日本血吸蟲凋亡誘導因子SjAIF功能區片段的克隆及表達分析
2015-03-17 02:23:59韓宏曉馬茜茜劉艷濤傅志強林矯矯
中國人獸共患病學報 2015年2期
陸 看,韓宏曉,洪 煬,馬茜茜,劉艷濤,韓 倩,馬 帥,傅志強,林矯矯,3
日本血吸蟲凋亡誘導因子SjAIF功能區片段的克隆及表達分析
陸 看1,2,韓宏曉2,洪 煬2,馬茜茜2,劉艷濤2,韓 倩2,馬 帥2,傅志強2,林矯矯2,3
目的 克隆表達日本血吸蟲凋亡誘導因子(SjAIF)功能區片段,并對其生物學特性進行初步分析。方法 應用PCR技術擴增日本血吸蟲凋亡誘導因子的功能區片段,構建原核重組表達質粒并誘導其表達,實時定量PCR分析其在不同發育時期童蟲和成蟲的轉錄水平,通過Western-blot和間接ELISA法分析重組蛋白的抗原性和免疫原性,應用免疫組化分析該蛋白在蟲體內的分布情況。結果 克隆了SjAIF功能區片段,大小為831 bp。成功構建了原核重組表達質粒pET-28a(+)-SjAIF,并在大腸桿菌中獲得表達,重組蛋白分子量35 kD。實時定量PCR分析表明SjAIF基因在童蟲和成蟲的各個發育階段均有轉錄,其中7 d~21 d蟲體表達量較低,42 d和28 d蟲體表達量較高,雌蟲表達量高于雄蟲。重組蛋白具有較好的抗原性和免疫原性,免疫小鼠后誘導產生了較高水平的特異性IgG抗體。該蛋白主要存在于日本血吸蟲體被,少部分分布于實質組織中。結論 成功表達了SjAIF基因的功能區片段,對其生物學特性進行了初步分析,為進一步研究該基因生物學功能和作用提供了基礎。
日本血吸蟲;細胞凋亡;凋亡誘導因子
血吸蟲病(schistosomiasis)是由血吸蟲尾蚴感染人或其它哺乳動物而引起的一種危害嚴重的人獸共患寄生蟲病,廣泛流行于亞洲、非洲和拉丁美洲。該病流行于全球76個國家和地區,約6億人受到威脅,2億人感染[1]。在我國流行的是日本血吸蟲病。自1977年吡喹酮成功合成至今,該藥一直是唯一一種大規模使用的治療血吸蟲病藥物。而吡喹酮耐藥蟲株的出現使治療血吸蟲新藥物研發更加緊迫[2-3]。日本血吸蟲在不同宿主體內的生長發育情況與蟲體體細胞凋亡相關的發現,為研制血吸蟲病新藥物以及防治提供了新思路[4-6]。
細胞凋亡是由多基因嚴格控制的細胞程序性死亡,對于維持生物體正常的生長發育以及內環境的穩定具有重要作用。哺乳動物發生細胞凋亡主要有三條通路,即死亡受體通路、線粒體通路和內質網通路。有研究表明,血吸蟲與其他多細胞生物一樣具有凋亡相關的編碼基因,但是血吸蟲凋亡的確切機制尚不明確[6]。凋亡誘導因子(apoptosis-inducing factor,AIF)是線粒體通路中的凋亡因子之一,是一種位于線粒體膜間隙的黃素蛋白,具有氧化還原和促凋亡雙重功能。
本實驗室利用RT-PCR技術獲得SjAIF的全長cDNA序列,對SjAIF的功能區片段進行了克隆、表達以及生物信息學分析,制備了針對該基因片段重組蛋白的多克隆抗體,分析了該基因在不同時期童蟲和成蟲及雌雄蟲間的表達差異,對該基因編碼蛋白在蟲體內的分布情況進行了免疫組織定位,為進一步研究其生物學功能和作用提供基礎資料。
1 材料與方法
1.1 材料
1.1.1 主要試劑和酶 TrizolRNA提取試劑盒、逆轉錄酶、RNA酶抑制劑購自Invitrogen;Ex Taq DNA聚合酶、pMD-19T載體、T4DNA連接酶、限制性內切酶SacⅠ和XhoⅠ、熒光實時定量PCR(SYBR Green)檢測試劑盒購自TaKaRa生物工程(大連)有限公司;瓊脂糖凝膠電泳DNA純化回收試劑盒和質粒快速提取試劑盒購自Axygen公司;硝酸纖維素膜(Whatman)購自北京經科宏達生物技術有限公司;Ni-NTA HisBind Resin購自中科新生命生物科技有限公司;DAB底物顯色液、DAPI溶液、四甲基聯苯胺(TMB)購自天根生化科技(北京)有限公司;山羊抗小鼠IgG-HRP、山羊抗兔IgG-HRP、Cy3標記的山羊抗小鼠IgG(H+L)二抗購自康為世紀生物科技有限公司。
1.1.2 菌種、實驗動物和血清 感受態大腸桿菌DH5α、BL21(DE3)購自北京全式金生物科技有限公司;6 w齡雄性BALB/c小鼠購自斯萊克實驗動物中心;雄性新西蘭大白兔購自上海羅涇飛達實驗動物養殖場;表達載體pET-28a(+)和感染42 d兔陽性血清由本實驗室保存。
1.2 方法
1.2.1 蟲體的收集 新西蘭大白兔以腹部貼片法分別感染2 000~20 000條日本血吸蟲尾蚴,于感染后7、14、21、28、35、42 d分別剖殺,以肝門靜脈灌注法收集蟲體,用PBS充分洗滌蟲體。將部分42 d合抱蟲體吹打分開,分別收集雌蟲和雄蟲。將各時期蟲體以及雌雄蟲保存于液氮中備用。
1.2.2 RNA的提取 取保存于液氮中的各個時期蟲體以及雌雄蟲,按照TrizolRNA提取試劑盒說明書分別提取蟲體總RNA。
1.2.3 生物信息學分析和目的基因的擴增 利用DNAStar軟件分析AIF的理論等電點,蛋白質相對分子質量等參數。利用在線軟件Clustal Omega(http://www.ebi.ac.uk/Tools/msa/clustalo/)對AIF蛋白進行多重比對,利用SignalP(www. cbs.dtu/services/Singnalp)在線軟件進行信號肽預測,利用在線軟件SMART(http://smart.embl-heidelberg.de/)和NCBI(http://www.ncbi.nlm.nih.gov/)對SjAIF進行結構預測和功能分析,并由此確定本實驗的功能區片段。根據目的基因的功能區片段設計引物,上游引物P1:5′-GCCGAGCTC GTACCTGAGTCACTTGTG-3′(下劃線處為SacⅠ酶切位點),下游引物P2: 5′-GCCCTCGAG AGCTAGACATGGTGGAAGCAC-3′(下劃線處為XhoⅠ酶切位點),由上海華津生物科技有限公司合成。以反轉錄的日本血吸蟲14 d蟲體cDNA為模板進行PCR擴增。擴增條件為94 ℃預變性5 min; 94 ℃ 30 s,62 ℃ 30 s,72 ℃ 1 min,共30個循環;延伸溫度設為72 ℃ 10 min。PCR產物按照DNA純化試劑盒的步驟純化后,將其連接pMD19-T載體,并將陽性質粒送往上海華津生物科技有限公司測序。
1.2.4 重組表達質粒的構建 將陽性質粒雙酶切后連接到表達載體pET28a(+),構建重組表達質粒pET28a(+)-SjAIF片段,并轉化至感受態BL21中。通過菌液PCR、雙酶切以及測序鑒定陽性重組質粒。
1.2.5 重組質粒在大腸桿菌中的表達以及純化 將鑒定好的pET28a(+)-SjAIF/BL21接種于LB培養基中,以37 ℃,250 r/min培養約2~4 h至OD600≈0.6時,加入IPTG,使其終濃度為1 mmol/L,進行誘導表達,并確定其最佳表達條件和表達形式。將以包涵體形式存在的重組蛋白以Ni-NTA HisBind Resin純化。
1.2.6 熒光實時定量PCR 以日本血吸蟲的看家基因α-tublin為內參。SjAIF實時定量PCR的擴增引物sense:5′-ATATGGCGGTGTGGCGTTTATG-3′,anti-sense:5′-CACTCCTGTTTGCCTACAAGTTTC-3′,其擴增片段長度為172 bp;Sjα-Tublin的實時定量PCR的擴增引物sense:5′-CTGATTTTCCATTCGTTTG-3′,anti-sense:5′-GTTGTCTACCATGAAGGCA-3′,其擴增片段長度為213 bp。引物由上海華津生物科技有限公司合成。Real-time PCR反應體系:2×SYBR Premix Ex Taq 10 μL,上游引物和下游引物各0.4 μL,ROX Reference Dye Ⅱ0.4 μL ,EASY Dilution 6.8 μL,cDNA模板2 μL。反應條件:95 ℃預變性30 s,95 ℃5 s,60 ℃34 s,72 ℃15 s共40個循環,每個反應3孔重復。分析得出相對于看家基因的目的基因含量。
1.2.7 Western blotting檢測 將純化的重組蛋白進行SDS-PAGE電泳,經電轉移至硝酸纖維素膜上。用重組蛋白SjAIF免疫的BALB/c小鼠血清和感染日本血吸蟲42 d的兔陽性血清為一抗孵育,以未感染血吸蟲的小鼠和兔血清作為對照,再分別以辣根過氧化物酶標記的山羊抗小鼠IgG和山羊抗兔IgG作為二抗進行孵育。用DAB作為底物避光顯色。
1.2.8 ELISA檢測免疫小鼠血清抗重組蛋白特異性IgG抗體水平 將30只雄性BALB/c小鼠分為蛋白免疫組、206佐劑組和PBS組,每組10只。免疫程序為:每2周免疫1次,共免疫3次,在1免前和每次免疫1周后眼眶靜脈采血,第3次免疫后2周,每只小鼠經腹部貼片接種日本血吸蟲尾蚴,并于感染后6 w后剖殺,收集剖殺后血清備用。ELISA檢測各組血清中的特異性IgG抗體水平。
1.2.9 免疫組化分析 取21 d血吸蟲蟲體制成6 μm的冰凍切片。切片用丙酮固定30 min,10%的山羊血清封閉2 h,以重組蛋白免疫BALB/c小鼠的3免血清為一抗室溫孵育2 h,同時以未感染小鼠血清作為陰性血清對照。用Cy3標記的羊抗鼠IgG為二抗室溫避光孵育,最后用DAPI溶液復染10 min。在各步驟之間用PBST洗滌3次,每次5 min。
2 結 果
2.1SjAIF基因的生物信息學分析 生物信息學分析表明SjAIFcDNA全長為3 441 bp,編碼的蛋白不含信號肽,理論等電點為6.61。本實驗綜合SjAIF的功能區結構域及抗原性分析,選擇編碼該蛋白的第200-476位氨基酸的核苷酸片段進行克隆、表達和初步分析。選擇來自埃及血吸蟲、華支睪吸蟲、細粒棘球絳蟲、多房棘球絳蟲、原雞、人(Accession No.分別為KGB36175.1、GAA47621.1、CDS21975.1、CDI96960.1、NP_001007491.1、O95831) 6個物種的AIF蛋白進行氨基酸序列的多重比對。結果顯示,該基因所編碼的氨基酸序列與埃及血吸蟲的AIF相似性最高,達到68%,與華支睪吸蟲、細粒棘球絳蟲、多房棘球絳蟲、原雞和人的相似性分別為45%,41%,43%,35%和33%(圖 1)。
2.2 重組質粒在大腸桿菌中的表達及純化 通過PCR、酶切鑒定和測序分析,表明成功構建了重組表達質粒pET28a(+)-SjAIF(圖2、圖3)。
重組表達質粒在大腸桿菌BL21中獲得表達,且在誘導5h時表達量最高(圖4)。SDS-PAGE結果顯示重組蛋白分子量約為35 kDa,與預期結果相符。重組蛋白以包涵體形式存在。經Ni-NTA樹脂純化后,獲得較純的重組蛋白。
2.3SjAIF基因在不同時期童蟲和成蟲以及雌雄蟲表達差異分析 RT-PCR結果顯示,SjAIF在所檢測的各個時期均有表達,在14 d蟲體表達量最低,42 d和28 d蟲體表達量較高,雌蟲表達量高于雄蟲(圖5)。
2.4 Western-blotting 檢測重組蛋白的免疫原性 以重組蛋白免疫鼠血清和感染血吸蟲42 d的兔陽性血清為一抗孵育,結果顯示在35 kDa處有明顯的識別條帶,而以健康小鼠或兔血清作為對照則沒有陽性條帶出現,說明重組蛋白具有很好的免疫原性和抗原性(圖6)。
2.5 小鼠血清抗SjAIF特異性IgG抗體水平的檢測 結果表明,免疫組小鼠在一免后就產生了特異性IgG抗體,在二免、三免至剖殺后特異性IgG水平持續升高。而206佐劑對照組和PBS對照組小鼠血清的特異性IgG抗體水平基本保持不變,一直維持在較低水平,只在血吸蟲攻擊感染后6周才見到略有升高(圖7)。

圖1 不同物種SjAIF基因氨基酸序列的同源性分析
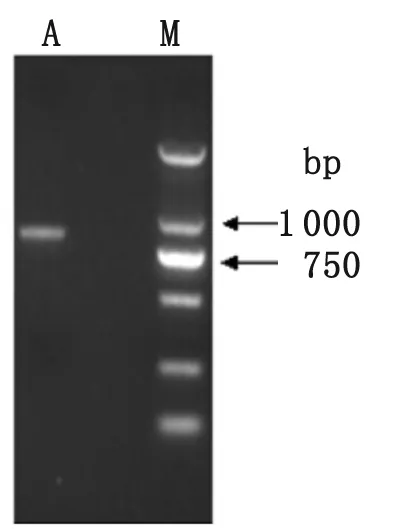
M: DNA marker; A: Product of PCR
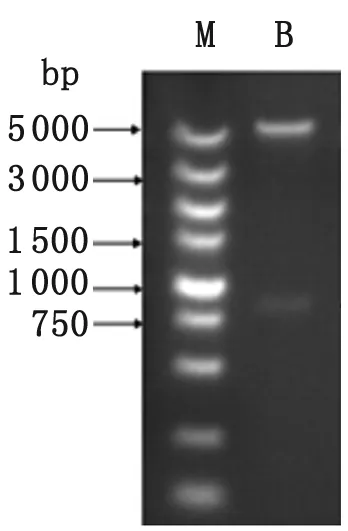
M: DNA marker; B: pET-28a-SjAIF digested withSacI andXhoI.
圖3 重組質粒pET-28a-SjAIF的雙酶切鑒
Fig.3 Recombinant plasmid identified by enzyme digestion
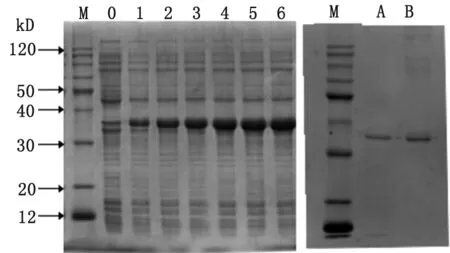
M: Marker; 0-6: Recombinant plasmid pET-28a(+)-SjAIF induced with IPTG for 0-6h; A, B: Purified recombinant protein ofSjAIF.
圖4 SDS-PAGE分析pET-28a(+)-SjAIF/BL21的蛋白表達情況
Fig.4 SDS-PAGE analysis of the expression of pET-28a(+)-SjAIF/BL21 inE.coli
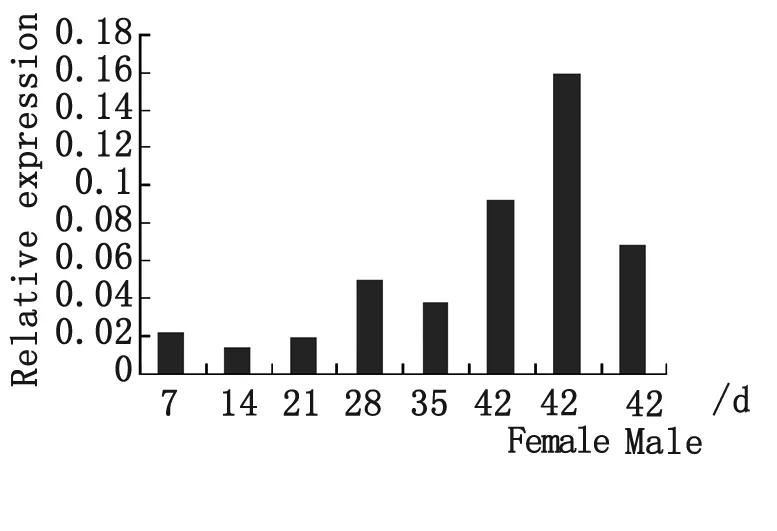
圖5 熒光實時定量PCR分析SjAIF基因在日本血吸蟲不同階段童蟲和成蟲的表達分析
Fig.5 Expression profiles ofSjAIF at differential stages of schistosomula and adult worms ofS.japonicumby real-time PCR
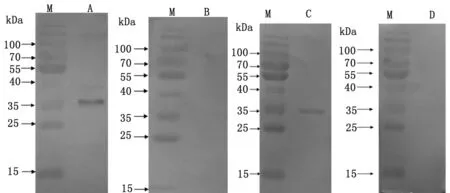
M: marker; A:Recombinant protein was probed with mouse sera againstSjAIF; C:Recombinant protein was probed with sera from rabbit infected withS.japonicum; B,D: Recombinant protein was probed with the normal mice or rabbit sera respectively.
圖6 pET28a(+)-SjAIF重組蛋白的Western blotting分析
Fig.6 Western blotting analysis ofSjAIF expression product
2.5SjAIF蛋白在蟲體內的分布分析:在熒光顯微鏡下可以觀察到用Cy3標記的羊抗鼠IgG二抗發出紅色熒光以及用DAPI復染核酸后發出的藍色熒光,結果表明SjAIF蛋白主要存在于日本血吸蟲體被,少部分分布于實質組織中(圖8)。
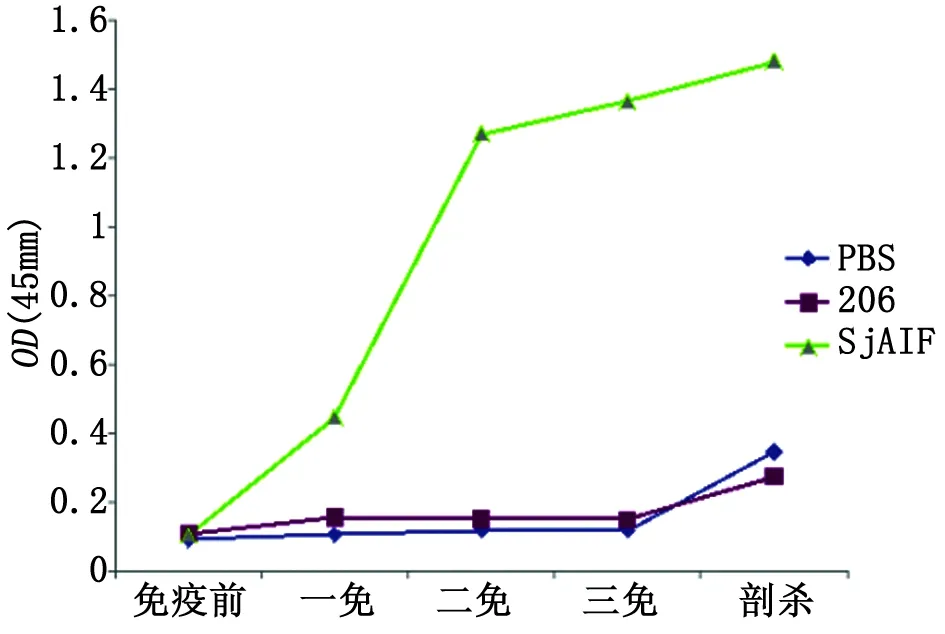
圖7 BALB/c小鼠血清抗SjAIF抗原特異性IgG抗體水平檢測
Fig.7 Specific IgG level againstSjAIF by ELISA
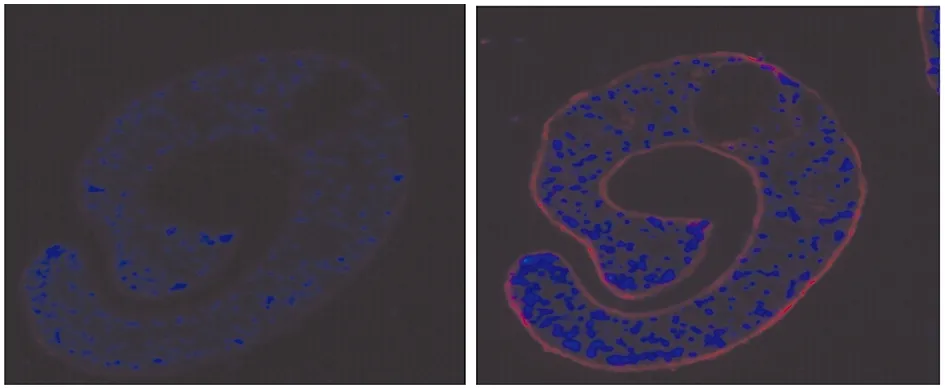
圖8SjAIF蛋白在21d日本血吸蟲蟲體內的分布分析(10×40)
Fig.8 Localization analysis ofSjAIF by immunolocalization in 21d-day-old worms ofS.japonicum(10×40)
3 討 論
細胞凋亡是多種生物平衡細胞增殖與組織發育的重要機制。凋亡過程的紊亂可能直接或間接導致許多疾病的發生,如癌癥和自身免疫性疾病等。根據不同哺乳動物對日本血吸蟲感染的易感性可把日本血吸蟲終末宿主分為適宜宿主、非適宜宿主和抗性宿主。東方田鼠是迄今為止發現的唯一一種日本血吸蟲無法在其體內發育成熟的哺乳類動物抗性宿主[7]。已有研究表明,一些不同宿主來源的日本血吸蟲凋亡相關基因呈現差異表達,日本血吸蟲可能存在類似哺乳動物或線蟲等的凋亡通路[6,8]。有實驗發現,腫瘤治療藥物伊馬替尼能誘導曼氏血吸蟲蟲體生殖器官細胞發生凋亡,最終致使蟲體死亡[9]。細胞凋亡研究有可能為血吸蟲新藥靶和候選疫苗分子的篩選提供新思路。
凋亡誘導因子(AIF)是一種位于線粒體內外膜間隙的黃素蛋白,具有氧化還原酶和促凋亡雙重功能。正常情況下AIF能夠依賴其氧化還原酶的功能催化細胞色素C和NAD之間的電子傳遞而阻止凋亡。當線粒體受到凋亡刺激后,引起線粒體膜通透性轉運孔的結構發生變化而開放,導致凋亡誘導因子AIF從線粒體釋放到細胞質中,最終引起細胞凋亡[10]。AIF是線粒體凋亡通路中的凋亡因子之一,是第一個被鑒定出可以不依賴半胱天冬酶(caspase)信號通路而直接介導細胞發生凋亡的分子[11]。但后來又有研究發現在線蟲體內,CED-3(caspase的同源物)的活性對于WAH-1(AIF的同源類似物)從線粒體釋放很重要,表明在線蟲體內WAH-1的促凋亡活性依賴于CED-3[12]。另有研究發現AIF引起的細胞凋亡與caspase級聯反應之間并不是完全獨立的,激活的caspase能使純化的線粒體釋放AIF因子,Bcl-2家族在某種程度上能抑制AIF從線粒體的釋放[13-14]。因此猜測在細胞死亡的級聯反應中,AIF和caspase可能存在一種合作關系,而它們相互之間的作用可能取決于凋亡刺激物和細胞的類型。而AIF在日本血吸蟲中的作用及其機制尚不明確。
生物信息學分析表明本實驗研究的SjAIF功能區片段包含一個吡啶核苷酸-二硫化物氧化還原酶家族蛋白,是一種腫瘤相關蛋白。有研究發現FAD依賴的吡啶核苷酸-二硫化物氧化還原酶DepH可以催化抗癌藥物FK228前體形成二硫鍵,促進FK228的活化,進而促進癌細胞凋亡和癌癥的康復[15]。由此猜測SjAIF的吡啶核苷酸-二硫化物氧化還原酶結構域的功能行使是否與FAD/NAD(P)結合域結構域(第221-428位氨基酸)的功能有關?但血吸蟲在此方面的研究尚不多見,本實驗為發現促進血吸蟲凋亡的潛在新靶點奠定了基礎。本實驗研究結果表明SjAIF基因在血吸蟲感染宿主7 d、14 d、21 d、28 d、35 d、42 d都有表達,在7 d~21 d表達量較低,可能與維持線粒體的結構穩定,以提供大量的能量用于血吸蟲的形態結構和生理功能的改變,使血吸蟲在終末宿主體內正常的生長發育有關。Western blotting結果表明該重組表達產物具有良好的抗原性,用SjAIF重組抗原免疫BALB/c小鼠后,可誘導小鼠體內的抗重組抗原的特異性IgG抗體迅速產生,并且維持在一個較高的水平。免疫組化結果表明該蛋白主要存在于日本血吸蟲體被與實質組織中。
本實驗首次克隆表達了SjAIF基因的功能區片段,并對其生物學特性進行了初步研究,為進一步探討SjAIF在血吸蟲生長發育中的作用以及其功能奠定了基礎。
[1]Zhang SY, Jiang SF. Development plan of the schistosomiasis vaccines[J]. Foreign Med Sci Parasit Dis, 2004, 31(2): 66-68. (in Chinese) 張述義, 蔣守富. 血吸蟲病疫苗開發行動計劃[J].國外醫學: 寄生蟲病分冊, 2004, 31(2): 66-68.
[2]Liang YS, Li HJ, Dai JR, et al. Studies on resistance ofSchistosomatopraziquantel XIII resistance ofSchistosomajaponicumto praziquantel is experimentally induced in laboratory[J]. Chin J Schisto Ctrl, 2011, 23(6): 605-610. (in Chinese) 梁幼生, 李洪軍, 戴建榮, 等. 血吸蟲對吡喹酮抗藥性的研究XIII日本血吸蟲吡喹酮抗藥性的實驗誘導[J]. 中國血吸蟲病防治雜志, 2011, 23(6): 605-610.
[3]Cioli D. Chemotherapy of schistosomiasis: an update[J]. Parasitol Today, 1998, 14(10): 418-422. DOI: 10.1016/S0169-4758(98)01323-4
[4]Peng JB, Hong Y, Gobert GN, et al. Apoptosis governs the elimination ofSchistosomajaponicumfrom the non-permissive hostMicrotusfortis[J]. PLoS One, 2011, 6(6): e21109. DOI: 10.1371/journal.pone.0021109
[5]Guo XY, Hong Y, Han HX, et al. Observation of apoptosis in 14 days-old Schistosomulum ofSchistosomajaponicum[J]. Chin Vet Sci, 2014, 44(06): 558-562. (in Chinese) 郭小勇, 洪煬, 韓宏曉, 等. 日本血吸蟲14d童蟲凋亡現象的觀察[J]. 中國獸醫科學, 2014, 44(06): 558-562.
[6]Han HX, Peng JB, Gobert GN, et al. Apoptosis phenomenon in the schistosomulum and adult worm life cycle stages ofSchistosomajaponicum[J]. Parasitol Int, 2013, 62(2): 100-108. DOI: 10.1016/j.parint.2012.09.008
[7]Li H, He YY, Lin JJ, et al. The observation for the phenomenon ofMicrotusfortisaganistingSchistosomajaponicum[J]. Chin J Vet Parasitol, 2000, 8(2): 12-15. (in Chinese) 李浩, 何艷燕, 林矯矯, 等. 東方田鼠抗日本血吸蟲病現象的觀察[J]. 中國獸醫寄生蟲病雜志, 2000, 8(2): 12-15.
[8]Peng JB, Han HX, Gobert GN, et al. Differential gene expression inSchistosomajaponicumSchistosomula from Wistar rats and BALB/c mice[J]. Parasite Vector, 2011, 4(1): 155-166. DOI: 10.1186/1756-3305-4-155
[9]Beckmann S, Grevelding CG. Imatinib has a fatal impact on morphology, pairing stability and survival of adultSchistosomamansoniinvitro[J]. Int J Parasitol, 2010, 40(5): 521-526. DOI: 10.1016/j.ijpara.2010.01.007
[10]Wang CZ, Cao C, Ma QJ. The regulation of apoptosis by apoptosis-inducing factor[J]. Chem Life, 2005, 25(6): 454-456. (in Chinese) 王昌正, 曹誠, 馬清鈞. 凋亡誘導因子(AIF)對細胞凋亡的調控[J]. 生命的化學, 2005, 25(6): 454-456.
[11]Susin SK, Lorenzo HK, Zamzaml N, et al. Molecular characterization of mitochondrial apoptosis-inducing factor[J]. Nature, 1999, 397 (6718): 441-446. DOI: 10.1038/17135
[12]Wang XC, Yang CL, Chai JJ, et al. Mechanisms of AIF-mediated apoptotic DNA degradation in caenorhabditis elegans[J]. Science, 2002, 298(5598): 1587-1592. DOI: 10.1126/science.1076194
[13]Cai JY, Yang J, Jones DP. Mitochondrial control of apoptosis: the role of cytochrome c[J]. Biochimica et Biophysica Acta, 1998, 1366: 139-149. DOI: 10.1016/S0005-2728(98)00109-1
[14]Cande C, Cecconi F, Dessen P, et al. Apoptosis-inducing factor (AIF): key to the conserved caspase-independent pathways of cell death?[J]. J Cell Sci, 2002, 115(24): 4727-4734. DOI: 10.1242/jcs.00210
[15]Wang C, Wesener SR, Zhang HL, et al. An FAD-dependent pyridine nucleotide-disulfide oxidoreductase is involved in disulfide bond formation in FK228 anticancer depsipeptide[J]. Chem Biol, 2009, 16(6): 585-593. DOI: 10.1016/j.chembiol.2009.05.005
歡迎訂閱
中國人獸共患病學報
郵發代號:34-46 全年訂價:96元
Cloning and expression analysis of a functional fragment of apoptosis-inducing factor (AIF) gene inSchistosomajaponicum
LU Kan1,2,HAN Hong-xiao2,HONG Yang2,MA Qian-qian2,LIU Yan-tao2,HAN Qian2,MA Shuai2, FU Zhi-qiang2,LIN Jiao-jiao2,3
(1.CollegeofLifeandEnvironmentalSciences,ShanghaiNormalUniversity,Shanghai200234,China;2.ShanghaiVeterinaryResearchInstitute,ChineseAcademyofAgriculturalScience/KeyLaboratoryofAnimalParasitology,MinistryofAgriculture,Shanghai200241,China;3.JiangsuCo-innovationCenterforPreventionandControlofImportantAnimalInfectiousDiseasesandZoonoses,Yangzhou225009,China)
We cloned and expressed a functional fragment of apoptosis-inducing factor (AIF) gene inSchistosomajaponicumand further analyzed its biological characteristics. PCR technique was employed to amplify the functional fragment ofSjAIF by employing a cDNA of 14 d (day) schistosomula as template. The fragment of AIF was subcloned into a pET28a(+) vector and the recombinant plasmid was transformed into competentE.coil/BL21 for producing recombinant protein. The expression level ofSjAIF was determined at several different development stages of schistosomula and adult worms by using real-time RT-PCR. The recombinant protein was purified and then its antigenicity was accessed by Western blotting and ELISA. The distribution of the protein inSchistosomajaponicumwas analyzed by immunolocalization. Real-time PCR analysis revealed that the expression ofSjAIF was lower in 7 d, 14 d and 21 d than that in other stages. Western-blotting showed that the recombinant had good immunogenicity. The vaccinated group showed a good ability to induce IgG as examined by ELISA. Immunolocalization analysis revealed that theSjAIF was mainly distributed in tegument and parenchyma. The fragment ofSjAIF was obtained and its molecular characterizations were preliminarily investigated. This study provided an important basis for further investigation of the biological characteristics and mechanism of the protein inSchistosomajaponicum. Keywords:Schistosomajaponicum; apoptosis; apoptosis-inducing factor
Lin Jiao-jiao, Email: jjlin@shvri.ac.cn
國家自然科學基金(No.81271871)資助
林矯矯,Email:jjlin@shvri.ac.cn
1.上海師范大學生命與環境科學學院,上海 200234; 2.中國農業科學院上海獸醫研究所,農業部動物寄生蟲學重點開放實驗室,上海 200241; 3.江蘇省動物重要疫病與人獸共患病防控協同創新中心,揚州 225009
10.3969/cjz.j.issn.1002-2694.2015.02.002
R383
A
1002-2694(2015)02-0102-07
2014-10-08;
2014-12-22
Supported by the National Natural Science Foundation of People’s Republic of China (No. 81271871)
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59
