N-苯基-2-萘胺對(duì)擬柱胞藻生長(zhǎng)、抗氧化酶及光合系統(tǒng)Ⅱ的影響
2015-02-28 08:28:35冉小飛楊燕君楊宋琪施軍瓊吳忠興
水生生物學(xué)報(bào) 2015年4期
劉 瑞 白 芳 冉小飛 楊燕君 楊宋琪 施軍瓊 吳忠興
(西南大學(xué)生命科學(xué)學(xué)院, 三峽庫(kù)區(qū)生態(tài)環(huán)境教育部重點(diǎn)實(shí)驗(yàn)室, 重慶市三峽庫(kù)區(qū)植物生態(tài)與資源重點(diǎn)實(shí)驗(yàn)室, 重慶 400715)
N-苯基-2-萘胺對(duì)擬柱胞藻生長(zhǎng)、抗氧化酶及光合系統(tǒng)Ⅱ的影響
劉 瑞 白 芳 冉小飛 楊燕君 楊宋琪 施軍瓊 吳忠興
(西南大學(xué)生命科學(xué)學(xué)院, 三峽庫(kù)區(qū)生態(tài)環(huán)境教育部重點(diǎn)實(shí)驗(yàn)室, 重慶市三峽庫(kù)區(qū)植物生態(tài)與資源重點(diǎn)實(shí)驗(yàn)室, 重慶 400715)
為探究化感物質(zhì)對(duì)入侵水華藍(lán)藻-擬柱胞藻(Cylindrospermopsis raciborskii)的影響, 研究對(duì)不同濃度(0、0.05、0.10、0.50、1.00和2.00 mg/L)N-苯基-2-萘胺處理下擬柱胞藻的生長(zhǎng)、抗氧化酶活性及葉綠素?zé)晒庹T導(dǎo)動(dòng)力學(xué)進(jìn)行了測(cè)定。結(jié)果表明, 藻葉綠素 a含量隨 N-苯基-2-萘胺濃度升高而顯著降低, 濃度大于0.50 mg/L時(shí), 擬柱胞藻葉綠素a含量顯著低于對(duì)照。72h, N-苯基-2-萘胺對(duì)擬柱胞藻的EC50為1.02 mg/L。葉綠素?zé)晒庹T導(dǎo)動(dòng)力學(xué)參數(shù)表明, 濃度小于0.50 mg/L時(shí), 擬柱胞藻的最大光化學(xué)效率(φP0)、單位面積上有活性的反應(yīng)中心(RC/CS0)、以吸收光能為基礎(chǔ)的性能指數(shù)(PIABS)及以單位面積為基礎(chǔ)的性能指數(shù)(PICS)均顯著高于對(duì)照, 而反映2ms時(shí)有活性的反應(yīng)中心的開(kāi)放程度的ψ0則顯著低于對(duì)照組。當(dāng)N-苯基-2-萘胺濃度大于0.50 mg/L時(shí), 以上參數(shù)均呈現(xiàn)相反趨勢(shì)。這表明低濃度N-苯基-2-萘胺能提高擬柱胞藻的光合效率, 而在高濃度時(shí), 擬柱胞藻PSⅡ有活性的反應(yīng)中心減少, 電子傳遞受到阻礙, 光合效率下降。與對(duì)照相比, 在濃度高于0.50 mg/L條件下, 擬柱胞藻的過(guò)氧化氫酶(CAT)、過(guò)氧化物酶(SOD)及谷胱甘肽過(guò)氧化物酶(GPX)的活性顯著增加, 表明擬柱胞藻能產(chǎn)生大量的抗氧化酶來(lái)減少 N-苯基-2-萘胺的損傷。這些結(jié)果表明 N-苯基-2-萘胺可能通過(guò)抑制PSⅡ的電子傳遞和減少其有活性的反應(yīng)中心來(lái)影響擬柱胞藻的生長(zhǎng), 也暗示了N-苯基-2-萘胺能作為潛在的抑藻物質(zhì)來(lái)控制擬柱胞藻水華。
光合作用; 擬柱胞藻; 抗氧化體系; 化感物質(zhì)
近年來(lái), 藍(lán)藻水華在許多富營(yíng)養(yǎng)化湖泊和水庫(kù)中頻繁暴發(fā)[1, 2]。藍(lán)藻水華的暴發(fā), 造成湖泊水質(zhì)惡化、透明度下降、厭氧程度提高、生物多樣性下降和優(yōu)勢(shì)物種改變, 從而破壞了水體生態(tài)系統(tǒng)結(jié)構(gòu)和功能, 對(duì)人類健康、生態(tài)安全造成巨大的破壞[3]。據(jù)統(tǒng)計(jì), 在世界范圍內(nèi)大約有40個(gè)藍(lán)藻屬, 其中包括最常見(jiàn)的微囊藻 Microcystis Kützing、魚(yú)腥藻Anabaena Bory、束絲藻Aphanizomenon Morren等均可以形成水華[4]。然而, 自1979年澳大利亞北昆士蘭帕門(mén)島 148人中毒物質(zhì)-擬柱胞藻毒素(Cylindrospermopsin)被揭示出來(lái)后, 擬柱胞藻(Cylindrospermopsis Seenaya et Subba Raju)的研究才開(kāi)始引起藻類學(xué)者、生態(tài)學(xué)者和管理部門(mén)的關(guān)注[5]。
擬柱胞藻屬于藍(lán)藻門(mén)(Cyanophyta)、念珠藻目(Nostocales)、念珠藻科(Nostocaceae), 以拉氏擬柱胞藻(C. raciborskii)為模式種。由于擬柱胞藻已經(jīng)由熱帶地區(qū)擴(kuò)張到溫帶地區(qū), 甚至北緯 53°—54°, 因此其被認(rèn)為是一種入侵的水華藍(lán)藻[6]。近年來(lái)的研究表明擬柱胞藻不僅已在我國(guó)水體分布, 而且呈現(xiàn)出明顯的增加趨勢(shì), 甚至在一些大型水庫(kù)、湖泊中已成為優(yōu)勢(shì)種類并產(chǎn)生水華[7, 8]。此外, 擬柱胞藻能夠產(chǎn)生肝毒素以及貝類麻痹毒素等多種毒素[9, 10], 對(duì)魚(yú)類、牲畜甚至是人類的健康都有巨大的威脅。因此,預(yù)防和控制擬柱胞藻水華具有十分重要的意義。
藻類水華控制的方法主要包括物理法、化學(xué)法以及生物法。前兩種方法比較費(fèi)時(shí)、費(fèi)錢(qián), 甚至對(duì)環(huán)境造成二次污染, 因此, 均不被推薦廣泛使用[11, 12]。生物源抑藻物質(zhì)在抑藻方面的作用相對(duì)于前兩種方法具有安全和對(duì)水環(huán)境生態(tài)系統(tǒng)危害小等優(yōu)點(diǎn), 已引起各國(guó)學(xué)者的高度重視[13, 14]。1969 年Fitzgerald[15]發(fā)現(xiàn)水生植物的代謝產(chǎn)物可以控制藻類的生長(zhǎng), 水生植物通過(guò)化感作用抑制藻類的生長(zhǎng)繁殖引起人們的廣泛興趣, 并希望通過(guò)對(duì)化感物質(zhì)的研究開(kāi)發(fā)高效專一的除藻劑, 進(jìn)而為湖泊富營(yíng)養(yǎng)化的生態(tài)治理探索切實(shí)可行的方法。
N-苯基-2-萘胺是高等植物鳳眼蓮(Eichhornia crassipes)和小花地不容(Stephania micrantha)等產(chǎn)生的一種次生代謝產(chǎn)物[16, 17]。N-苯基-2-萘胺代謝產(chǎn)物2-萘胺為致癌物質(zhì), 但其在人體內(nèi)代謝為 2-萘胺的轉(zhuǎn)化率低于 1%[18]。美國(guó)政府工業(yè)衛(wèi)生學(xué)家會(huì)議(ACGIH)指出, N-苯基-2-萘胺本身不被定義為人類致癌物質(zhì)[19]。而藻類細(xì)胞對(duì)該化合物的敏感性極高,微劑量就能顯著抑制藻類細(xì)胞的生長(zhǎng)[20, 21]。
本實(shí)驗(yàn)通過(guò)分析不同濃度下 N-苯基-2-萘胺對(duì)擬柱胞藻生長(zhǎng), 抗氧化酶及光合系統(tǒng)Ⅱ熒光動(dòng)力學(xué)參數(shù)的影響, 旨在探究N-苯基-2-萘胺對(duì)擬柱胞藻抑藻機(jī)理, 為擬柱胞藻等藍(lán)藻水華的預(yù)防和治理提供依據(jù)。
1 材料與方法
1.1 藻體培養(yǎng)
擬柱胞藻FACHB-1096由中國(guó)科學(xué)院水生生物研究所淡水藻種庫(kù)提供。藻種擴(kuò)大培養(yǎng)至對(duì)數(shù)期,取對(duì)數(shù)期藻液, 離心后分別接入含不同濃度 N-苯基-2-萘胺(0、0.05、0.10、0.50、1.00和2.00 mg/L)的200 mL MA (pH 8.6)[22]培養(yǎng)基中, 于光照培養(yǎng)箱中培養(yǎng), 每個(gè)濃度設(shè) 3個(gè)重復(fù)。培養(yǎng)條件為: 溫度(25±0.1)℃ , 25 μmol photons /(m2·s)、光暗比為12h︰12h。為避免培養(yǎng)過(guò)程中藻體成團(tuán), 每天定時(shí)搖動(dòng)錐形瓶幾次。
1.2 葉綠素及EC50測(cè)定
取5 mL的藻液, 4000×g離心力下離心10min,去掉上層培養(yǎng)液, 藻體加入 90%的丙酮, 根據(jù)Nusch的方法進(jìn)行提取[23]。采用概率單位法(Probit method), 利用SPSS 16.0 中Probit 程序?qū)M柱胞藻在N-苯基-2-萘胺暴露72h后的EC50值進(jìn)行計(jì)算[24, 25]。
1.3 葉綠素a熒光動(dòng)力學(xué)曲線
取5 mL藻液暗適應(yīng)20min, 用便攜式植物效率分析儀(Handy-PEA,英國(guó)漢莎公司)測(cè)定不同濃度N-苯基-2-萘胺處理后的擬柱胞藻多相葉綠素?zé)晒庾兓y(cè)定時(shí)光化光為3000 μmol quanta/(m2·s), 收集10μs至2s的熒光信號(hào)。測(cè)定的JIP參數(shù), 參照Strasser 和Strasser[29]的方法選取了表1中參數(shù)進(jìn)行分析。
1.4 抗氧化酶SOD、CAT、GPX活性測(cè)定
取30 mL在不同濃度N-苯基-2-萘胺中培養(yǎng)的藻液, 4000×g離心 5min, 棄上清, 下層藻液用蒸餾水洗滌后液氮研磨, 加入4℃預(yù)冷的4 mL PBS (PBS的濃度為0.05 mol/L、pH為7.8)緩沖液提取粗酶液。過(guò)氧化氫酶(CAT)以及超氧化物歧化酶(SOD)活性的測(cè)定分別參照Choo等[26]和Wu等[27]的方法。谷胱甘肽過(guò)氧化物酶 GPX活性采用谷胱甘肽過(guò)氧化物酶(GSH-PX)試劑盒(南京建成生物工程研究所)測(cè)定。可溶性蛋白采用考馬斯亮藍(lán)法測(cè)定[28]。酶活性單位以單位蛋白濃度(U/mg 蛋白)表示。

表1 選取的JIP-測(cè)定參數(shù)及其意義Tab. 1 The selected JIP test parameters and there meaning
1.5 數(shù)據(jù)分析
實(shí)驗(yàn)數(shù)據(jù)處理使用統(tǒng)計(jì)軟件SPSS 16.0, 所有數(shù)據(jù)均采用單因素方差分析(one-way ANOVA)。用LSD (P<0.05)檢驗(yàn)處理組與對(duì)照組之間的差異, 當(dāng)P <0.05時(shí), 差異顯著, P<0.01時(shí), 差異極顯著。
2 結(jié)果
2.1 葉綠素a含量及EC50值
隨著濃度的增加, 高濃度處理組(>0.50 mg/L)葉綠素a含量顯著低于對(duì)照(圖1)。相比之下, 低濃度處理組(<0.50 mg/L)葉綠素 a含量顯著上升(P<0.05)。隨時(shí)間變化, 濃度為0.10 mg/L的處理組,葉綠素a含量與對(duì)照組相比呈現(xiàn)輕微降低(P>0.05)。通過(guò)SPSS中Probit程序計(jì)算, 獲得N-苯基-2-萘胺對(duì)擬柱胞藻的72h EC50值為1.02 mg/L。
2.2 葉綠素?zé)晒鈩?dòng)力學(xué)曲線O-J段
在不同濃度 N-苯基-2-萘胺處理后, 低濃度(<0.50 mg/L)處理組曲線與對(duì)照組無(wú)明顯差異(P>0.05, 圖2)。N-苯基-2-萘胺濃度高于0.50 mg/L 時(shí), 曲線發(fā)生較大變化, J點(diǎn)幾乎消失, 但各濃度在曲線O點(diǎn)至J點(diǎn)間均未出現(xiàn)明顯拐點(diǎn)。

圖1 N-苯基-2-萘胺濃度對(duì)擬柱胞藻葉綠素a含量的影響Fig. 1 Effect of N-phenyl-2-naphthylamine concentration on the Chl. a content of C. raciborskii
2.3 葉綠素?zé)晒鈩?dòng)力學(xué)參數(shù)
結(jié)果表明單位面積上有活性的反應(yīng)中心(RC/ CS0), 最大光化學(xué)效率(φP0), 以吸收光能為基礎(chǔ)的性能指數(shù)(PIABS)及以單位面積為基礎(chǔ)的性能指數(shù)(PICS)表現(xiàn)出相同的趨勢(shì), 即低濃度處理(<0.50 mg/L)顯著高于對(duì)照, 而高濃度時(shí)顯著低于對(duì)照(P<0.01, 圖3)。與對(duì)照組相比, φP0在0.05、0.10、0.50、1.00 和 2.00 mg/L五個(gè)濃度時(shí), 分別是對(duì)照組的 1.08、1.05、0.34、0.22和 0.13倍, 而 PIABS與 PICS則分別為1.23、1.08、0.02、0.04、0.01和1.24、1.08、0.02、0.05和0.01倍。反映反應(yīng)中心捕獲的激子中,用來(lái)推動(dòng)電子傳遞到電子傳遞鏈中超過(guò) QA的其他
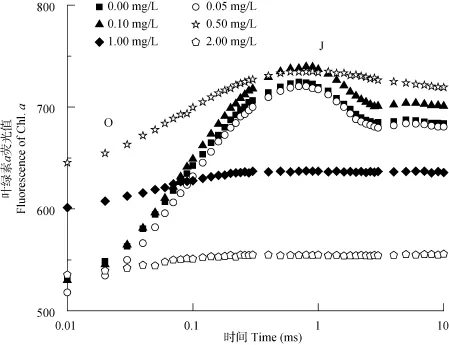
圖2 不同濃度N-苯基-2-萘胺處理后擬柱胞藻快速葉綠素?zé)晒馇€O-J段變化Fig. 2 Fast fluorescence transient curves from O to J of C. raciborskii treated with different concentrations of N-phenyl-2-naphthylamine
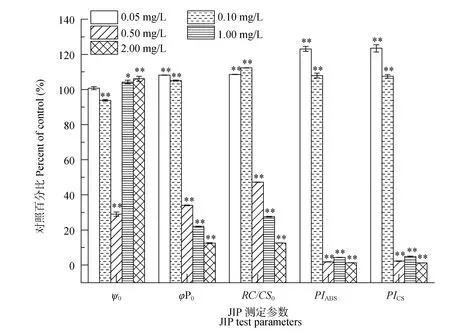
圖3 不同濃度N-苯基-2-萘胺處理后擬柱胞藻JIP 測(cè)定參數(shù)的變化(與對(duì)照相比百分?jǐn)?shù))Fig. 3 Changes in the JIP-test parameters expressed as a percentage of the control when C. raciborskii were exposed for 48h to various concentrations of N-phenyl-2-naphthylamine
電子受體的激子占用來(lái)推動(dòng)QA還原激子的比率, 即照光 2ms 時(shí)有活性的反應(yīng)中心的開(kāi)放程度的參數(shù) ψ0則表現(xiàn)出相反的趨勢(shì), 即低濃度處理時(shí)(0.10和0.50 mg/L), ψ0顯著低于對(duì)照, 分別降低了 6.19%、71.01%, 而高濃度(1.00和2.00 mg/L)處理組則顯著高于對(duì)照, 分別增加了4.26%和6.12% (P<0.01)。
2.4 抗氧化酶SOD、CAT及GPX活性
與對(duì)照相比, SOD和GPX活性隨N-苯基-2-萘胺濃度的升高顯著的上升(P<0.01), 而 CAT活性只在高濃度的 N-苯基-2-萘胺處理下呈顯著上調(diào)(P<0.01, 圖4)。在2.00 mg/L處理下, CAT、SOD及GPX活性分別為對(duì)照的2.78、1.58、2.57倍。
3 討論
先前的研究已經(jīng)表明, N-苯基-2-萘胺一種對(duì)藻類作用極強(qiáng)的化感物質(zhì)[21, 30—32]。本研究發(fā)現(xiàn)隨 N-苯基-2-萘胺濃度的升高, 葉綠素a含量明顯下降(圖1), 這與Qian等[21, 32]研究結(jié)果一致, 表明N-苯基-2-萘胺抑制了擬柱胞藻葉綠素的合成。Bj?rkman[33]研究表明葉綠素濃度降低反映了光合反應(yīng)中心復(fù)合體天線色素減少, 說(shuō)明N-苯基-2萘胺處理后, 擬柱胞藻光合系統(tǒng)可能發(fā)生損傷。在 72h, N-苯基-2萘胺處理下, 擬柱胞藻EC50為1.02 mg/L。相比一些已鑒定出的化感物質(zhì), 如 β-紫羅蘭酮、蘆竹堿、兒茶酸和鞣花酸, 其EC50值分別為(21.23 ± 1.87) mg/L[34]、2.10 mg/L[35]、5.00和5.10 mg/L[36]。因此, N-苯基-2-萘胺具有較強(qiáng)的抑制能力, 可作為一種潛在的擬柱胞藻水華抑制劑。
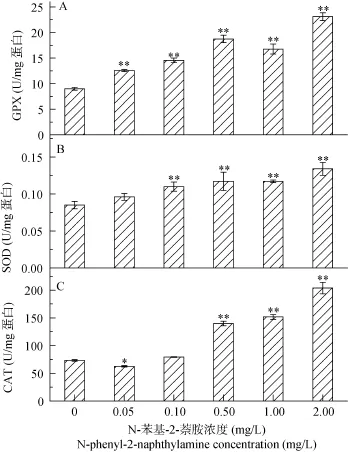
圖4 不同濃度N-苯基-2-萘胺處理后擬柱胞藻GPX(A)、SOD(B)、CAT(C)及活性Fig. 4 GPX(A), SOD (B) and CAT(C) activity in C. raciborskii following exposure to different concentrations of N-phenyl-2-naphthylamine for 48h
研究表明, 當(dāng) PSⅡ的供體側(cè)受到傷害時(shí), 水裂解系統(tǒng)被抑制, 放氧復(fù)合體(OEC)受損, 導(dǎo)致經(jīng)過(guò)極短的時(shí)間(在J點(diǎn)之前), 葉綠素?zé)晒猱a(chǎn)量就會(huì)上升,出現(xiàn)K點(diǎn)(照光后大約 300 μs處的特征位點(diǎn)), 多相熒光O-J 變?yōu)镺-K-J[37, 38]。在本研究中, 未出現(xiàn)K 點(diǎn)(圖 2), 說(shuō)明不同濃度 N-苯基-2-萘胺處理后, 擬柱胞藻PSⅡ供體側(cè)均未受到損傷。研究發(fā)現(xiàn), 在正常條件下, 有活性的PSⅡ反應(yīng)中心將捕獲的光能轉(zhuǎn)化為激發(fā)能, 并將其中的一部分轉(zhuǎn)化為化學(xué)能, 推動(dòng)碳同化反應(yīng), 其余部分耗散掉。然而, 在受到某些脅迫時(shí), 反應(yīng)中心可能失活, 不能將吸收的光能傳遞給電子傳遞鏈。這種失活的反應(yīng)中心可能是作為防止電子傳遞受到光損傷的一種保護(hù)機(jī)制[39]。本研究發(fā)現(xiàn), 單位面積上有活性的反應(yīng)中心在低濃度N-苯基-2-萘胺處理時(shí)(<0.50 mg/L), 顯著高于對(duì)照, 而高濃度時(shí)顯著低于對(duì)照組(圖 3), 表明高濃度的 N-苯基-2-萘胺導(dǎo)致了擬柱胞藻 PSⅡ反應(yīng)中心的失活或降解, 也暗示了擬柱胞藻的受體側(cè)可能受到了抑制, 進(jìn)而導(dǎo)致吸收的光能不能被有效地利用。
φP0表征電子傳遞的能量占有活性的反應(yīng)中心捕獲的能量的比率, 也反映了 PSⅡ最大光化學(xué)效率。在高濃度N-苯基-2-萘胺(>0.50 mg/L)處理下, 擬柱胞藻φP0顯著低于對(duì)照(圖3), 這一結(jié)論支持了前人關(guān)于化感物質(zhì)能抑制藻類電子傳遞鏈能量傳遞[34], 表明高濃度的 N-苯基-2-萘胺降低了擬柱胞藻 PSⅡ用于電子傳遞的能量。與對(duì)照相比, 高濃度N-苯基-2-萘胺處理時(shí), ψo(hù)值顯著增加(圖3), 表明光照2 ms時(shí), PSⅡ反應(yīng)中心的開(kāi)放程度增加, 即用于電子傳遞的能量與用于 QA還原的能量的比例增加[29, 40]。這表明 N-苯基-2-萘胺可能對(duì)擬柱胞藻光合系統(tǒng) II電子傳遞鏈?zhǔn)荏w側(cè)產(chǎn)生了影響。當(dāng)濃度高于 0.50 mg/L 時(shí), 擬柱胞藻光合結(jié)構(gòu)遭到損傷, PSⅡ受體側(cè)電子傳遞受阻, 光合效率下降, 且有活性的反應(yīng)中心減少, 最終導(dǎo)致整個(gè)細(xì)胞光合作用受到強(qiáng)烈的抑制,這與劉彥彥等[41]關(guān)于白屈菜紅堿主要影響銅綠微囊藻光合系統(tǒng)反應(yīng)中心電子供體側(cè)電子傳遞的結(jié)論不同, 說(shuō)明不同化感物質(zhì)對(duì)藻類光合系統(tǒng)的影響有所差異。另外, 在低濃度的N-苯基-2-萘胺(<0.50 mg/L)處理下, 擬柱胞藻 RC/CS0、φP0與對(duì)照相比均有顯著上升, 說(shuō)明低劑量的N-苯基-2-萘胺對(duì)擬柱胞藻生長(zhǎng)有一定的促進(jìn)作用, 支持了前人關(guān)于化感物質(zhì)對(duì)藻類影響的結(jié)果[42—44]。
PIABS及 PICS均在一定程度上反映出脅迫對(duì)光合結(jié)構(gòu)的破壞, PIABS與PICS比φP0更加敏感。已有的研究表明高等植物在受到某些脅迫時(shí), 光合機(jī)構(gòu)遭到破壞, PIABS與 PICS會(huì)顯著下降[45—47]。本研究發(fā)現(xiàn)擬柱胞藻在高濃度 N-苯基-2-萘胺處理下也表現(xiàn)出下降的趨勢(shì)(圖3), 表明擬柱胞藻的光合結(jié)構(gòu)受到了極大的破壞, 光合效率大幅度下降。
在N-苯基-2-萘胺處理下, 擬柱胞藻細(xì)胞內(nèi)自由基(ROS)升高, 隨 N-苯基-2-萘胺的濃度增大, 為維持機(jī)體平衡, 一系列抗氧化酶如清除過(guò)氧化氫的CAT[48], 催化 O2–歧化反應(yīng)的 SOD以及能夠清除過(guò)氧化氫和有機(jī)氫過(guò)氧化物的 GPX[49]活性均升高(圖4), 這一結(jié)論與 Qian等研究結(jié)果是一致的[21], 進(jìn)一步說(shuō)明 N-苯基-2-萘胺處理導(dǎo)致擬柱胞藻產(chǎn)生了氧化損傷。
綜上所述, 在高濃度N-苯基-2-萘胺處理下, 擬柱胞藻的光合結(jié)構(gòu)遭到損傷, PSⅡ電子傳遞受阻,光合效率下降, 且有活性的反應(yīng)中心減少, 抑制整個(gè)細(xì)胞光合作用, 導(dǎo)致了擬柱胞藻生長(zhǎng)受到阻礙。研究已經(jīng)表明 N-苯基-2-萘胺對(duì)小鼠的 LD50超過(guò)了1000 mg/kg[50], 而本研究及其他研究表明微劑量能顯著抑制藻類細(xì)胞的生長(zhǎng)[20, 21], 因此, N-苯基-2-萘胺可以作為一個(gè)潛在的抑藻物質(zhì)來(lái)控制擬柱胞藻水華。然而, 應(yīng)用N-苯基-2-萘胺作為抑藻物質(zhì)仍需大量的野外研究, 特別是N-苯基-2-萘胺劑量及時(shí)間效應(yīng)上仍需大量的工作及數(shù)據(jù)支撐。
[1] O’Neil J M, Davis T W, Burford M A, et al. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change [J]. Harmful Algae, 2012, 14: 313—334
[2] Zhao S, Chen W, Jia Y L, et al. Characteristics of microcystin-degrading bacterium isolated from sedimentations ponds for the treatments of cyanobacterial blooms [J]. Acta Hydrobiologica Sinica, 2013, 37(3): 522—523 [趙爽,陳偉, 賈云璐, 等. 藍(lán)藻水華堆積處理池中微囊藻毒素降解細(xì)菌的分離及降解特性. 水生生物學(xué)報(bào), 2013, 37(3): 522—523]
[3] Carmichael W W. Health Effects of Toxin Producing Cyanobacteria: “The Cyano HABs” [J]. Human and Ecological Risk Assessment, 2001, 7(5): 1393—1407
[4] Paerl H W, Fulton R S, Moisander P H, et al. Harmful freshwater algal blooms, with an emphasis on cyanobacteria [J]. The Scientific World Journal, 2001, 1: 76—113
[5] Byth S. Palm Island mistery disease [J]. The Medical Journal of Australia, 1980, 2(1): 40—42
[6] Briand J F, Leboulanger C, Humbert J F. Cylindrospermopsis raciborskii (Cyanobacteria) invasion at mid-latitudes: selection, wide physiological tolerance, or global warming [J]? Journal of Phycology, 2004, 40(2): 231—238
[7] Wu Z X, Shi J Q, Li R H. Phylogenetic analysis of two cyanobacterial genera Cylindrospermopsis and Raphidiopsis based on multi-gene sequences [J]. Harmful Algae, 2011, 10: 419—425
[8] Lei L M, Peng L, Huang X H, et al. Occurrence and dominance of Cylindrospermopsis raciborskii and dissolved cylindrospermopsin in urban reservoirs used for drinking water supply, South China [J]. Environmental Monitoring and Assessment, 2014, 186(5): 3079—3090
[9] Ohtani I, Moore R E, Runnegar M T C. Cylindrospermopsin: a potent hepatotoxin from the blue-green alga Cylindrospermopsis raciborskii [J]. Journal of the American Chemical Society, 1992, 114(20): 7941—7942
[10] de Souza R C R, Carvalho M C, Truzzi A C. Cylindrospermopsis raciborskii (Wolosz.) Seenaya and Subba Raju (Cyanophyceae) dominance and a contribution to the knowledge of Rio Pequeno arm, Billings Reservoir, Brazil [J]. Environmental Toxicology and Water Quality, 1998, 13(1): 73—81
[11] Yang Q H. Technical advances of eutrophic water restoration in china [J]. Journal of Liaodong University (Natural Science), 2008, 2(15): 26—30 [楊清海. 中國(guó)富營(yíng)養(yǎng)化水體修復(fù)技術(shù)進(jìn)展. 遼東學(xué)院學(xué)報(bào)(自然科學(xué)版), 2008, 2(15): 26—30]
[12] Hu G J, Zhang W H. Research advance of prevention and control of water bloom in lake by biological technology [J]. Safety and Environmental Engineering, 2008, 15(3): 36—40[胡光濟(jì), 張維昊. 湖泊水華生物防治技術(shù)研究進(jìn)展. 安全與環(huán)境工程, 2008, 15(3): 36—40]
[13] Li F M, Hu H Y. Allelopathy and inhibitory effect of extracts from macrophytes on algae growth [J]. China Water& Wastewater, 2004, 20(11): 18—21 [李鋒民, 胡洪營(yíng). 大型水生植物浸出液對(duì)藻類的化感抑制作用. 中國(guó)給水排水, 2004, 20(11): 18—21]
[14] Yang P, Zhang S L, Dong Y. Advances in Inhibitory effects of alkaloids on growth of algae [J]. Fisheries Science, 2012, 31(12): 754—755 [楊攀, 張樹(shù)林, 董陽(yáng). 生物堿類化合物對(duì)藻類生長(zhǎng)抑制作用的研究進(jìn)展. 水產(chǎn)科學(xué), 2012, 31(12): 754—755]
[15] Fitzgerald G P. Some factors in the competition or antagonism among bacteria, algae and aquatic weeds [J]. Journal of Phycology, 1969, 5(4): 351—359
[16] Sultankhodzhaev M N, Tadzhibaev M M. N-phenyl-b-naphthylamine from three species of plants [J]. Chemistry of Natural Compounds, 1976, 12(3): 406—407
[17] Sun W H, Yu S W, Yang S Y, et al. Allelochemicals from root exudates of water hyacinth (Eichhornis crassipes) [J]. Acta Phytophysiologica Sinica, 1993, 19(1): 92—96 [孫文浩,余叔文, 楊善元, 等. 鳳眼蓮根系分泌物中的克藻化合物.植物生理學(xué)報(bào), 1993, 19(1): 92—96]
[18] Weiss T, Brüning T, Bolt H M. Dephenylation of the rubber chemical N-phenyl-2-naphthylamine to carcinogenic 2-Naphthylamine: a classical problem revisited [J]. Critical Reviews in Toxicology, 2007, 37(7): 553—566
[19] ACGIH (American Conference of Governmental Industrial Hygienists), N-phenyl-beta-naphthylamine: TLVs chemical substances 7th edition documentation. In: American Conference of Governmental Industrial Hygienists, Cincinnati, OH. ACGIH Publication 7 DOC—453. 2001
[20] Altenburger R, Brack W, Greco W R, et al. On the mode of action of N-phenyl-2-naphthylamine in plants [J]. Environmental Science and Technology, 2006, 40(19): 6163—6169
[21] Qian H F, Xu X Y, Chen W, et al. Allelochemical stress causes oxidative damage and inhibition of photosynthesis in Chlorella vulgaris [J]. Chemosphere, 2009, 75(3): 368—375 [22] Ichimura T, Isolation and culture methods of algae (S?rui no bunri to baiy? h? . 2.5.B. Tansui s?rui) [A]. In: Nishizawa K, Chihara M (Eds.), Methods in phycological studies (S?rui Kenkyü h?) [C]. Tokyo: Kyoritu Shuppan. 1979, 294—305 [23] Nusch E A. Comparison of different methods for chlorophyll and phaeopigment determination [J]. Archiv für Hydrobiologie-Beiheft Ergebnisse der Limnologie, 1980, 14: 14—36
[24] SPSS Inc. SPSS 10.0 Syntax Reference Guide[M]. Chicago: SPSS Inc. 1999
[25] Zhao J S, Yu S X. Estimation of probit methods on EC50 [J]. Asian Journal of Ecotoxicology, 2010, 5(3): 420—425 [趙勁松, 于書(shū)霞. 利用概率單位法估算 EC50的研究. 生態(tài)毒理學(xué)報(bào), 2010, 5(3): 420—425]
[26] Choo K S, Snoeijs P, Pedersén M. Oxidative stress tolerance in the filamentous green algae Cladophora glomerata and Enteromorpha ahlneriana [J]. Journal of Experimental Marine Biology and Ecology, 2004, 298(1): 111—123
[27] Wu Z, Gan N, Huang Q, et al. Response of Microcystis to copper stress—do phenotypes of Microcystis make a difference in stress tolerance [J]? Environmental Pollution, 2007, 147(2): 324—330
[28] Bradford M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizingthe principle of protein-dye binding [J]. Analytical Biochemistry, 1976, 72(1—2): 248—254
[29] Strasser R J, Srivastava A, Govindjee. Polyphasic chlorophyll a fluorescence transients in plants and cyanobacteria [J]. Photochemistry and Photobiology, 1995, 61(1): 32—42
[30] Dwivedi P C, Rao K. Electron donor—acceptor complexes of N-phenyl-2-naphthylamine with quinones [J]. Indian Journal of Chemistry, 1972, 10: 944—945
[31] Leonova N M, Zhukova E E, Evstigneeva R P. A model of the quinone-binding site of photosystem I: Investigation of charge transfer complexes of phylloquinone and its derivatives with tryptophan [J]. Biofizika, 2004, 49(6): 970—978
[32] Qian H, Yu S, Sun Z, et al. Effects of copper sulfate, hydrogen peroxide and N-phenyl-2-naphthylamine on oxidative stress and the expression of genes involved photosynthesis and microcystin disposition in Microcystis aeruginosa [J]. Aquatic Toxicology, 2010, 99(3): 405—412
[33] Bj?rkman O. Response to different quantum flux densities [A]. In: Lange O L, Nobel P S, Osmond C B, et al (Eds.), Physiological Plant Ecology I [C]. Berlin: Springer-Verlag. 1981, 57—107
[34] Shao J, Xu Y, Wang Z, et al. Elucidating the toxicity targets of β-ionone on photosynthetic system of Microcystis aeruginosa NIES-843 (Cyanobacteria) [J]. Aquatic Toxicology, 2011, 104(1—2): 48—55
[35] Hong Y, Hu H Y, Xie X, et al. Gramine-induced growth inhibition, oxidative damage and antioxidant responses in freshwater cyanobacterium Microcystis aeruginosa [J]. Aquatic Toxicology, 2009, 91(3): 262—269
[36] Nakai S, Inoue Y, Hosomi M, et al. Myriophyllum spicatum-released allelopathic polyphenols inhibiting growth of blue-Green algae Microcystis Aeruginosa [J]. Water Research, 2000, 34(1): 3026—3032
[37] Strivastava A, Strasser R J. Stress and stress management of land plants during a regular day [J]. Journal of Plant Physiology, 1996, 148(3—4): 445—455
[38] Chen H X, Li W J, An S Z, et al. Characterization of PS Ⅱphotochemistry and thermostability in salt-treated Rumex leaves [J]. Journal of Plant Physiology, 2004, 161(3): 257—264
[39] Lee H Y, Hong Y N, Chow W S. Photoinactivation of phtosystem
complexes and photoprotection by non-functional neighbours in Capsicum annuum L. leaves [J]. Planta, 2001, 212(3): 332—342
[40] Li P M, Gao H Y, Strasser R J. Application of the chlorophyll fluorescence induction dynamics in photosynthesis study [J]. Journal of Plant Physiology and MolecularⅡ Biology, 2005, 31(6): 559—566 [李鵬民, 高輝遠(yuǎn), R J Strasser. 快速葉綠素?zé)晒庹T導(dǎo)動(dòng)力學(xué)分析在光合作用研究中的應(yīng)用, 植物生理學(xué)與分子生物學(xué)學(xué)報(bào), 2005, 31(6): 559—566]
[41] Liu Y Y, Shao J H, Liu D M, et al. Effect of chelerythrine on the growth and photosynthetic system of Microcystis aeruginosa [J]. Acta Hydrobiologica Sinica, 2015, 39(1): 149—154 [劉彥彥, 邵繼海, 劉德明, 等. 白屈菜紅堿對(duì)銅綠微囊藻生長(zhǎng)和光合系統(tǒng)的影響, 水生生物學(xué)報(bào), 2015, 39(1): 149—154]
[42] Mulderij G, Mau B, Donk E, et al. Allelopathic activity of Stratiotes aloides on phytoplankton-towards identification of allelopathic substances [J]. Hydrobiologia, 2007, 584(1): 89—100
[43] Zhang T T, Zheng C Y, Hu W. The allelopathy and allelopathic mechanism of phenolic acids on toxic Microcystis aeruginosa [J]. Journal of Applied Phycology, 2010, 22(1): 71—77
[44] Liu G T, Zhou C F, Sun L F, et al. Effects of eichhornia crassipes allelochemicals on the growth of two mono-and co-cultured algae Microcystis aeruginosa and Scenedesmus obliquus [J]. Acta Scientiae Circumstantiae, 2011, 31(10): 2303—2311 [劉光濤, 周長(zhǎng)芳, 孫利芳, 等. 鳳眼蓮化感物質(zhì)對(duì)銅綠微囊藻、斜生柵藻生長(zhǎng)及細(xì)胞數(shù)相對(duì)比例的影響.環(huán)境科學(xué)學(xué)報(bào), 2011, 31(10): 2303—2311]
[45] Platt T, Gallegos C L, Harrison W G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton [J]. Journal of Marine Research, 1980, 38(4): 687—701 [46] Appenroth K J, St?ckel J, Srivastava A, et al. Multiple effects of chromate on the photosynthetic apparatus of Spirodela polyrhiza as probed by OJIP chlorophyll a fluorescence measurements [J]. Environmental Pollution, 2001, 115(1): 49—64
[47] Van Heerden P D R, Strasser R J, Krüger G H J. Reduction of dark chilling stress in N2-fixing soybean by nitrate as indicated by chlorophyll a fluorescence kinetics [J]. Physiologia Plantarum, 2004, 121(2): 239—249
[48] Asada K. Ascorbate peroxidase—a hydrogen peroxidescavenging enzyme in plants [J]. Physiologia Plantarum, 1992, 85(2): 235—241
[49] Drotar A, Phelps P, Fall R. Evidence for glutathione peroxidase activities in cultured plant cells [J]. Plant Science, 1985, 42(1): 35—40
[50] ACGIH (American Conference of Governmental Industrial Hygienists), Documentation of the TLVs and BEIs with other worldwide occupational exposure values. N-phenyl-beta- naphthylamine. ACGIH Worldwide, Cincinnati (CD-ROM). 2006
EFFECT OF N-PHENYL-2-NAPHTHYLAMINE ON THE GROWTH, ANTIOXIDATIVE ENZYME AND PSIN Ⅱ CYLINDROSPERMOPSIS RACIBORSKII
LIU Rui, BAI Fang, RAN Xiao-Fei, YANG Yan-Jun, YANG Song-Qi, SHI Jun-Qiong and WU Zhong-Xing
(Key Laboratory of Eco-environments in Three Gorges Reservoir Region (Ministry of Education), Chongqing Key Laboratory of Plant Ecology and Resources Research in Three Gorges Reservoir Region, School of Life Science, Southwest University, Chongqing 400715, China)
In this study, we investigated the effects of N-phenyl-2-naphthylamine, a type of allelochemical, on the invasive cyanobacteria Cylindrospermopsis raciborskii. We measured the growth, the activities of antioxidative enzymes, and polyphasic Chl. a fluorescence transients of Cylindrospermopsis raciborskii that was cultured in the presence of N-phenyl-2-naphthylamine at different concentrations (0, 0.05, 0.10, 0.50, 1.00 and 2.00 mg/L). When the concentration of N-phenyl-2-naphthylamine was higher than 0.50 mg/L, the Chl. a content was significantly lower than the control. This indicated that the level of Chl. a could be reduced by the increase in N-phenyl-2-naphthylamine. The EC50of N-phenyl-2-naphthylamine on C. raciborskii was 1.02 mg/L after a 72h exposure. At the concentration lower than 0.50 mg/L, N-phenyl-2-naphthylamine caused a significant increase in a variety of parameters of polyphasic Chl. a fluorescence transients, including the maximum quantum yield of primary photochemistry (φPo), the density of RCs(RC/CSo), the performance index on absorption basis (PIABS), and the performance index on cross section basis (PICS). One exception was ψo(hù)that declined in the presence of low concentration of N-phenyl-2-naphthylamine. On the other hand, φPo, RC/CSo, PIABS, PICS, and ψo(hù)were all reduced when the concentration of N-phenyl-2-naphthylamine was higher than 0.50 mg/L (eg. 1.00 mg/L and 2.00 mg/L). These data suggested that low dose of N-phenyl-2-naphthylamine might promote the photosynthetic efficiency of C. raciborskii, while high dose could reduce density of RCs and harm the PS Ⅱ and electron transfer, and thus result in the inhibition of photosynthetic efficiency in C. raciborskii. Compared to the control, the activities of superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX) were all obviously elevated along with the increase in N-phenyl-2-naphthylamine concentration. This implied that C. raciborskii might counter the damage of N-phenyl-2-naphthylamine with enhanced antioxidant enzymatic activities. In conclusion, N-phenyl-2-naphthylamine could inhibit the growth of C. raciborskii by disrupting the electron transfer of PS Ⅱ and lowering the density of RCs, and therefore it could be considered as a potential compound for the control of Cylindrospermopsis blooming.
Photosynthesis; Cylindrospermopsis raciborskii; Antioxidative system; Allelochemical compound
Q948.11
A
1000-3207(2015)04-0774-08
10.7541/2015.101
2014-06-30;
2014-09-18
中央高校基本科研業(yè)務(wù)費(fèi)專項(xiàng)資金項(xiàng)目(XDJK2014D015); 西南大學(xué)博士基金(SWU110065); 國(guó)家自然科學(xué)基金(31170372)資助
劉瑞, (1989—), 男, 湖北建始人; 碩士研究生; 主要從事藻類生態(tài)學(xué)研究。E-mail: 20081989liurui@163.com
吳忠興(1975— ), 男, 教授; 主要從事藻類生理生態(tài)及分子系統(tǒng)學(xué)研究。E-mail: wuzhx@swu.edu.cn
