氮磷硅對(duì)咖啡雙眉藻和縊縮菱形藻繁殖速度和油脂積累的影響
2015-04-11 03:26:24王瑤華吳洪喜黃振華陳肖肖王召根
海洋科學(xué) 2015年4期
王瑤華, 吳洪喜 , 黃振華 , 陳肖肖 , 胡 園 , 王召根,
(1.上海海洋大學(xué) 水產(chǎn)與生命學(xué)院, 上海 201306; 2.浙江省海洋水產(chǎn)養(yǎng)殖研究所, 浙江 溫州 325005; 3.浙江省近岸水域生物資源開(kāi)發(fā)與保護(hù)重點(diǎn)實(shí)驗(yàn)室, 浙江 溫州 325005)
能源短缺已成為當(dāng)今世界面臨的三大問(wèn)題(人口膨脹, 環(huán)境污染, 能源短缺)之一。隨著石化能源的日益枯竭, 綠色、清潔生物能的開(kāi)發(fā)與利用已成為當(dāng)今各國(guó)政府和科學(xué)家關(guān)注的熱點(diǎn)。微藻因具有繁殖速度快、含油量高、單位面積產(chǎn)量高等特點(diǎn)被認(rèn)為是最具替代石化能潛力的新能源之一[1]。目前, 國(guó)內(nèi)外許多研究結(jié)果表明, 氮、磷、硅對(duì)微藻的繁殖速度和油脂積累有較大的影響[2-5], 但研究的對(duì)象主要是綠藻類和浮游硅藻類, 對(duì)底棲硅藻的研究還鮮見(jiàn)報(bào)道。本文以溫州洞頭海區(qū)分離出的 2種富油底棲硅藻咖啡雙眉藻(Amphora coffeaeformis)和縊縮菱形藻(Nitzschia constricta)為研究對(duì)象, 探討了氮、磷、硅對(duì)其繁殖速度和油脂積累的影響, 為底棲硅藻的能源開(kāi)發(fā)和利用積累基礎(chǔ)資料。
1 材料與方法
1.1 主要材料、儀器及試劑
藻種: 咖啡雙眉藻(Amphora coffeaeformis)、縊縮菱形藻(Nitzschia constricta), 均從洞頭自然海區(qū)中分離所得。
培養(yǎng)容器: 1000 mL錐形瓶。
主要儀器: 熒光分光光度計(jì)(型號(hào): 960CRT, 上海精密科學(xué)儀器有限公司生產(chǎn)); 可見(jiàn)分光光度計(jì)(型號(hào): 722型, 上海欣茂儀器有限公司生產(chǎn)); 智能光照培養(yǎng)箱(型號(hào): ZDX-150, 寧波海曙賽福實(shí)驗(yàn)儀器廠生產(chǎn)); 超聲清洗機(jī)(型號(hào): SB-5200DT, 寧波新芝生物科技股份有限公司生產(chǎn)); 立式壓力蒸汽滅菌器(型號(hào): LDZX型, 上海申安醫(yī)療器械廠生產(chǎn))。
主要試劑: NaNO3(分析純); NaH2PO4(分析純);NaSiO3(分析純);尼羅紅(Nile red)(純度≥95%, 生工生物工程(上海)股份有限公司生產(chǎn))。
1.2 方法
1.2.1 藻種的來(lái)源
采用水滴法和平板法從溫州洞頭海區(qū)掛板上的藻泥中分離出若干種底棲硅藻, 擴(kuò)大培養(yǎng)后測(cè)定其總脂含量, 選定含脂肪較高的咖啡雙眉藻和縊縮菱形藻為研究對(duì)象。
1.2.2 試驗(yàn)設(shè)計(jì)和培養(yǎng)基的制備
N、P、Si分別以f/2培養(yǎng)基為基礎(chǔ), 各設(shè)置5個(gè)質(zhì)量濃度水平, 分別為f/2培養(yǎng)基中相應(yīng)質(zhì)量濃度的1/7、1/5、1/3和9/7, 其他成分不變(表1), 各水平均設(shè)置2個(gè)重復(fù)。配置培養(yǎng)基用的海水取自當(dāng)?shù)睾^(qū),鹽度 30、pH 8.2, 并經(jīng) 0.45μm醋酸纖維膜過(guò)濾和120 ℃、0.1 MPa條件下滅菌20 min。
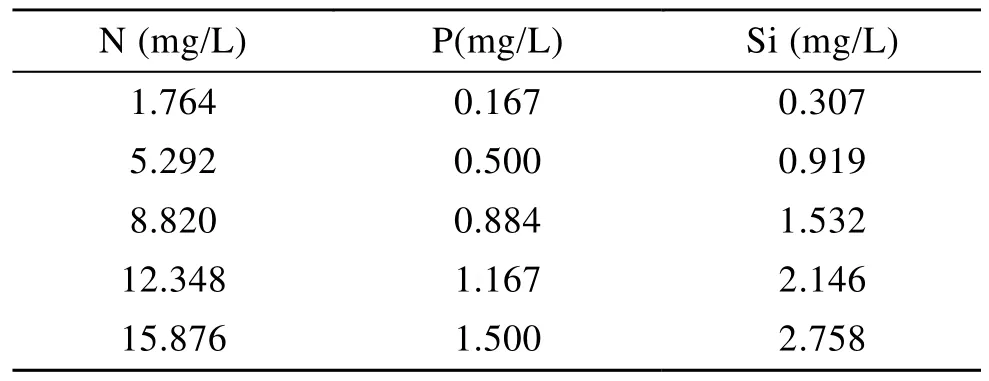
表1 不同N、P、Si質(zhì)量濃度的試驗(yàn)設(shè)計(jì)Tab.1 The experiment design with different concentrations of N, P and Si
1.2.3 尼羅紅染色液的配制
稱9 mg尼羅紅粉末溶于30 mL丙酮, 搖勻, 取1 mL混合液加到2 mL丙酮中, 即得到100μg/mL尼羅紅染色液, 4℃避光保存。
1.2.4 接種與培養(yǎng)管理
取對(duì)數(shù)生長(zhǎng)期的咖啡雙眉藻和縊縮菱形藻藻液離心濃縮, 用少量滅菌海水重懸, 取等量藻液分別接入按試驗(yàn)設(shè)計(jì)配置的不同質(zhì)量濃度的 N、P和 Si的培養(yǎng)基中。
控制試驗(yàn)溫度(25±1)℃, 鹽度 30, 藻液不通氣,所有試驗(yàn)均在光照培養(yǎng)箱中進(jìn)行, 光暗比12 h∶12 h。
1.2.5 工作曲線的制定
1.2.5.1 細(xì)胞密度測(cè)定工作曲線的制定
取一定量的咖啡雙眉藻和縊縮菱形藻置于超聲清洗儀中均勻分散, 用可見(jiàn)分光光度計(jì)測(cè)定其在 680 nm下的A680(光密度), 并用血球計(jì)數(shù)板計(jì)數(shù)出相應(yīng)的藻細(xì)胞密度, 得到A680與藻細(xì)胞密度的關(guān)系(圖1)和線性回歸方程:
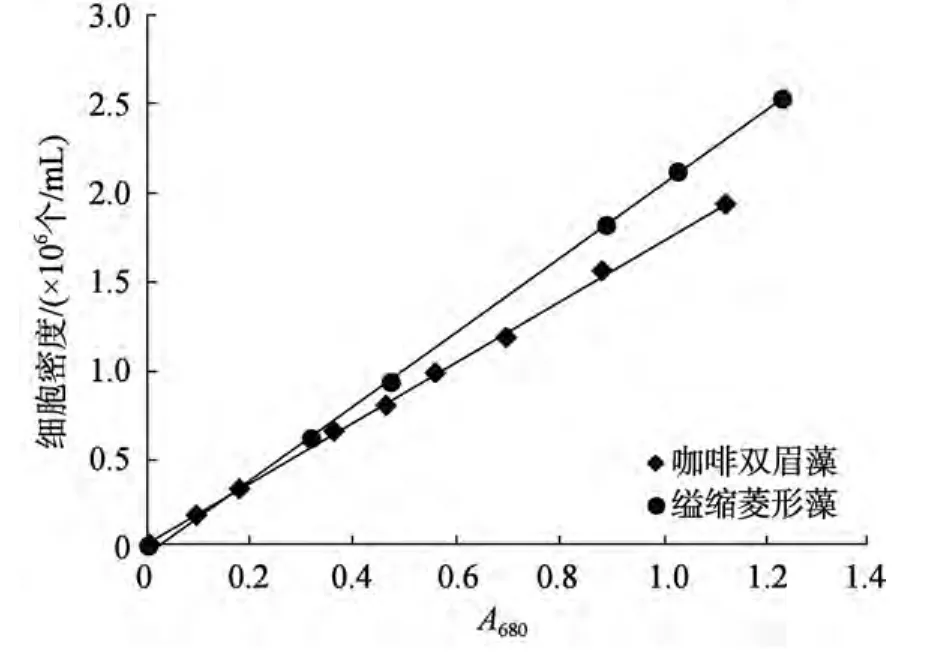
圖1 咖啡雙眉藻和縊縮菱形藻細(xì)胞密度與A680的關(guān)系Fig.1 The quantitative relationship between A680 and cell density of Amphora coffeaeformis and Nitzschia constricta
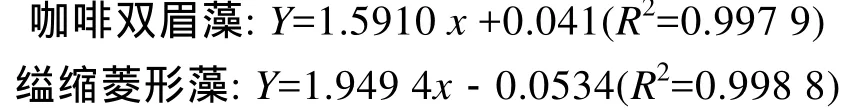
式中,Y為硅藻細(xì)胞密度(106個(gè)/mL);x為A680值。
兩者具有良好的線性關(guān)系, 本實(shí)驗(yàn) 2種底棲硅藻細(xì)胞密度由測(cè)得的A680代入上述相應(yīng)的公式(方程)求得。
1.2.5.2 油脂相對(duì)含量測(cè)定條件的確定
用熒光分光光度計(jì), 以 485 nm 為激發(fā)光波長(zhǎng),在500~750 nm范圍內(nèi)分別對(duì)海水、海水+尼羅紅、藻液、藻液+尼羅紅進(jìn)行掃描, 由于海水自身在這個(gè)范圍內(nèi)沒(méi)有自發(fā)熒光, 所以海水的發(fā)射光譜幾乎為一條直線; 海水+NR溶液在650 nm處有1個(gè)峰值,為尼羅紅在水中的自發(fā)熒光, 尼羅紅在水中溶解度很低, 故峰值較小; 藻液和藻液+NR的發(fā)射光譜在680 nm左右都有 1個(gè)峰值, 為葉綠素的發(fā)射峰; 藻液+NR在520~650 nm處出現(xiàn)1個(gè)很高的峰, 對(duì)比藻液的發(fā)射光譜, 可以推測(cè)出575 nm處為底棲硅藻的脂發(fā)射峰(圖2)。
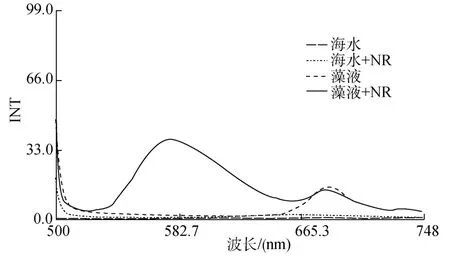
圖2 海水和藻液及其被尼羅紅染色后的發(fā)射光譜掃描圖Fig.2 Fluorescence emission spectra of seawater and microalgel before and after dyeing
向等體積、同密度的藻液中加入不同質(zhì)量濃度(0.2、0.4、0.6、0.8、1.0、1.2、1.4μg/mL)的尼羅紅,進(jìn)行熒光檢測(cè)。結(jié)果顯示尼羅紅在1.0 μg/mL質(zhì)量濃度時(shí)獲到最大的熒光強(qiáng)度, 高于這一質(zhì)量濃度,熒光強(qiáng)度反而下降。取一定量藻液, 稀釋成一定的密度梯度, 染色后檢測(cè)其熒光強(qiáng)度。結(jié)果顯示: 當(dāng)A680<0.686時(shí), 藻細(xì)胞密度與相對(duì)脂熒光強(qiáng)度的相關(guān)系數(shù)最大。底棲硅藻具有易沉底的特性, 選用偏低的細(xì)胞密度進(jìn)行熒光檢測(cè)以減小誤差。
1.2.6 測(cè)定方法
1.2.6.1 硅藻藻細(xì)胞密度的測(cè)定
隔天將培養(yǎng)容器和藻液在超聲清洗機(jī)中充分震蕩, 均勻分散后, 取 3 mL藻液用分光光度計(jì)測(cè)定680 nm處的A680。根據(jù)A680和已經(jīng)求出的工作曲線,計(jì)算出相應(yīng)的硅藻細(xì)胞密度。
1.2.6.2 油脂相對(duì)含量的測(cè)定
隔天將培養(yǎng)容器和藻液在超聲清洗機(jī)中充分震蕩, 均勻分散后取3 mL藻液測(cè)定其在染色前后485 nm激發(fā)波長(zhǎng)下 575 nm處的熒光強(qiáng)度(尼羅紅質(zhì)量濃度1μg/mL)。相對(duì)熒光強(qiáng)度=熒光強(qiáng)度/細(xì)胞密度; 熒光強(qiáng)度=藻液染色后熒光強(qiáng)度-藻液染色前熒光強(qiáng)度-尼羅紅的自發(fā)熒光強(qiáng)度(均為 575 nm處的熒光強(qiáng)度)。
1.2.7 數(shù)據(jù)處理
用 SPSS 15.0統(tǒng)計(jì)軟件對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行單因素方差分析(One-way ANOVA), 數(shù)據(jù)統(tǒng)計(jì)結(jié)果表示為平均值±標(biāo)準(zhǔn)差。
2 結(jié)果
2.1 氮對(duì) 2種底棲硅藻的繁殖和油脂含量的影響
試驗(yàn)結(jié)果顯示: NO3-N質(zhì)量濃度為1.746 mg/L時(shí),咖啡雙眉藻和縊縮菱形藻都幾乎不繁殖, 且藻細(xì)胞從實(shí)驗(yàn)的第7 天開(kāi)始發(fā)白、衰亡。隨著NO3-N質(zhì)量濃度的升高, 2種底棲硅藻的繁殖速度加快, 但當(dāng)質(zhì)量濃度達(dá)到8.820 mg/L后, 再繼續(xù)升高, 2種底棲硅藻的繁殖速度并不明顯加快(圖3和圖4)。從曲線可以看出 2種藻對(duì)氮質(zhì)量濃度的適應(yīng)范圍很相似, 只是咖啡雙眉藻對(duì)低氮的適應(yīng)力稍強(qiáng)于縊縮菱形藻。方差分析表明: 15d時(shí), 培養(yǎng)液中NO3-N質(zhì)量濃度對(duì)2種底棲硅藻繁殖有顯著的影響(P<0.05)。
實(shí)驗(yàn)表明, 不同NO3-N質(zhì)量濃度下, 2種底棲硅藻的油脂含量均呈先降后升, 到指數(shù)生長(zhǎng)末期后開(kāi)始大量積累。實(shí)驗(yàn)結(jié)束時(shí)(15d), 藻細(xì)胞油脂含量除質(zhì)量濃度最低組(1.746 mg/L)沒(méi)有升高, 甚至降低外,其他組都有所升高, 但質(zhì)量濃度越高, 油脂含量越低(脂熒光強(qiáng)度越低)。15.876 mg/L質(zhì)量濃度組只有5.920 mg/L 質(zhì)量濃度組的一半(圖3和圖4)。第7 天時(shí)顯微觀察顯示, 大部分底棲硅藻細(xì)胞已經(jīng)死亡,但并未解體, 仍有部分油滴被包裹在硅質(zhì)的細(xì)胞壁內(nèi), 所以雖然細(xì)胞密度下降, 其相對(duì)熒光強(qiáng)度并未快速下降。方差分析表明: 實(shí)驗(yàn)結(jié)束時(shí), 培養(yǎng)液中NO3-N質(zhì)量濃度對(duì) 2種底棲硅藻油脂含量有顯著影響(P<0.05)。
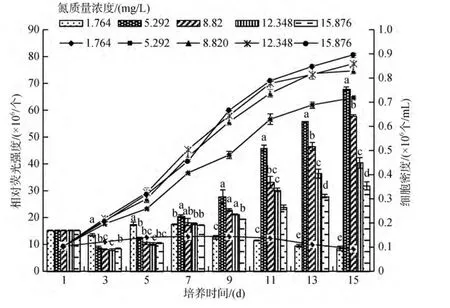
圖3 不同NO3-N質(zhì)量濃度下咖啡雙眉藻的繁殖曲線和油脂積累比較Fig.3 The growth curves and lipid accumulation of Amphora coffeaeformis under different NO3-N concentrations
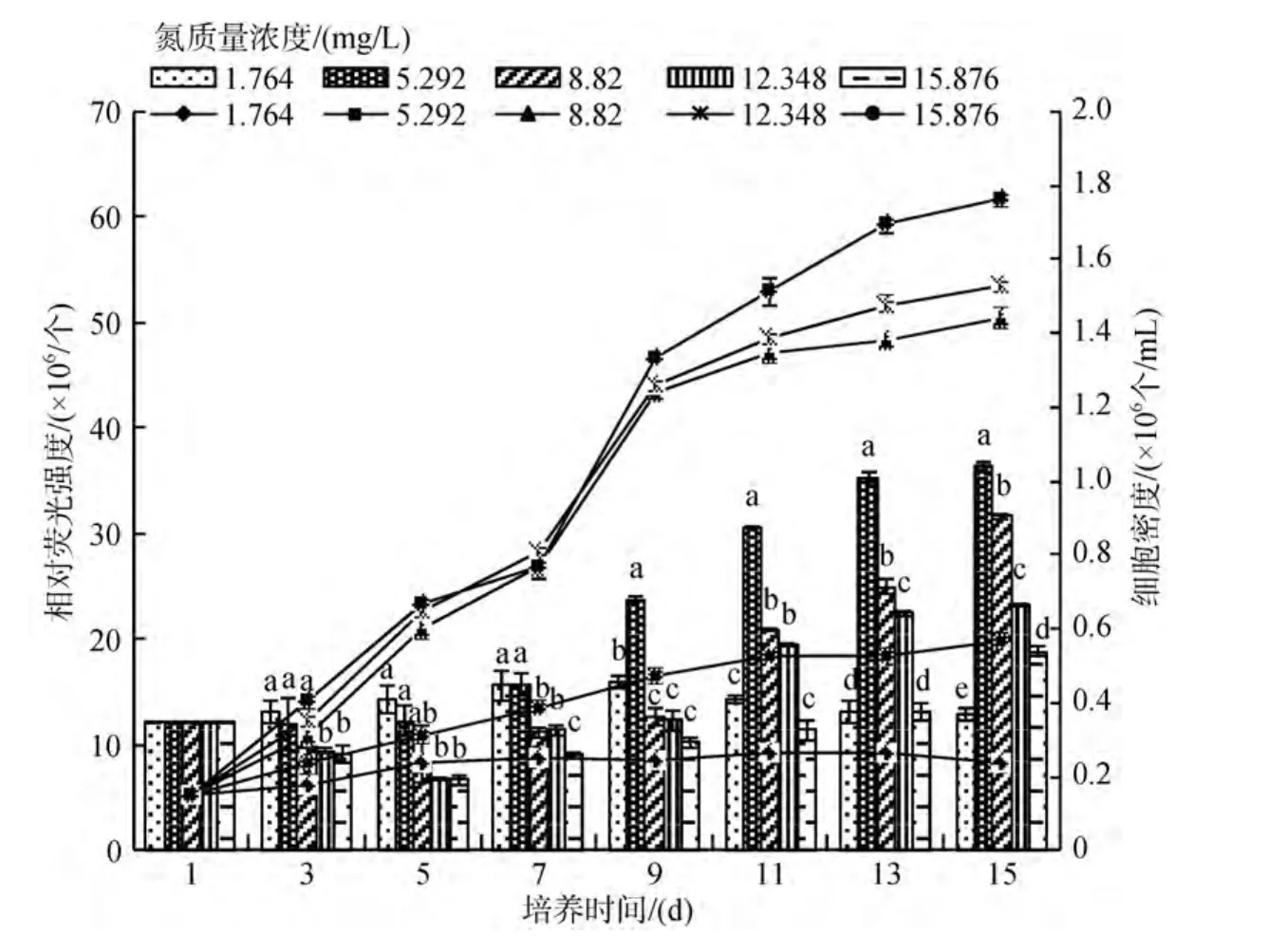
圖4 不同NO3-N質(zhì)量濃度下縊縮菱形藻的繁殖曲線和油脂積累比較Fig.4 The growth curves and lipid accumulation of Nitzschia constricta under different NO3-N concentrations
2.2 磷對(duì) 2種底棲硅藻的繁殖和油脂含量的影響
結(jié)果顯示, 在實(shí)驗(yàn)設(shè)計(jì)的 H2PO4-P質(zhì)量濃度范圍內(nèi), 2種底棲硅藻均能正常生長(zhǎng), 藻細(xì)胞繁殖速度均隨H2PO4-P質(zhì)量濃度的增高而加快, 但到了較高質(zhì)量濃度(0.884 mg/L)后, 藻細(xì)胞的繁殖速度不再明顯加快, 從生長(zhǎng)曲線可以看出, 咖啡雙眉藻較縊縮菱形藻更能適應(yīng)低磷環(huán)境(圖5和圖6), 培養(yǎng)液H2PO4-P質(zhì)量濃度對(duì)2種底棲硅藻繁殖有顯著影響(P<0.05)。
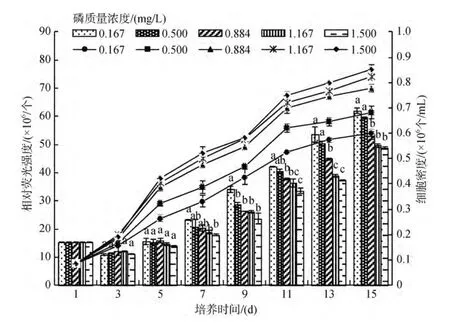
圖5 不同H2PO4-P質(zhì)量濃度下咖啡雙眉藻的繁殖曲線和油脂積累Fig.5 The growth curves and lipid accumulation of Amphora coffeaeformis under different H2PO4-P concentrations
隨著H2PO4-P質(zhì)量濃度的增高, 2種底棲硅藻細(xì)胞的油脂含量均呈現(xiàn)先降后升的變化規(guī)律, 且低質(zhì)量濃度的磷更有利于2種底棲硅藻細(xì)胞脂肪的積累。實(shí)驗(yàn)結(jié)束(15 d)時(shí), 咖啡雙眉藻和縊縮菱形藻細(xì)胞的油脂含量, 低磷質(zhì)量濃度(0.164 mg/L)組分別是最高磷質(zhì)量濃度(1.500 mg/L)組的1.3倍和1.6倍(圖5和圖6), 低質(zhì)量濃度磷對(duì)縊縮菱形藻油脂的影響大于咖啡雙眉藻。培養(yǎng)液中H2PO4-P質(zhì)量濃度對(duì)2種底棲硅藻油脂含量的影響, 除高于 0.884 mg/L質(zhì)量濃度組外, 其余均呈顯著影響(P<0.05)。
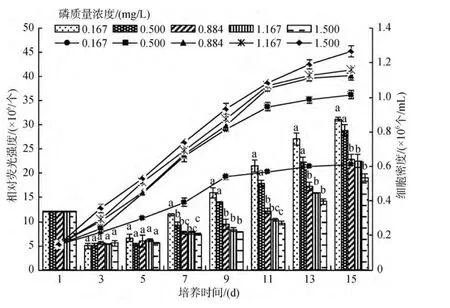
圖6 不同H2PO4-P質(zhì)量濃度下縊縮菱形藻的繁殖曲線和油脂積累Fig.6 The growth curves and lipid accumulation of Nitzschia constricta under different H2PO4-P concentrations
2.3 硅對(duì) 2種底棲硅藻的繁殖和油脂積累的影響
硅對(duì) 2種硅藻的生長(zhǎng)情況影響相似, 在實(shí)驗(yàn)設(shè)計(jì)的質(zhì)量濃度范圍內(nèi), SiO3-Si質(zhì)量濃度對(duì) 2種底棲硅藻繁殖速度的影響均隨質(zhì)量濃度的增加而升高(圖7和圖8), 且呈顯著性(P<0.05), 但當(dāng)質(zhì)量濃度達(dá)1.532 mg/L后, 繼續(xù)升高對(duì)2種底棲硅藻細(xì)胞繁殖速度的影響變?yōu)椴伙@著(P>0.05)。實(shí)驗(yàn)后期發(fā)現(xiàn), 低質(zhì)量濃度組硅藻細(xì)胞較高質(zhì)量濃度組硅藻細(xì)胞先老化,可見(jiàn), 硅有利于硅藻繁殖周期的延長(zhǎng)。
硅對(duì)2種底棲硅藻油脂積累影響與氮、磷相似,隨著SiO3-Si質(zhì)量濃度的增高, 藻細(xì)胞脂肪含量也呈升高→降低→升高的規(guī)律變化, 較低質(zhì)量濃度的硅(低于0.919 mg/L)更有利于硅藻細(xì)胞油脂的積累。在實(shí)驗(yàn)設(shè)計(jì)的質(zhì)量濃度范圍內(nèi), 實(shí)驗(yàn)結(jié)束(15 d)時(shí), 咖啡雙眉藻和縊縮菱形藻的油脂含量, 最低硅質(zhì)量濃度組(0.307mg/L)分別是最高硅質(zhì)量濃度組(2.758 mg/L)的1.3倍和1.5倍(圖7和圖8), 低質(zhì)量濃度的SiO3-Si更有利于2種底棲硅藻的油脂積累。方差分析表明: 實(shí)驗(yàn)結(jié)束(15d)時(shí), SiO3-Si對(duì)2種底棲硅藻油脂含量的影響, 質(zhì)量濃度低于1.532 mg/L影響顯著(P<0.05),質(zhì)量濃度高于1.532 mg/L影響不顯著(P>0.05)。

圖7 不同SiO3-Si質(zhì)量濃度下咖啡雙眉藻的繁殖曲線和油脂積累Fig.7 The growth curves and lipid accumulation of Amphora coffeaeformis under different SiO3-Si concentrations
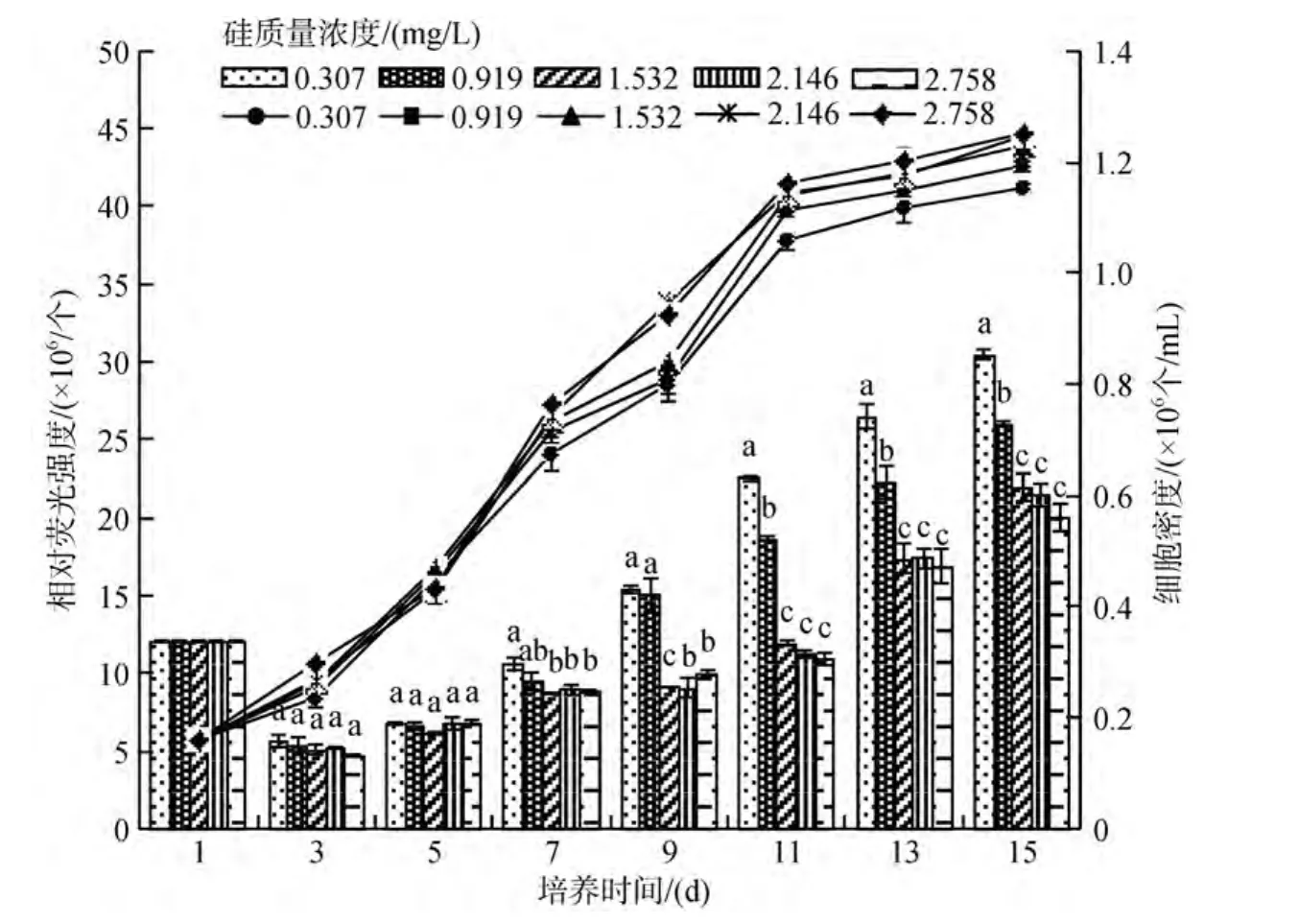
圖8 不同SiO3-Si質(zhì)量濃度下縊縮菱形藻的繁殖曲線和油脂積累Fig.8 The growth curves and lipid accumulation of Nitzschia constricta under different SiO3-Si concentrations
3 討論
3.1 環(huán)境脅迫可能是促進(jìn)硅藻細(xì)胞積累油脂的直接原因
添加不同質(zhì)量濃度的N、P、Si營(yíng)養(yǎng)培養(yǎng)實(shí)驗(yàn)表明, 縊縮菱形藻和咖啡雙眉藻的相對(duì)油脂含量總體呈現(xiàn)先降低后升高的趨勢(shì)。前期油脂含量的降低可能是由為硅藻細(xì)胞進(jìn)入新的環(huán)境, 需要一個(gè)適應(yīng)的過(guò)程, 到指數(shù)生長(zhǎng)期或更遲, 培養(yǎng)基中的主要營(yíng)養(yǎng)元素, 即將被消耗盡, 于是硅藻細(xì)胞開(kāi)始積累油脂。N、P和Si越早被消耗完, 硅藻細(xì)胞越早開(kāi)始積累油脂, 其油脂積累也越多。可見(jiàn), 不利的環(huán)境條件, 尤其是營(yíng)養(yǎng)鹽的缺乏, 更有利于硅藻細(xì)胞油脂的積累。
3.2 氮質(zhì)量濃度對(duì)縊縮菱形藻和咖啡雙眉藻的繁殖和油脂含量的影響
N對(duì)底棲硅藻繁殖速度的影響已見(jiàn)不少報(bào)道[6-7]。本實(shí)驗(yàn)結(jié)果表明, 咖啡雙眉藻繁殖適宜的氮質(zhì)量濃度為5.920~8.82 mg/L, 較李雅娟等[6](咖啡雙眉藻最適氮質(zhì)量濃度為2.5~5 mg/L)和王起華等[8](咖啡雙眉藻最適氮質(zhì)量濃度為1.54~6.16 mg/L)的研究結(jié)果高,這可能是由于本實(shí)驗(yàn)的藻種分離于營(yíng)養(yǎng)鹽較高的海區(qū), 該藻株產(chǎn)生了耐高營(yíng)養(yǎng)鹽特性; 縊縮菱形藻適宜的氮質(zhì)量濃度為 8.82 ~15.876 mg/L, 較董金利報(bào)道[9]的縊縮菱形藻在>12.35 mg/L才能繁殖要低, 這可能是因?yàn)槎鹄?.23 ~12.35 mg/L區(qū)間內(nèi)沒(méi)有設(shè)置實(shí)驗(yàn)組, 影響了實(shí)驗(yàn)精度。氮是構(gòu)成蛋白質(zhì)的必要成分, 在缺氮情況下藻細(xì)胞形成的蛋白質(zhì)量減少,硅藻光合作用形成的能量物質(zhì)有可能更多地轉(zhuǎn)向合成油脂[10]。Chelf[11]認(rèn)為不同的氮源質(zhì)量濃度對(duì)油脂積累有很大影響, 且目前普遍認(rèn)為氮缺乏會(huì)導(dǎo)致微藻細(xì)胞內(nèi)大量積累油脂[12-13], 本實(shí)驗(yàn)也印證了這一規(guī)律, 培養(yǎng)液中不同氮質(zhì)量濃度對(duì) 2種底棲硅藻繁殖和油脂含量的影響均較大, 適當(dāng)降低培養(yǎng)基中氮質(zhì)量濃度有利于油脂的積累。
3.3 磷質(zhì)量濃度對(duì) 2種底棲硅藻的繁殖和油脂含量的影響
2種底棲硅藻繁殖速度隨培養(yǎng)基中磷質(zhì)量濃度的增高而加快, 在 0.884~1.500 mg/L范圍內(nèi)均能正常繁殖, 這一現(xiàn)象符合藻類利用磷的特點(diǎn), 大多數(shù)微藻能夠主動(dòng)吸收磷, 當(dāng)環(huán)境磷充足時(shí), 細(xì)胞將吸收的多于生理需要的磷儲(chǔ)存于細(xì)胞內(nèi), 環(huán)境缺磷時(shí)又可利用這些儲(chǔ)存的磷進(jìn)行代謝[14]。一些藻類磷缺乏也能提高其油脂含量, 如三角褐指藻(Phaeodactylum tricornutumBohlin)、角毛藻(Chaetoceros)、巴夫藻(Pavlova viridis)等; 而另一些藻類磷缺乏反而導(dǎo)致油脂含量的降低, 如屠夫綠球藻(Nannochloris atomus)和融合微藻(Tetraselmissp.)[15]。本實(shí)驗(yàn)的咖啡雙眉藻和縊縮菱形藻屬于前者, 但磷缺乏較氮缺乏對(duì)藻細(xì)胞油脂積累的影響要小。隨著磷質(zhì)量濃度的降低, 藻細(xì)胞中油脂的含量回升越早, 這可能是因?yàn)榈统跏假|(zhì)量濃度的磷可以較早地激活油脂合成途徑。
3.4 硅質(zhì)量濃度對(duì) 2種底棲硅藻的繁殖和油脂積累的影響
本實(shí)驗(yàn)中, 硅質(zhì)量濃度在所設(shè)范圍內(nèi), 2種底棲硅藻均能正常繁殖, 但差異較小, 硅質(zhì)量濃度的增加沒(méi)有明顯地促進(jìn) 2種底棲硅藻的繁殖, 且低初始硅質(zhì)量濃度對(duì) 2個(gè)藻株繁殖的影響沒(méi)有低初始氮、磷質(zhì)量濃度的顯著。董金利[9]、馬志珍等[16]、王淵源等[17]報(bào)道在實(shí)驗(yàn)室內(nèi)用自然海水培養(yǎng)底棲硅藻, 即使不加硅鹽仍能良好繁殖。大貝政治等采用營(yíng)養(yǎng)去除法的實(shí)驗(yàn)發(fā)現(xiàn), 用不含硅的人工海水培養(yǎng)基培養(yǎng)一種卵形藻時(shí), 對(duì)其生長(zhǎng)的抑制作用很小。本實(shí)驗(yàn)中低硅質(zhì)量濃度組獲得較高的油脂相對(duì)含量, 這是因?yàn)楣枞狈?huì)引起油脂合成過(guò)程中起作用的乙酰-CoA羧化酶活力的增強(qiáng)[10], 低初始硅質(zhì)量濃度對(duì) 2個(gè)藻株繁殖的影響沒(méi)有低初始氮、磷質(zhì)量濃度影響的顯著(PN 本實(shí)驗(yàn)僅研究了氮、磷、硅單因子對(duì)硅藻細(xì)胞油脂積累的影響, 事實(shí)上營(yíng)養(yǎng)鹽的配比(主要是氮磷比)對(duì)藻類的代謝產(chǎn)物和繁殖速率也有顯著的影響[7,19-20],因此多因子組合對(duì) 2種底棲硅藻繁殖速度和油脂積累影響還需要進(jìn)一步深入實(shí)驗(yàn)和探討。 [1] Yusuf C.Biodiesel from microalgae[J].Biotechnology Advances, 2007, 25(3): 294-306. [2] 王曉青, 王嫁, 江雪, 等.三角褐指藻(Phaeodactylum tricornutum)油脂富集培養(yǎng)的研究[J].四川大學(xué)學(xué)報(bào)(自然科學(xué)版), 2011, 48(3): 691-696. [3] 魏東, 張學(xué)成.氮源和 N/P對(duì)眼點(diǎn)擬微綠球藻的生長(zhǎng)、總脂含量和脂肪酸組成的影響[J].海洋科學(xué),2000, 24(7): 46-50. [4] Evans C T, Ratledge C.Influence of Nitrogen Metabolism on Lipid Accumulation by Rhodosporidium to ruloides CBS14[J].Gen Microbiol, 1984, 130(7): 1704-1705. [5] Yoon S H, Rhee J A.Quantitative Physiology of Rhodotorula glutinis for Microbial lipid Production[J].Process Biochem, 1983, 18(5): 2-4. [6] 李雅娟, 王起華.氮、磷、鐵、硅營(yíng)養(yǎng)鹽對(duì)底棲硅藻生長(zhǎng)速率的影響[J].大連水產(chǎn)學(xué)院學(xué)報(bào), 1998, 13(4):7-14. [7] 郭峰, 朱凌俊, 柯才煥, 等.兩種海洋底棲硅藻的培養(yǎng)條件研究[J].廈門大學(xué)報(bào)(自然科學(xué)版), 2005, 44(6):831-835. [8] Wang Qa, Li M , Wang S, et al.Studies on culture conditions of bent hic diatoms for feeding abalone: ⅡEffects of salinity, pH, nitrogenous and phosphate nutrients on growth rate[J].Chin J Oceanol Limnol ,1998, 16 (1) : 78-83. [9] 董金利.海洋底棲硅藻Nitzschia constrict的分離、培養(yǎng)及固定化研究[D].福州: 福建師范大學(xué),2010. [10] 程軍, 葛婷婷, 馮佳, 等.硅藻生長(zhǎng)優(yōu)化富集油脂的促進(jìn)機(jī)理[J].新能源及工藝, 2010, 4: 27-32. [11] Chelf P.Environmental control of lipid and biomass production in two diatom species[J].Journal of Applied Phycology, 1990, 2: 121-130. [12] Li Y, Han D, Sommerfeld M, et al.Photosynthetic carbon partitioning and lipid production in the oleaginous microalgaPseudochlorococcumsp.(Chlorophyceae)under nitrogen-limited conditions[J].Bioresource Technology,2011, 102 (1): 123-129. [13] Coverti A, Casazza A A, Ortiz E Y, et al.Effect of temperature and nitrogen concentration on the growth and lipid content ofNannochloropsis oculataandChlorella vulgarisfor biodiesel production[J].Chemical Engineering and Processing: Process Intensification,2009, 486: 1146-1151. [14] 王軍, 楊素玲, 叢威, 等.營(yíng)養(yǎng)條件對(duì)產(chǎn)烴葡萄藻生長(zhǎng)的影響[J].過(guò)程工程學(xué)報(bào), 2003, 32: 141-145. [15] Reitan K I, Rainuzzo J R, Olsen Y.Effect of nutrient limitation on fatty acid and lipid content of marine microalgae[J].Phycol, 1994, 30(6): 972-979. [16] 馬志珍, 季梅芳, 陳匯遠(yuǎn).一種可作為鮑和海參餌料的底棲舟形藻的培養(yǎng)條件的研究[J].海洋通報(bào), 1985,4(4): 36-39. [17] 王淵源, 姜慶國(guó), 江航宇.培養(yǎng)小形舟形藻的氮、磷肥料量[J].1986, 10(5): 35-37. [18] Mortensen S H, Borsheim K Y, Rainuzzo J, et al.Fatty acid and elemental composition of the marine diatomChaetoceros gracilisSchütt.Effects of silicatedeprivation, temperature and light intensity.[J].Journal of Experimental Marine Biology and Ecology, 1988,122(2): 173-185. [19] 李文權(quán), 蔡阿根, 王憲, 等.光和營(yíng)養(yǎng)鹽對(duì)三角褐指藻生化組成的影響[J].中國(guó)環(huán)境科學(xué), 1994, 14(3):185-189. [20] 張立言, 向保卿, 馬志珍.兩種硅藻生長(zhǎng)繁殖條件的研究[J].海洋水產(chǎn)研究, 1982, 4: 79-87.
猜你喜歡
食品工業(yè)(2023年12期)2023-12-12 12:56:16
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
瘋狂英語(yǔ)·新讀寫(xiě)(2020年3期)2020-06-06 09:06:14
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:08
中國(guó)糧油學(xué)報(bào)(2019年4期)2019-07-12 09:06:38
食品與機(jī)械(2018年11期)2019-01-03 06:15:34
中國(guó)糧油學(xué)報(bào)(2018年12期)2018-01-26 13:32:04
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
