測(cè)定在96孔微量血凝板中進(jìn)行測(cè)定血清凝集素凝集譜。每孔加入25 μL TBS-Ⅰ,取 25 μL 血清樣品進(jìn)行連續(xù)2倍稀釋,再加入25 μL 2%血細(xì)胞,室溫靜置1~2 h,按文獻(xiàn)[24]方法觀察記錄紅細(xì)胞凝集情況。
1.2.4鹽度對(duì)血凝效價(jià)的影響配制不同鹽度的TBS-Ⅰ(0.15~1.35 mol/L NaCl),各取25 μL加至微量板中,再以2倍梯度稀釋待測(cè)三疣梭子蟹血清樣品,加入相同體積的2%小鼠血細(xì)胞,按上述方法測(cè)定血清凝集效價(jià)。
1.2.5pH值對(duì)血凝效價(jià)的影響分別用1 mol/L HCl和 1 mol/L NaOH將TBS-Ⅰ的pH值調(diào)為3.0、4.0、5.0、6.0、70、7.4、8.0、9.0、10.0,取不同pH值的TBS-Ⅰ 25 μL分別加至微量板中,再以2倍梯度稀釋待測(cè)血清樣品,加入等量的2%小鼠血細(xì)胞,室溫靜置1~2 h,測(cè)定血清凝集效價(jià)。
1.2.6Ca2+、Mg2+、EDTA對(duì)血凝效價(jià)的影響分別以TBS-Ⅱ配制含20 mmol/L Ca2+、20 mmol/L Mg2+、20 mmol/L EDTA的3種溶液,另設(shè)1組對(duì)照,分別取上述4種溶液 25 μL 加至微量板中,再以2倍梯度稀釋待測(cè)三疣梭子蟹血清樣品,加入等體積2%小鼠血細(xì)胞,測(cè)定血清凝集效價(jià)。
1.2.7糖抑制試驗(yàn)分別以TBS-Ⅰ配制200 mmol/L葡萄糖、木糖、蔗糖、海藻糖、甘露糖、乳糖、D-半乳糖、果糖、GlcNAc溶液以及200、400 mmol/L ManNAc溶液,分別取各溶液25 μL加至微量板中,再以2倍梯度稀釋待測(cè)三疣梭子蟹血清樣品,先在37 ℃中溫育30 min,加入 等體積的2%小鼠血細(xì)胞,測(cè)定血清凝集效價(jià)。
1.2.8血清的飽和硫酸銨溶液分級(jí)沉淀取血清樣品 10 mL,用PBS溶液稀釋至2倍,置于電磁攪拌器上攪拌,逐滴加入飽和硫酸銨溶液6.7 mL,邊加邊攪拌,4 ℃沉淀過(guò)夜。第2天把沉淀溶液分裝于7 mL離心管,4 ℃、4 000 r/min離心30 min。將所得沉淀加入適量的PBS溶解,轉(zhuǎn)移到透析袋中,透析2 d(間隔6~8 h換1次PBS溶液),最后得到的溶液就是血清經(jīng)25%飽和硫酸銨沉淀的鹽析成分。然后根據(jù)離心后上清液的量,加入適量飽和硫酸銨溶液,用相同方法分別得到30%、40%、50%、60%的飽和硫酸銨沉淀的血清鹽析成分,用蛋白測(cè)定儀分別測(cè)定各組分蛋白濃度。
1.2.9不同鹽析組分的凝集特性及糖抑制特性分別按照“1.2.3”節(jié)、“1.2.7”節(jié)的方法測(cè)定飽和硫酸銨溶液沉淀的各鹽析組分對(duì)不同動(dòng)物血細(xì)胞凝集效價(jià)及其糖抑制特性。
1.2.10SDS-PAGE電泳按照Laemmli方法[25],灌注 1 mm 厚5%濃縮膠和12%分離膠的不連續(xù)SDS-PAGE膠,根據(jù)濃度稀釋各樣品,并分別加入4×上樣緩沖液,煮沸3~5 min,用微量取樣器取10~20 μg蛋白上樣,垂直電泳槽恒壓80 V電泳,待跑至分離膠后,再將電壓調(diào)至110 V。電泳結(jié)束后用考馬斯亮藍(lán)快速染色液染色蛋白條帶,觀察結(jié)果并拍照。
2結(jié)果與分析
2.1血清凝集素對(duì)動(dòng)物紅細(xì)胞的凝集譜
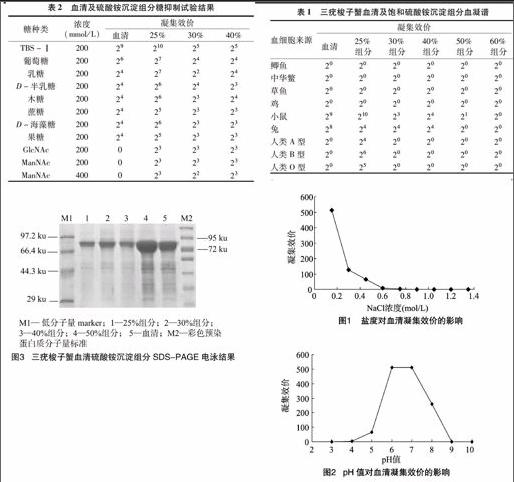
由表1可知,三疣梭子蟹血清凝集素對(duì)小鼠、兔、鯽魚(yú)、中華鱉、草魚(yú)、雞和人類A、B、O型血細(xì)胞均有不同程度凝集,其中以對(duì)小鼠血細(xì)胞的凝集效價(jià)最高,達(dá)210;其次是兔血細(xì)胞,
表1三疣梭子蟹血清及飽和硫酸銨沉淀組分血凝譜
2.2鹽度對(duì)血凝效價(jià)的影響
由圖1可知,在NaCl濃度是0.15 mol/L 時(shí)凝集活力最高;而隨著鹽度升高,凝集活力逐漸下降;當(dāng)鹽度達(dá)到 0.6 mol/L 以上時(shí),凝集活性被強(qiáng)烈抑制,凝集素已基本失去凝集活性。
2.3pH值對(duì)血凝效價(jià)的影響
由圖2可知,三疣梭子蟹血清在pH值為 6.0~7.4時(shí)凝集效價(jià)最高,為29;pH值低于6.0時(shí)凝集效價(jià)逐漸下降,pH值為4.0時(shí)凝集活性被強(qiáng)烈抑制,凝集素已基本失去凝集活性;pH值為8.0時(shí)凝集效價(jià)是28,pH值為9時(shí)凝集效價(jià)急劇下降,為20,基本處于失活狀態(tài)。
2.4Ca2+、Mg2+、EDTA對(duì)血凝效價(jià)的影響
測(cè)定了在三疣梭子蟹血清中分別添加Ca2+、Mg2+、EDTA對(duì)其凝集活力的影響,結(jié)果表明分別添加20 mmol/L Ca2+、20 mmol/L Mg2+均對(duì)其凝集活力具有增強(qiáng)作用,其凝集效價(jià)由28提高到29;但是添加20 mmol/L EDTA后,血清凝集效價(jià)降為23,幾乎失去活性。這表明EDTA能夠在血清環(huán)境下強(qiáng)烈抑制凝集素的凝集活性,三疣梭子蟹血清凝集素對(duì)Ca2+、Mg2+有明顯的依賴性。
2.5糖抑制試驗(yàn)結(jié)果
由表2可知,葡萄糖等9種常見(jiàn)糖對(duì)三疣梭子蟹血清的凝集活性有不同程度的抑制作用,其中N-乙酰基葡萄糖胺(GlcNAc)和N-乙酰甘露糖(ManNAc)對(duì)三疣梭子蟹凝集素的抑制作用最強(qiáng),200 mmol/L濃度即能完全抑制其凝集活性;其次為乳糖、D-半乳糖、果糖、木糖、蔗糖、D-海藻糖,200 mmol/L 葡萄糖對(duì)血清凝集素的抑制作用較小。從表2還可見(jiàn),各沉淀組分和GlcNAc或ManNAc結(jié)合時(shí),受到的抑制作用最強(qiáng)。表明三疣梭子蟹的血清凝集素對(duì)GlcNAc和ManNAc有特異性的結(jié)合能力。
表2血清及硫酸銨沉淀組分糖抑制試驗(yàn)結(jié)果
糖種類濃度
(mmol/L)凝集效價(jià)血清25%30%40%TBS-Ⅰ200292102525葡萄糖20026272424乳糖20024272224D-半乳糖20024262423木糖20024262324蔗糖20024252323D-海藻糖20024262323果糖20024252323GlcNAc2000232323ManNAc2000232323ManNAc4000232223
2.6飽和硫酸銨分級(jí)沉淀各組分的血凝譜
研究了25%、30%、40%、50%、60%飽和硫酸銨沉淀組分對(duì)動(dòng)物血細(xì)胞的凝集活性,結(jié)果發(fā)現(xiàn)25%沉淀組分的凝集活性最高,顯示三疣梭子蟹凝集素主要在25%硫酸銨飽和度析出。與血清的血凝譜相比,在30%、40%組分時(shí)凝集活性已經(jīng)較低,而到50%和60%組分時(shí)已經(jīng)基本沒(méi)有凝集活性(表1)。
2.7SDS-PAGE電泳結(jié)果
各鹽析組分樣品以10 μg蛋白上樣,用考馬斯亮藍(lán)快速染色液染色蛋白條帶。由圖3可見(jiàn),各泳道均有大量蛋白存在于72~95 ku,還須進(jìn)一步純化得到單一凝集素條帶。
3結(jié)論與討論
目前關(guān)于三疣梭子蟹凝集素特性的研究較少,僅有報(bào)道比較了包括三疣梭子蟹在內(nèi)的3種梭子蟹血清凝集素的細(xì)胞凝集活性[16]。本研究較為系統(tǒng)地研究了三疣梭子蟹血清凝集素的特性,結(jié)果表明:三疣梭子蟹血清凝集素對(duì)鯽魚(yú)、中華鱉、草魚(yú)、雞和人類A、B、O型血細(xì)胞沒(méi)有凝集作用,但是對(duì)小鼠、兔的血細(xì)胞具有較高的凝集活性,其中對(duì)小鼠血細(xì)胞的凝集活性高達(dá)210,這與梁青龍等的報(bào)道[16]一致,但該文沒(méi)有選擇小鼠紅細(xì)胞,測(cè)出兔細(xì)胞的凝集活性最高。本研究結(jié)果也與郝珂等報(bào)道的有關(guān)鋸緣青蟹血清凝集素的凝集特性[21-22]相似。因此,選擇小鼠紅細(xì)胞作為研究三疣梭子蟹血清凝集素特性的指示細(xì)胞,三疣梭子蟹凝集活性隨著鹽度的升高而降低,在鹽度大于0.6 mol/L時(shí)基本失活;凝集素作用的最適pH值為 6.0~7.4。已有研究表明,無(wú)脊椎動(dòng)物的血細(xì)胞凝集素在發(fā)揮凝集活動(dòng)作用時(shí),常須要一些二價(jià)金屬離子的參與[5],本研究選擇常見(jiàn)的二價(jià)金屬離子Ca2+、Mg2+,結(jié)果表明梭子蟹的血清凝集素對(duì)Ca2+、Mg2+有明顯依賴性,而且金屬離子螯合劑EDTA可以明顯抑制其凝集活性,說(shuō)明在三疣梭子蟹血清凝集素的凝集活動(dòng)中,二價(jià)金屬離子是必不可少的。
甲殼類動(dòng)物血清凝集素可與多種糖類發(fā)生特異性結(jié)合,凝集紅細(xì)胞的特性實(shí)際是和糖基位點(diǎn)發(fā)生作用,所以其結(jié)合也會(huì)被相應(yīng)的糖類所抑制。本研究分析了三疣梭子蟹血清凝集素對(duì)9種常見(jiàn)糖的敏感性,發(fā)現(xiàn)GlcNAc、ManNAc可特異性抑制血清凝集素的活性,表明梭子蟹凝集素的主要識(shí)別位點(diǎn)為GlcNAc、ManNAc,該結(jié)果與文獻(xiàn)報(bào)道的N-乙酰基糖類和唾液酸類糖蛋白是甲殼類動(dòng)物體內(nèi)凝集素具有特異性結(jié)合的主要2種糖基位點(diǎn)的結(jié)果[26]一致。
本研究采用(NH4)2SO4分級(jí)沉淀法對(duì)梭子蟹血清進(jìn)行了初步分離,發(fā)現(xiàn)其凝集素的活力區(qū)主要在25%飽和(NH4)2SO4區(qū),具有很高的凝集活性,并且對(duì)GlcNAc、ManNAc 有特異性結(jié)合。SDS-PAGE結(jié)果表明,純化蛋白集中存在于72 ~95 ku。50%飽和(NH4)2SO4區(qū)蛋白含量特別高,蛋白大小區(qū)域與25%飽和(NH4)2SO4區(qū)差不多,但其凝集活性很低,效價(jià)僅為21 ,推測(cè)該部分主要為梭子蟹的血藍(lán)蛋白成分。這與以往報(bào)道的蝦類血藍(lán)蛋白具有一定的凝集活性,可與脊椎動(dòng)物紅細(xì)胞產(chǎn)生凝集反應(yīng)的結(jié)果[27]有些不一致。本研究結(jié)果為進(jìn)一步分離純化提取三疣梭子蟹血清凝集素及研究其特性和功能奠定了良好基礎(chǔ)。
參考文獻(xiàn):
[1]Hamasaki K,F(xiàn)ukunaga K,Kitada S. Batch fecundity of the swimming crab Portunus trituberculatus (Brachyura: Portunidae)[J]. Aquaculture, 2006, 25(1): 359-365.
[2]王國(guó)良,金珊,李政,等. 三疣梭子蟹(Portunus trituberculatus)乳化病的組織病理和超微病理研究[J]. 海洋與湖沼,2006,37(4):297-303.
[3]劉淇,李海燕,王群,等. 梭子蟹牙膏病病原菌——溶藻弧菌的鑒定及其系統(tǒng)發(fā)育分析[J]. 海洋水產(chǎn)研究,2007,28(4):9-13.
[4]孫冊(cè),朱政,奠漢慶. 凝集素[M]. 北京:科學(xué)出版社,1986.
[5]Dodd R B,Drickamer K. Lectin-like proteins in model organisms: implications for evolution of carbohydrate-binding activity[J]. Glycobiology, 2001, 11(5): 71R-79R.
[6]Vasta G R, Ahmed H, Odom E W. Structural and functional diversity of lectin repertoires in invertebrates, protochordates and ectothermic ertebrates vertebrates[J]. Current Opinion in Structural Biology, 2004, 14(5): 617-630.
[7]陳皓文,孫丕喜,宋慶云. 外源凝集索——水產(chǎn)動(dòng)物御敵的有力兵器[J]. 黃渤海海洋,1995,13(3):61-70.
[8]彭其勝,郭文場(chǎng),楊振國(guó),等. 中國(guó)對(duì)蝦血淋巴液中的凝集素[J]. 中國(guó)水產(chǎn)科學(xué),2000,7(4):14-18.
[9]曹劍香,簡(jiǎn)紀(jì)常,吳灶和,等. 凡納濱對(duì)蝦血清凝集素、溶血素的特性研究[J]. 海洋科學(xué),2006,30(6):1-5.
[10]Wang H,Song L S,Li C H,et al. Cloning and characterization of a novel C-type lectin from Zhikong scallop Chlamys farreri[J]. Molecular Immunology,2007,44(5): 722-731.
[11]Wang X W,Xu W T,Zhang X W,et al. A C-type lectin is involved in the innate immune response of Chinese white shrimp[J]. Fish & Shellfish Immunology,2009,27(4): 556-562.
[12]Sun Y D,F(xiàn)u L D,Jia Y P,et al. A hepatopancreas-specific C-type lectin from the Chinese shrimp Fenneropenaeus chinensis exhibits antimicrobial activity[J]. Molecular Immunology,2008,45(2):348-361.
[13]Sun J,Wang L,Wang B J,et al. Purification and characterisation of a natural lectin from the serum of the shrimp Litopenaeus vannamei[J]. Fish & Shellfish Immunology,2007,23(2):292-299.
[14]Liu Y C,Li F H,Dong B,et al. Molecular cloning, characterization and expression analysis of a putative C-type lectin (Fclectin) gene in Chinese shrimp Fenneropenaeus chinensis[J]. Molecular Immunology,2007,44(4):598-607.
[15]Kong H J,Park E M,Nam B H,et al. A C-type lectin like-domain (CTLD)-containing protein (PtLP) from the swimming crab Portunus trituberculatus[J]. Fish & Shellfish Immunology,2008, 25(3):311-314.
[16]梁青龍,戴聰杰,陳寅山. 三種梭子蟹血清凝集素的細(xì)胞凝集活性比較[J]. 福建水產(chǎn),2005(2):45-50.
[17]曹廣力,朱越雄,薛仁宇,等. 中華絨螯蟹血清中外源凝集素的凝集作用及影響因素[J]. 水產(chǎn)養(yǎng)殖,1999(5):16-18.
[18]Zhang H,Chen L Q,Qin J,et al. Molecular cloning, characterization and expression of a C-type lectin cDNA in Chinese mitten crab, Eriocheir sinensis[J]. Fish & Shellfish Immunology,2011,31(2): 358-363.
[19]Na Y J,Kim Y J,Park B T,et al. A novel lectin isolated from the hemolymph of the Marine hair crab Erimacrus isenbeckii[J]. Protein and Peptide Letters,2007,14(8):800-803.
[20]戴聰杰,王桂忠,何劍鋒,等. 鋸緣青蟹(Scylla serrata)凝集素的分布及部分性質(zhì)研究[J]. 廈門大學(xué)學(xué)報(bào):自然科學(xué)版,2005,44(4):555-558.
[21]郝珂,錢冬,劉問(wèn),等. 鋸緣青蟹血清凝集素的特性和鹽析提取研究[J]. 淡水漁業(yè),2008,38(4):7-11.
[22]郝珂,錢冬,劉問(wèn),等. 鋸緣青蟹凝集素的提取及單克隆抗體制備[J]. 水產(chǎn)學(xué)報(bào),2009,33(4):590-596.
[23]奧斯伯F,布倫特R,金斯頓R E,等. 精編分子生物學(xué)實(shí)驗(yàn)指南[M]. 金由辛,譯. 北京:科學(xué)出版社,1999.
[24]戴華生. 新實(shí)驗(yàn)病毒學(xué)[M]. 北京:中國(guó)學(xué)術(shù)出版社,1983.
[25]汪家政,范明. 蛋白質(zhì)技術(shù)手冊(cè)[M]. 北京:科學(xué)出版社,2000.
[26]孫杰,安利國(guó),王雷,等. 甲殼綱動(dòng)物凝集素的結(jié)構(gòu)特征和分離純化方法[J]. 海洋科學(xué),2006,30(6):73-76.
[27]Zhang Y L,Wang S Y,Xu A L,et al. Affinity proteomic approach for identification of an IgA-like protein in Litopenaeus vannamei and study on its agglutination characterization[J]. Journal of Proteome Research,2006,5(4):815-821.
