基于SRAP分子標(biāo)記構(gòu)建的菊苣遺傳連鎖圖譜
2015-04-15 08:01:03梁小玉季楊白史且黃琳凱張新全
草業(yè)學(xué)報(bào) 2015年5期
梁小玉,季楊,白史且,黃琳凱,張新全*
(1.四川省畜牧科學(xué)研究院,四川 成都 610066;2. 四川省草原科學(xué)研究院,四川 成都 611731;3. 四川農(nóng)業(yè)大學(xué),四川 雅安 625014)
?
基于SRAP分子標(biāo)記構(gòu)建的菊苣遺傳連鎖圖譜
梁小玉1,季楊1,白史且2,黃琳凱3,張新全3*
(1.四川省畜牧科學(xué)研究院,四川 成都 610066;2. 四川省草原科學(xué)研究院,四川 成都 611731;3. 四川農(nóng)業(yè)大學(xué),四川 雅安 625014)
基于SRAP標(biāo)記,以遺傳關(guān)系和表型差異大的菊苣親本PI 651947和PI 652007雜交獲得的84個(gè)F1單株為作圖群體,進(jìn)行連鎖圖譜的構(gòu)建。采用Map Manager QTX b20軟件進(jìn)行連鎖分析,分別構(gòu)建了PI 651947和PI 652007的分子連鎖框架圖,共獲得77個(gè)SRAP標(biāo)記,其中父本遺傳圖譜涉及4個(gè)連鎖群,包含19個(gè)標(biāo)記,圖譜總長(zhǎng)為450.9 cM,標(biāo)記間平均圖距為23.7 cM。母本遺傳圖譜涉及13個(gè)連鎖群,包含58個(gè)標(biāo)記,圖譜總長(zhǎng)為1404.8 cM,標(biāo)記間平均圖距為24.2 cM。研究結(jié)果可為菊苣重要農(nóng)藝性狀QTL定位奠定基礎(chǔ),為菊苣分子育種研究提供了基礎(chǔ)信息。
菊苣;遺傳連鎖圖譜;SRAP分子標(biāo)記
菊苣(Cichoriumintybus)為菊科(Asteraceae)溫帶多年生草本植物,在食品、醫(yī)藥、畜牧業(yè)及化工等多個(gè)領(lǐng)域發(fā)揮重要的作用,是當(dāng)今世界最具發(fā)展?jié)摿Φ慕?jīng)濟(jì)植物之一[1]。菊苣是自然界中菊粉含量最高的植物,而菊粉是目前國(guó)際食品界受到高度重視的集醫(yī)療、保健于一體的優(yōu)秀的功能性食品基料[2-3]。菊苣不僅富含菊粉,而且具有高產(chǎn)、優(yōu)質(zhì)、高抗、適應(yīng)性范圍廣等優(yōu)點(diǎn),在美國(guó)、新西蘭等畜牧業(yè)發(fā)達(dá)國(guó)家是非常重要的優(yōu)質(zhì)牧草。因此,提高菊苣產(chǎn)量和菊粉含量是當(dāng)前菊苣研究和育種的首要目標(biāo)。菊苣產(chǎn)量和菊粉含量均是受多基因控制的復(fù)雜的經(jīng)濟(jì)數(shù)量性狀,遺傳基礎(chǔ)復(fù)雜,容易受遺傳背景和環(huán)境因子的共同影響。傳統(tǒng)遺傳改良方法進(jìn)展較慢,效率低,分子技術(shù)則是一種快速有效的改良數(shù)量性狀的遺傳改良方法。菊苣分子遺傳圖譜的構(gòu)建將有助于重要的農(nóng)藝性狀基因的定位和克隆[4],從而大幅度提高菊苣的育種水平和育種效率,為優(yōu)良新品種選育奠定基礎(chǔ)。
國(guó)外學(xué)者對(duì)菊苣遺傳圖譜構(gòu)建及QTL定位進(jìn)行了研究,迄今為止,已公布的菊苣圖譜僅有4張,而國(guó)內(nèi)尚未開(kāi)展這方面的研究。1997年Matteo等[5]采用單假測(cè)交方法,總計(jì)371個(gè)標(biāo)記(包括16個(gè)RAPD、283個(gè)AFLP和72個(gè)SAMPL)構(gòu)建了第1張菊苣分子遺傳連鎖圖。研究者采用菊苣和苦苣(Cichoriumendivia)種間雜交的雜合材料和苦苣種自交產(chǎn)生的純合材料為親本,構(gòu)建了46個(gè)單株的F1群體,最后獲得了13個(gè)連鎖群,圖譜總長(zhǎng)1330 cM。2010年,Cassan等[6]選擇營(yíng)養(yǎng)生長(zhǎng)期對(duì)N肥敏感差異很大的兩個(gè)食用型菊苣為親本構(gòu)建了302個(gè)重組自交系群體,采用兩種標(biāo)記繪制其遺傳連鎖圖譜,最終上圖標(biāo)記包括9個(gè)SSR和73個(gè)RAPD,標(biāo)記位點(diǎn)間平均圖距為13.5 cM,并進(jìn)行QTL定位分析。結(jié)果顯示,控制N轉(zhuǎn)化的基因?qū)哲狞S化芽球產(chǎn)量和品質(zhì)的性狀影響是截然相反的,與產(chǎn)量呈正相關(guān)而品質(zhì)反之。認(rèn)為通過(guò)分子標(biāo)記輔助選擇育種對(duì)農(nóng)藝性狀和生理性狀QTLs基因分析,可以幫助選擇適宜施N量獲得優(yōu)質(zhì)的菊苣黃化芽球。同年,Cadalen等[7]選擇2個(gè)工業(yè)菊苣和1個(gè)食用型菊苣為親本,基于SSR和STS總計(jì)734個(gè)標(biāo)記分別構(gòu)建了3個(gè)群體的3張遺傳連鎖圖。利用檢測(cè)到的193個(gè)“橋標(biāo)記”構(gòu)建了9個(gè)同源連鎖群,涉及472個(gè)標(biāo)記,覆蓋圖距為878 cM。2013年,Gonthier等[8]采用工業(yè)用菊苣品種處理后獲得的雄性不育的K28和花粉可育的K59 兩份材料為親本構(gòu)建了389個(gè)單株的F1群體,基于13個(gè)與雄性不育位點(diǎn)相關(guān),6個(gè)與自交不親和位點(diǎn)相關(guān)的AFLP 標(biāo)記轉(zhuǎn)化的8個(gè)SCAR標(biāo)記及256對(duì)AFLP引物構(gòu)建了高密度的菊苣遺傳連鎖圖譜。研究結(jié)果顯示,包含NMS1-locus和S-locus的染色體區(qū)域的圖距都局限于0.8 cM范圍內(nèi)。現(xiàn)有研究表明,菊苣遺傳圖譜構(gòu)建及QTL定位研究較少,遺傳研究相對(duì)滯后,而且主要是集中在以菊苣種和苦苣種為親本的食用型和工業(yè)型菊苣圖譜構(gòu)建,以飼用型菊苣為親本的菊苣圖譜尚未見(jiàn)報(bào)道,也未見(jiàn)基于SRAP標(biāo)記的菊苣遺傳圖譜構(gòu)建的報(bào)道。由于已構(gòu)建的菊苣圖譜較少,而且構(gòu)建圖譜的親本來(lái)源不同,涉及種間和種內(nèi)的雜交材料,很難將現(xiàn)有的圖譜信息進(jìn)行整合分析和利用,在一定程度上限制了菊苣分子生物學(xué)研究的進(jìn)程,制約了菊苣育種水平的提高。而SRAP是針對(duì)外顯子區(qū)域進(jìn)行擴(kuò)增的一種新型標(biāo)記,與RFLP、SSR、RAPD以及AFLP標(biāo)記相比,操作更簡(jiǎn)單、成本低廉、穩(wěn)定性好、遺傳多態(tài)性高,可直接切膠回收用于測(cè)序,便于克隆目標(biāo)片段,目前已經(jīng)被成功應(yīng)用于多種植物的圖譜構(gòu)建[4,9-15]。
本研究以飼用型菊苣種的栽培材料和野生材料為作圖親本,采用“雙假測(cè)交”方法,利用SRAP標(biāo)記構(gòu)建菊苣遺傳圖譜,為飼用菊苣產(chǎn)量及其相關(guān)重要農(nóng)藝性狀QTL定位、菊苣雜種優(yōu)勢(shì)機(jī)理等研究搭建一個(gè)良好的平臺(tái),同時(shí)也為菊苣及其近緣種比較分析奠定一定基礎(chǔ)。
1 材料與方法
1.1 F1作圖群體的構(gòu)建
菊苣材料由美國(guó)國(guó)家基因庫(kù)提供,選擇來(lái)源和農(nóng)藝性狀差異很大的兩份材料為作圖親本,母本PI 651947為源自北荷蘭的晚熟、飼用型栽培材料,直立株型、植株高大、葉片寬大、葉緣為全緣或者淺裂;父本PI 652007為源自波蘭彼得哥什的晚熟野生材料,蓮座期匍匐株型、植株低矮、葉片窄、葉緣為深裂[16](圖1)。2010年進(jìn)行人工雜交,雜交前分別針對(duì)父、母本,選擇健壯、飽滿即將張開(kāi)并已露色的花序除去周圍的小花序后套袋。觀察到父本花序有大量花粉時(shí)開(kāi)始雜交,直接從父本單株上剪下花序,然后輕輕把花粉抖落在母本花序上,授完粉之后,即刻套上雜交袋,并且掛上標(biāo)簽紙牌,注明雜交組合和授粉時(shí)間。母本植株雜交種子呈褐色時(shí)開(kāi)始收獲成熟種子。當(dāng)年將雜交種子在溫室大棚中利用育苗盤(pán)穴播育苗,9周后將F1群體移栽到試驗(yàn)基地中。提取單株DNA用于分子標(biāo)記雜交后代鑒定,真雜種用于F1作圖群體構(gòu)建。

圖1 父本PI 652007和母本PI 651947植株Fig.1 The plant male parental PI 652007 and female parental PI 651947 of parents
1.2 基因組DNA的提取及純度檢測(cè)
2011年3月,選取菊苣F1代雜種及親本材料單株健康幼嫩葉片采用CTAB方法提取基因組DNA[9]。利用0.8%瓊脂糖凝膠電泳和紫外分光光度計(jì)檢測(cè)DNA濃度和純度,合格的DNA稀釋成10 ng/μL后在-20℃冰箱內(nèi)保存?zhèn)溆谩?/p>
1.3 SRAP-PCR 反應(yīng)體系及擴(kuò)增程序
參照Li和Quiros[9],Mishra和Nishani[17]及Guo和Luo[18]發(fā)表的引物分別設(shè)計(jì)了SRAP標(biāo)記的20條上游和下游引物,由北京六合華大基因科技股份有限公司合成。PCR反應(yīng)體系為15 μL,包括1 μL DNA,0.3 μL(2.5 U/μL) Taq DNA 酶,0.8 μL 10 mmol/L上游引物,0.8 μL 10 mmol/L下游引物,2 μL 10×PCR buffer,2 μL 25 mmol/L Mg2+,2.4 μL 2.5 mmol/L dNTPs,5.7 μL無(wú)菌水。PCR反應(yīng)程序參考Li和Quiros[9]的方法進(jìn)行優(yōu)化,具體如下:94℃預(yù)變性4 min;94℃變性1 min,35℃退火1 min,72℃延伸1 min,共5個(gè)循環(huán);94℃變性1 min,50℃退火1 min,72℃延伸1 min,共35個(gè)循環(huán);72℃延伸10 min,4℃保存。PCR擴(kuò)增產(chǎn)物在6.0%聚丙烯酰胺凝膠中電泳,點(diǎn)樣6 μL,電泳緩沖液為1×TBE,穩(wěn)壓400 V,時(shí)間為2.0 h。電泳完成后銀染,最后用數(shù)碼相機(jī)拍照保存。
1.4 數(shù)據(jù)分析及遺傳圖譜構(gòu)建
采用“雙假側(cè)交”(two-way pseudo-testcross)策略,按照顯性標(biāo)記統(tǒng)計(jì)方法人工統(tǒng)計(jì)雙親中存在多態(tài)性以及在子代中發(fā)生分離的條帶。有帶記為“1”,無(wú)帶記為“0”,缺失或者無(wú)法判斷的記為“-”。根據(jù)孟德?tīng)柗蛛x規(guī)律,對(duì)F1群體所有條帶進(jìn)行卡方檢驗(yàn),符合1∶1分離比率的標(biāo)記用于遺傳圖譜的構(gòu)建,顯著水平為0.05。采用Map Manager QTX b20軟件進(jìn)行連鎖遺傳分析,分別構(gòu)建父本和母本的遺傳連鎖圖譜。構(gòu)圖概率值設(shè)為0.001 (LOD=3), 采用 Kosambi函數(shù)[19]將重組率轉(zhuǎn)換為圖距(cM),最后用MapDraw V 2.1軟件繪制遺傳連鎖圖譜。
2 結(jié)果與分析
2.1 菊苣雜種鑒定
從SRAP引物中隨機(jī)選取了40對(duì)引物對(duì)4個(gè)F1子代及其親本的多態(tài)性進(jìn)行擴(kuò)增,篩選出帶型清晰、穩(wěn)定性好的11對(duì)引物作為檢測(cè)菊苣雜交種純度的特異引物,分別為:E17M12,E6M12,E8M15,E10M15,E8M16,E6M17,E6M18,E4M19,E3M20,E7M20,E9M20。篩選出的引物在F1與其親本之間均呈多態(tài)性。對(duì)89個(gè)雜種的擴(kuò)增圖譜進(jìn)行分析,84個(gè)F1植株均具有雙親特征帶,鑒定為真雜種,5個(gè)只有母本特征帶則鑒定為假雜種。結(jié)果表明,菊苣自交率很小,為5.62%。其中,引物對(duì)E10M15鑒定效率最高,一次性能鑒定出69粒真雜種,達(dá)到真雜種比例的82.14%,E17M12鑒定效率最差,一次性只鑒定出28粒真雜種,僅占真雜種比例的33.33%。
2.2 引物篩選
用400對(duì)SRAP 引物組合對(duì)雙親進(jìn)行多態(tài)性篩選,共有384對(duì)引物能擴(kuò)增出清晰、明亮條帶,再分別對(duì)親本及89株F1代個(gè)體篩選,根據(jù)后代的分離情況,選出了擴(kuò)增穩(wěn)定、能夠用于圖譜構(gòu)建的SRAP 引物143對(duì),引物入選率為35.75%(圖2為引物E3M3對(duì)部分群體的標(biāo)記分離情況)。
2.3 標(biāo)記的多態(tài)性和偏分離分析
143對(duì)多態(tài)性引物在親本和F1群體間共擴(kuò)增出535條多態(tài)性條帶,主要分布在90~500 bp之間。平均每個(gè)組合產(chǎn)生3.74個(gè)標(biāo)記,最多達(dá)10個(gè)。經(jīng)卡方(χ2)測(cè)驗(yàn)(P=0.05),535個(gè)標(biāo)記中,偏離孟德?tīng)柗蛛x規(guī)律的標(biāo)記有121個(gè),占22.6 %,符合1∶1的標(biāo)記有216個(gè),符合3∶1的標(biāo)記有198個(gè)。利用1∶1的標(biāo)記作圖,共有56對(duì)引物的78個(gè)標(biāo)記位點(diǎn)用于遺傳圖譜構(gòu)建,其中59個(gè)標(biāo)記來(lái)自母本PI651947,19個(gè)標(biāo)記來(lái)自父本PI652007。
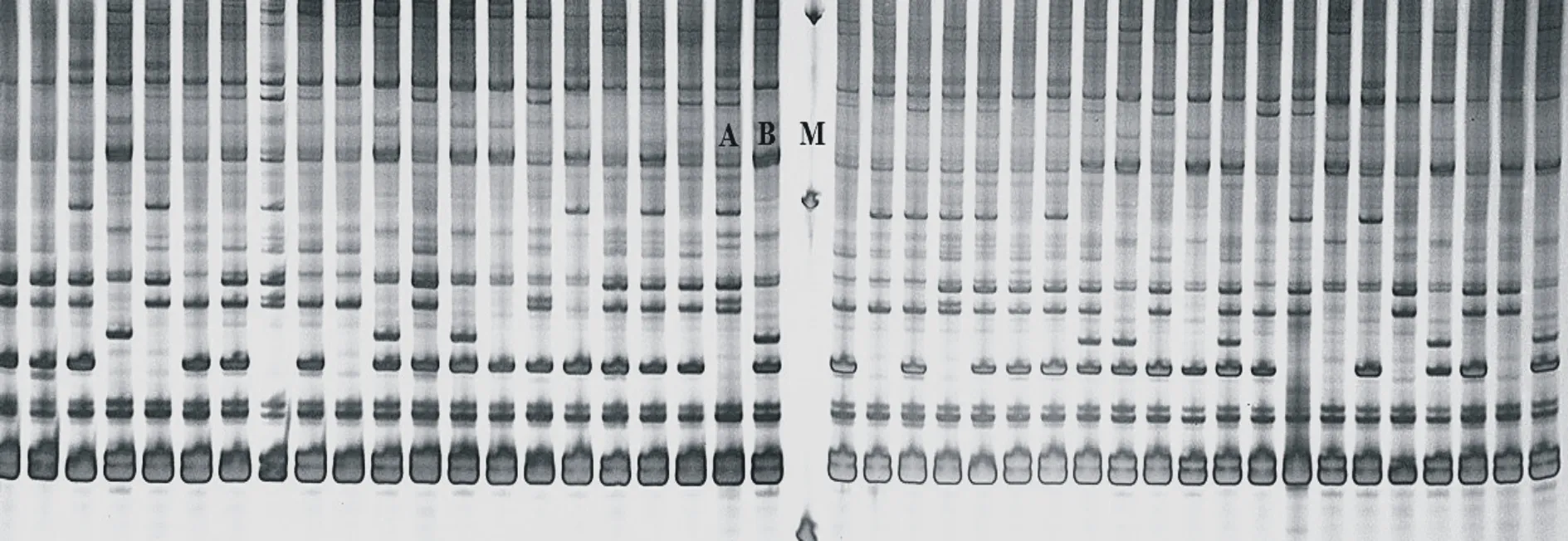
圖2 SRAP(E3M3)標(biāo)記在部分F1群體的分離 Fig.2 Segregation of SRAP(E3M3)marker in the partial F1 population A:PI651947; B:PI652007; M:標(biāo)記Marker;其他Others:F1個(gè)體 F1 individuals.
2.4 遺傳圖譜構(gòu)建
利用Map Manager QTX b20軟件對(duì)216個(gè)SRAP多態(tài)性標(biāo)記進(jìn)行連鎖遺傳分析(表1)。共有77個(gè)標(biāo)記進(jìn)入連鎖群,最終得到2張包含17個(gè)連鎖群的菊苣分子遺傳圖譜(圖3和4)。其中,父本的遺傳圖譜包含4個(gè)連鎖群(圖3),涉及19個(gè)SRAP標(biāo)記位點(diǎn),該圖譜總長(zhǎng)450.9 cM,標(biāo)記間平均圖距為23.7 cM,每個(gè)連鎖群長(zhǎng)度位于22.1~129.4 cM范圍內(nèi),標(biāo)記數(shù)介于2~8個(gè);母本的遺傳圖譜包含13個(gè)連鎖群(圖4),涉及58個(gè)SRAP標(biāo)記位點(diǎn),該圖譜基因組總長(zhǎng)度1404.8 cM,每個(gè)連鎖群的長(zhǎng)度位于22.9~480.4 cM范圍內(nèi),長(zhǎng)度最長(zhǎng)的連鎖群LG1為480.4 cM,有17個(gè)位點(diǎn)。每個(gè)連鎖群的標(biāo)記位點(diǎn)數(shù)2~17個(gè),每個(gè)標(biāo)記位點(diǎn)間平均圖距為24.2 cM。
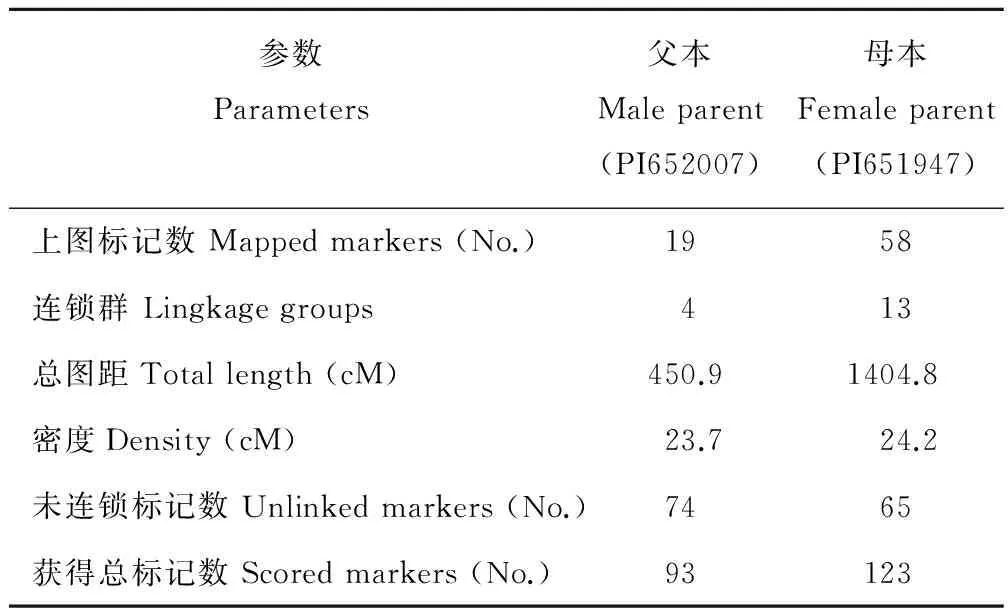
表1 用于連鎖分析的位點(diǎn)及連鎖分析結(jié)果

圖3 基于SRAP標(biāo)記構(gòu)建的菊苣父本(PI652007)遺傳連鎖圖譜Fig.3 PI 652007 genetic linkage map of chicory based on SRAP markers

圖4 基于SRAP標(biāo)記構(gòu)建的菊苣母本(PI 651947)遺傳連鎖圖譜Fig.4 PI 651947 genetic linkage map of chicory based on SRAP markers
3 討論
本研究采用SRAP標(biāo)記構(gòu)建菊苣遺傳連鎖圖譜,結(jié)果表明,SRAP標(biāo)記操作簡(jiǎn)單、經(jīng)濟(jì)成本低、擴(kuò)增譜帶清晰,多態(tài)性好,對(duì)于構(gòu)建高密度菊苣遺傳連鎖圖譜是有效的。
遺傳作圖群體的類型與群體大小直接關(guān)系到遺傳圖譜的精度和應(yīng)用范圍。菊苣是一個(gè)自然雜交的二倍體物種,存在嚴(yán)重自交不親和、花粉競(jìng)爭(zhēng)和雄性先熟等防止自花受粉的冗余系統(tǒng),具有高度雜合性,許多基因位點(diǎn)在F1代即發(fā)生分離,較難獲得純系,利用高世代群體構(gòu)建遺傳圖譜很難[20]。從目前已經(jīng)公布的4個(gè)二倍體菊苣分子遺傳圖譜看,群體樣本數(shù)在40~400以內(nèi),種間或種內(nèi)雜交所產(chǎn)生的F1代群體就會(huì)有高頻率的分離位點(diǎn),作圖群體主要是F1雜交群體。理論上本研究中選擇的親本為親緣關(guān)系遠(yuǎn)且多態(tài)性高的材料,獲得了84個(gè)F1真雜種單株群體,檢測(cè)到重組最小圖距為13.4 cM,表明親本和群體樣本數(shù)量上可以滿足構(gòu)建菊苣連鎖框架圖的需要。“雙擬測(cè)交”被認(rèn)為是解決多倍性異花授粉植物遺傳圖譜構(gòu)建的有效方法[21],該方法對(duì)F1群體作圖已經(jīng)被廣泛應(yīng)用于異花授粉植物中。與其他作圖群體比較,F(xiàn)1群體作圖不僅節(jié)約時(shí)間而且操作簡(jiǎn)單,也避免了自交不親和所造成的阻礙。因此,本研究采用“雙假側(cè)交”策略對(duì)F1群體進(jìn)行作圖,可視為回交(BC)群體模型來(lái)進(jìn)行圖譜構(gòu)建[22]。
本研究構(gòu)建了飼用菊苣的父本和母本2張遺傳連鎖圖譜,共包含了77個(gè)基于PCR擴(kuò)增技術(shù)的SRAP分子標(biāo)記。其中,母本圖譜包含13個(gè)連鎖群,涉及58個(gè)標(biāo)記,總圖距1404.8 cM,雖然上圖的標(biāo)記不多,但與Matteo等[5]構(gòu)建的連鎖圖譜的總圖距(1405 cM)一致,比較接近實(shí)際基因組,可以反映出雙親間遺傳距離大,差異位點(diǎn)多。父本圖譜僅包含了4個(gè)連鎖群,總圖距450.9 cM,遠(yuǎn)遠(yuǎn)小于已經(jīng)構(gòu)建的菊苣圖譜總長(zhǎng)度,主要原因可能是因?yàn)楦副緢D譜上圖標(biāo)記太少僅有19個(gè),因此對(duì)整個(gè)基因組的覆蓋率也較小。此外,父、母本圖譜上均存在較大的標(biāo)記空隙區(qū),連鎖群數(shù)目與染色體組的數(shù)目不一致,而通常情況下,分子連鎖群的數(shù)目與相應(yīng)物種染色體的數(shù)目是一致的。除了上圖標(biāo)記少外,分子標(biāo)記在染色體上分布的隨機(jī)性及染色體不同區(qū)段交換值的異質(zhì)性存在也會(huì)造成連鎖群上存在較大空隙,嚴(yán)重的則出現(xiàn)小片段的連鎖群[23]。
總的來(lái)看,總標(biāo)記數(shù)目相對(duì)較少、作圖軟件的局限性以及作圖群體較小是導(dǎo)致本研究所獲得的連鎖標(biāo)記不多圖距相對(duì)較大的主要原因,當(dāng)然,數(shù)據(jù)統(tǒng)計(jì)的失誤也可能造成標(biāo)記圖位差異的現(xiàn)象[24],有待于進(jìn)一步增加標(biāo)記數(shù)量,甚至利用不同分子標(biāo)記特性的互補(bǔ)來(lái)減小遺傳圖譜中的空隙[25]。
致謝:本試驗(yàn)曾得到蘭州大學(xué)謝文剛老師和四川農(nóng)業(yè)大學(xué)蔣林峰、汪霞、趙欣欣等同學(xué)的支持,特此致謝!
[1] Bais H P, Ravishankar G A.CichoriumintybusL. - cultivation, processing, utility, value addition and biotechnology, with an emphasis on current status and future prospects. Journal of the Science of Food and Agriculture, 2001, 81: 467-484.
[2] Kikuchi H, Inoue M, Saito H,etal. Industrial production of difructose anhydride III (DFA III) from crude inulin extracted from chicory roots using Arthrobacter sp H65-7 fructosyltransferase. Journal of Bioscience and Bioengineering, 2009, 107: 262-265.
[3] Park K J, Oliveira de R A, Brod F P R. Drying operational parameters influence on chicory roots drying and inulin extraction. Food and Bioproducts Processing, 2007, 85(3): 184-192.
[4] Jin M Y, Liu L Z, Fu F Y,etal. Construction of a genetic linkage map inBrassicanapusbased on SRAP, SSR, AFLP and TRAP. Molecular Plant Breeding, 2006, 4(4): 520-526.
[5] Matteo De S, Michele M, Margherita L,etal. A first linkage map ofCichoriumintybusL. using a one-way pseudo-testcross and PCR-derived markers. Molecular Breeding, 1997, 3: 415-425.
[6] Cassan L, Moreau L, Segouinc S,etal. Genetic map construction and quantitative trait loci (QTL) mapping for nitrogen use efficiency and its relationship with productivity and quality of the biennial crop Belgian endive (CichoriumintybusL.). Journal of Plant Physiology, 2010, 167: 1253-1263.
[7] Cadalen T, M?rchen C, Blassiau A,etal. Development of SSR markers and construction of a consensus genetic map for chicory (CichoriumintybusL.). Molecular Breeding, 2010, 25: 699-722.
[8] Gonthier L, Blassiau C, M?rchen M,etal. High-density genetic maps for loci involved in nuclear male sterility (NMS1) and sporophytic self-incompatibility (S-locus) in chicory (CichoriumintybusL., Asteraceae). Theoretical and Applied Genetics, 2013, 126: 2103-2121.
[9] Li G, Quiros C F. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica. Theoretical and Applied Genetics, 2001, 103: 455-461.
[10] Li Y Y, Shen J X, Wang T H,etalConstruction of a linkage map using SRAP, SSR and AFLP markers inBrassicanapusL. Scientia Agricultura Sinica, 2007, 40(6): 1118-1126.
[11] Ou C G, Deng B T, Bao S Y,etal. QTL mapping for contents of main carotenes and lycopene in carrot (DaucuscarotaL.). Hereditas (Beijing), 2010, 32(12): 1290-1295.
[12] Aneja B, Yadav N R, Chawla V, Yadav, R C. Sequence-related amplified polymorphism (SRAP) molecular marker system and its applications in crop improvement. Molecular Breeding, 2012, 30(4): 1635-1648.
[13] LI J Q, Wang L H, Zhan Q W,etal. Genetic diversity of 20 ryegrass accessions by SRAP markers . Acta Prataculturae Sinica, 2013, 22(2): 158-164.
[14] Gu X Y, Guo Z H, Zhang X Q,etal. Genetic diversity ofElymussibiricusgermplasm resources revealed by SRAP markers. Acta Prataculturae Sinica, 2014, 23(1): 205-216.
[15] Chen Q, Yuan X J, He Y L.Screening molecular markers for heat tolerance and its relation to summer tolerance in tall fescue single plants. Acta Prataculturae Sinica, 2013, 22(5): 84-95.
[16] Liang X Y, Zhang X Q, Bai S Q,etal. Multiple statistical analysis of the phenotypic characters ofCichoriumintybus.Acta Prataculturae Sinica, 2013, 22(6): 257-267.
[17] Mishra K M, Nishani S. Molecular identification and genetic relationships among coffee species (CoffeaL.) inferred from ISSR and SRAP marker analyses. Archives of Biological Sciences, 2011, 63: 667-679.
[18] Guo D L, Luo Z R. Genetic relationships of some pcna persimmons (Diospyroskakithunb.) from China and Japan revealed by SRAP analysis. Genetic Resources and Crop Evolution, 2006, 53: 1597-1603.
[19] Kosambi D D. The geometric method in mathematical statistics. The American Mathematical Monthly, 1944, 51(7): 382-389.
[20] Zheng Y Q, Liu J X. Advances in construction and application of turfgrass molecular genetic maps. Acta Prataculturae Sinica, 2009, 18(1): 155-162.
[21] Humphreys M W, Yadav R S, Cairns A J,etal. A changing climate for grassland research. New Phytologist, 2006, 169(1): 9-26.
[22] Yu C, Jin M Y, Zhang B Z,etal. Genetic linkage map of anthurium andraeanum based on SRAP molecular markers. Acta Horticulturae Sinica, 2012, 39(6): 1151-1158.
[23] Grattapaglia D, Sederoff R. Genetic linkage maps ofEucalyptusgrandisandEucalyptusurophyllausing a pseudo-testcross: Mapping strategy and RAPD markers. Genetics, 1994, 137: 1121-1137.
[24] Yang X, Yu Y J, Zhang F L,etal. Linkage map construction and quantitative trait loci analysis for bolting based on a double haploid population ofBrassicarapa. Journal of Intergrative Plant Biology, 2007, 49(5): 664-671.
[25] Bi C, Lu J N, Yin X G. Molecular linkage map constructed by SSR markers in castor. Journal of Inner Mongolia University for Nationalities(Natural Sciences), 2013, 28(5): 532-564.
參考文獻(xiàn):
[4] 金夢(mèng)陽(yáng), 劉列釗, 付福友, 等. 甘藍(lán)型油菜SRAP、SSR、AFLP和TRAP標(biāo)記遺傳圖譜構(gòu)建. 分子植物育種, 2006, 4(4): 520-526.
[10] 李媛媛, 沈金雄, 王同華, 等. 利用SRAP、SSR和AFLP標(biāo)記構(gòu)建甘藍(lán)型油菜遺傳連鎖圖譜. 中國(guó)農(nóng)業(yè)科學(xué), 2007, 40(6): 1118-1126.
[11] 歐承剛, 鄧波濤, 鮑生有, 等. 胡蘿卜(DaucuscarotaL.)中主要胡蘿卜素和番茄紅素含量的QTL分析. 遺傳, 2010, 32(12): 1290-1295.
[13] 李杰勤, 王麗華, 詹秋文, 等. 20個(gè)黑麥草品系的SRAP遺傳多樣性分析. 草業(yè)學(xué)報(bào), 2013, 22(2): 158-164.
[14] 顧曉燕, 郭志慧, 張新全, 等. 老芒麥種質(zhì)資源遺傳多樣性的SRAP分析. 草業(yè)學(xué)報(bào), 2014, 23(1): 205-216.
[15] 陳群, 袁曉君, 何亞麗. 高羊茅單株耐熱性相關(guān)分子標(biāo)記的篩選及其與越夏性的關(guān)系研究. 草業(yè)學(xué)報(bào), 2013, 22(5): 84-95.
[16] 梁小玉, 張新全, 白史且, 等. 菊苣主要表型性狀的多元統(tǒng)計(jì)分析. 草業(yè)學(xué)報(bào), 2013, 22(6): 257-267.
[20] 鄭軼琦, 劉建秀. 草坪草分子遺傳圖譜的構(gòu)建與應(yīng)用研究進(jìn)展. 草業(yè)學(xué)報(bào), 2009, 18(1): 155-162.
[22] 于翠, 金茂勇, 張寶珠, 等. 基于SRAP分子標(biāo)記的安祖花遺傳連鎖圖譜的構(gòu)建. 園藝學(xué)報(bào), 2012, 39(6): 1151-1158.
[25] 畢川, 陸建農(nóng), 殷學(xué)貴. 蓖麻遺傳圖譜構(gòu)建初報(bào).內(nèi)蒙古民族大學(xué)學(xué)報(bào)(自然科學(xué)版), 2013, 28(5): 532-564.
Construction of a genetic map for chicory usingsequence-related amplified polymorphism markers
LIANG Xiao-Yu1, JI Yang1, BAI Shi-Qie2, HUANG Lin-Kai3, ZHANG Xin-Quan3*
1.SichuanAcademyofAnimalSciences,Chengdu610066,China; 2.SichuanAcademyofGrasslandSciences,Chengdu611731,China; 3.DepartmentofGrasslandSciences,SichuanAgriculturalUniversity,Yaan625014,China
Sequence-related amplified polymorphism (SRAP) markers were used to construct a draft linkage map using an F1 population (84 individual plants) derived from a cross between PI 651947 and PI 652007; these lines possess different agronomic traits and DAN levels. Draft linkage maps of PI 651947 and PI 652007 were constructed using Map Manager QTX b20 software. 77 markers were used to construct parental linkage maps.The male map included 4 linkage groups with 19 molecular markers covering a total length of 450.9 centimorgans (cM). The average genetic distance between markers was 23.7 cM. The female map included 13 linkage groups with 58 molecular markers and covered a total length of 1404.8 cM. The average genetic distance between markers was 24.2 cM. This information established a foundation for extending genetic mapping in chicory and will serve as a framework for mapping QTLs and provide information required for further molecular studies.Key words: chicory (Cichoriumintybus); genetic linkage map; SRAP markers
10.11686/cyxb20150518
http://cyxb.lzu.edu.cn
2014-05-21;改回日期:2014-06-30
四川省畜牧科學(xué)研究院基本科研業(yè)務(wù)費(fèi)專項(xiàng)資金(SASA2014A02)和國(guó)家科技支撐計(jì)劃項(xiàng)目(2011BAD17B03)資助。
梁小玉(1976-),女,四川合江人,副研究員,博士。E-mail:liangxiaoyucao@163.com *通訊作者Corresponding author. E-mail: zhangxq@sicau.edu.cn
梁小玉, 季楊, 白史且, 黃琳凱, 張新全.基于SRAP分子標(biāo)記構(gòu)建的菊苣遺傳連鎖圖譜. 草業(yè)學(xué)報(bào), 2015, 24(5): 153-158.
Liang X Y, Ji Y, Bai S Q, Huang L K, Zhang X Q. Construction of a genetic map for chicory usingsequence-related amplified polymorphism markers. Acta Prataculturae Sinica, 2015, 24(5): 153-158.
