弓形蟲ROP16I/IIIDNA疫苗不能誘導(dǎo)小鼠有效的保護(hù)性免疫
2015-02-14 08:07:24周芹芝羅慶禮沈繼龍
中國人獸共患病學(xué)報(bào) 2015年11期
周芹芝,李 曼,陳 鶴,都 建,羅慶禮,沈繼龍
弓形蟲ROP16I/IIIDNA疫苗不能誘導(dǎo)小鼠有效的保護(hù)性免疫
周芹芝1,2,李 曼1,陳 鶴3,都 建1,羅慶禮1,沈繼龍1
目的 利用ROP16I/III基因真核表達(dá)重組質(zhì)粒免疫BALB/c雌性小鼠,檢測小鼠體液免疫和細(xì)胞免疫,評估ROP16I/III作為潛在的分子疫苗的價(jià)值。方法 克隆Chinese 1型弓形蟲ROP16I/III, 將其插入真核表達(dá)載體pEGFP-C2中構(gòu)建重組質(zhì)粒pEGFP-ROP16I/III,脂質(zhì)體法轉(zhuǎn)染T293細(xì)胞并觀察體外表達(dá)。Western blotting分析鑒定。將36只SPF級小鼠隨機(jī)均分為:①PBS組;②空質(zhì)粒組;③pEGFP-ROP16I/III組。每2周肌注免疫1次(100 μg/只),共3次;于免疫前及每次肌注前1 d和末次免疫后2周收集小鼠血清,間接ELISA法檢測血清抗弓形蟲IgG。于末次免疫2周后取小鼠脾臟,分離淋巴細(xì)胞培養(yǎng)測細(xì)胞因子。剩余小鼠腹腔注射Chinese 1型弓形蟲 Wh3株速殖子1 000個(gè)/只,觀察小鼠攻擊感染后的存活時(shí)間和存活率。結(jié)果 真核表達(dá)載體構(gòu)建成功,體外見到重組質(zhì)粒pEGFP-ROP16I/III在T293細(xì)胞的表達(dá);pEGFP-ROP16I/III末次免疫后血清中IgG水平及脾淋巴細(xì)胞細(xì)胞因子均無明顯升高(P>0.05);攻擊感染后,免疫組小鼠比對照組的存活時(shí)間無延長(P>0.05);生物信息學(xué)分析發(fā)現(xiàn)ROP16I/III缺乏線性B細(xì)胞表位。結(jié)論 Chinese1 型弓形蟲的ROP16I/III由于其分泌的定位和結(jié)構(gòu)特點(diǎn),不能誘導(dǎo)小鼠產(chǎn)生有效的抗Wh3株攻擊感染的免疫保護(hù)。
弓形蟲;ROP16I/III;分子疫苗
弓形蟲病(toxoplasmosis)是一種人獸共患病,嚴(yán)重危害人類健康和畜牧業(yè)生產(chǎn),目前尚無特效藥物用來預(yù)防和治療弓形蟲感染。唯一用于綿羊接種的弓形蟲疫苗為熱敏感無毒株疫苗S48(ts-4)[1]。但是由于弓形蟲生活史各階段抗原成分復(fù)雜,目前尚無用于其他動(dòng)物和人體的有效分子疫苗。研制安全有效的重組疫苗長期以來是弓形蟲病防治的研究熱點(diǎn)之一。DNA疫苗接種是將編碼抗原的基因插入載體質(zhì)粒中構(gòu)成重組體,直接接種機(jī)體,在接種部位攝取表達(dá)疫苗抗原分子,從而誘導(dǎo)機(jī)體產(chǎn)生抗感染免疫的有效途徑。在DNA疫苗的接種途徑中,直接肌注法被認(rèn)為是目前最簡單有效的途徑之一。目前用于弓形蟲疫苗研究的蟲源性蛋白包括表膜蛋白SAGs、棒狀體蛋白R(shí)OPs,致密顆粒蛋白GRAs,和微線體蛋白MICs等[2-5]。ROPs是蟲體分泌型細(xì)胞器棒狀體(rhoptry)的分泌蛋白,除了ROP2和ROP8外,多數(shù)具有絲氨酸-蘇氨酸蛋白激酶活性,也是蟲株重要的毒力因子[6-7]。ROP16是由弓形蟲棒狀體分泌的一組與細(xì)胞內(nèi)增殖和調(diào)節(jié)有關(guān)的效應(yīng)分子,參與納蟲泡的形成,以抵抗宿主細(xì)胞內(nèi)的殺傷。有研究報(bào)告,ROP16用于小鼠的免疫可誘導(dǎo)抗感染的免疫力[8,9]。不同弓形蟲基因型具有ROP16的多態(tài)性。我國流行的優(yōu)勢弓形蟲基因型為Chinese 1[10-12],其ROP16的多態(tài)性為ROPI/III。為了深入探討ROP16的潛在疫苗應(yīng)用價(jià)值,本研究克隆了Chinese 1基因型Wh3 蟲株的ROP16I/III,并構(gòu)建了真核表達(dá)質(zhì)粒,對小鼠進(jìn)行了重組ROP16I/III(rROP16I/III)的免疫保護(hù)性的研究。現(xiàn)將結(jié)果報(bào)告如下。
1 材料與方法
1.1 蟲株、菌種、質(zhì)粒及細(xì)胞 弓形蟲Wh3 株(Chinese 1 型),大腸桿菌DH5α,真核表達(dá)載體pEGFP-C2,HEK293T(人胚腎細(xì)胞株)細(xì)胞均由本實(shí)驗(yàn)室保存。
1.2 實(shí)驗(yàn)動(dòng)物 SPF級BALB/c雌性小鼠購自北京維通利華實(shí)驗(yàn)動(dòng)物技術(shù)有限公司,經(jīng)安徽醫(yī)科大學(xué)試驗(yàn)動(dòng)物倫理委員會(huì)審批(No.2014012)。
1.3 主要試劑 質(zhì)粒提取試劑盒GoldHi EndoFree Plasmid Maxi Kit購自康為世紀(jì)公司(中國);HRP標(biāo)記的羊抗鼠IgG,IgG1和IgG2a購自Invitrogen公司;FBS(Gibco,美國);SDS-PAGE 凝膠配置試劑盒(碧云天,中國);預(yù)染蛋白markers 和ECL底物發(fā)光檢測試劑盒(Fermentas,美國);逆轉(zhuǎn)錄試劑盒(Thermo,美國);DMEM高糖培養(yǎng)基,轉(zhuǎn)染試劑Lipofectamine 2000,Opti-MEM(Gibco公司,美國);SYBR(Takara, 中國);AxyPrep DNA凝膠回收試劑盒與AxyPrep質(zhì)粒小量提取試劑盒(Axygen公司,中國);細(xì)胞因子(IL-2、IL-4、IL-10和IFN-γ)ELISA試劑盒購自北京四正柏生物科技有限公司。
1.4 真核表達(dá)重組質(zhì)粒pEGFP-ROP16I/III的構(gòu)建與鑒定 根據(jù)GenBank中弓形蟲ROP16I/III的表達(dá)序列設(shè)計(jì)引物:上游引物 P1:5′-GAAGATCTAT GAAAGTGACC ACGAAAGA-3′,劃線序列為BgⅢ酶切位點(diǎn);下游引物P2:5′-CGAGCTCCAT CCGATGTGAAGAAAG-3′, 劃線序列為SacI酶切位點(diǎn)。以提取的Wh3蟲株的基因組DNA為模板,進(jìn)行PCR擴(kuò)增。PCR產(chǎn)物電泳鑒定、回收和純化,并克隆至同樣經(jīng)Bgm和SacI雙酶切的真核表達(dá)載體pEGFP-C2上,得到的陽性重組克隆pEGFP-ROP16I/III進(jìn)行酶切、測序和鑒定。
1.5 pEGFP-ROP16I/III轉(zhuǎn)染及鑒定 鑒定正確后提取質(zhì)粒,轉(zhuǎn)染HEK293T細(xì)胞。24孔板培養(yǎng)HEK293T細(xì)胞,轉(zhuǎn)染前1 d換為無血清 DMEM 培養(yǎng)基, 在500 μL無抗生素的DMEM培養(yǎng)基中接種2×105細(xì)胞,待細(xì)胞生長至90%~95%密度時(shí)開始轉(zhuǎn)染,6 h后移去轉(zhuǎn)染液,加DMEM完全培養(yǎng)基。培養(yǎng)24 h后,在熒光顯微鏡下觀察綠色熒光,分析重組質(zhì)粒pEGFP-ROP16I/III在HEK293T細(xì)胞中的表達(dá)。
1.6 重組質(zhì)粒在免疫小鼠體內(nèi)表達(dá) 提取免疫局部的小鼠后腿肌肉組織RNA,逆轉(zhuǎn)錄成cDNA后PCR擴(kuò)增ROP16I/III目的片段,鑒定其在體內(nèi)的表達(dá)。
1.7 弓形蟲裂解抗原的制備 弓形蟲在HFF細(xì)胞中培養(yǎng)。待蟲體即將HFF細(xì)胞全部裂解時(shí),離心收集速殖子。將速殖子懸液置-20 ℃反復(fù)凍融3次,置于冰上超聲破碎。經(jīng)BCA蛋白濃度測定試劑盒對速殖子裂解液進(jìn)行蛋白質(zhì)濃度測定,-80 ℃保存?zhèn)溆谩?/p>
1.8 動(dòng)物分組及免疫 36只SPF級BALB/c雌性小鼠隨機(jī)分成3組,每組12只。 第1組為實(shí)驗(yàn)組(肌肉注射pEGFP-ROP16I/III);第2組為空質(zhì)粒對照組;第3組為PBS對照組。重組質(zhì)粒提取純化后用PBS稀釋至100 μg/100 μL。實(shí)驗(yàn)組小鼠經(jīng)肌肉注射100 μL/只, 空質(zhì)粒對照組和空白對照組用同劑量免疫,每2周肌肉注射免疫1次,共免疫3次。
1.9 免疫小鼠抗體測定 免疫前及每次免疫前1 d和末次免疫后2周,小鼠尾靜脈采血收集血清,間接ELISA檢測血清抗弓形蟲IgG。用弓形蟲 RH 株速殖子裂解液抗原包被96孔酶標(biāo)板4 ℃過夜, BSA 37 ℃封閉2 h。 鼠血清1∶50稀釋作為一抗, HRP標(biāo)記的抗小鼠IgG作為二抗,1 h后底物顯色,終止反應(yīng)后用酶標(biāo)儀測定各孔吸光度值。免疫鼠血清 OD值/陰性對照 OD>2為陽性。
1.10免疫小鼠細(xì)胞因子的測定 末次免疫2周后,每組取3只小鼠,分別制備脾細(xì)胞懸液,加入淋巴細(xì)胞分離液分離淋巴細(xì)胞,調(diào)整濃度接種于96孔培養(yǎng)板中,加入刺激物37 ℃,5%CO2細(xì)胞培養(yǎng)箱中培養(yǎng)。分別于培養(yǎng)24 h、72 h和96 h后收集細(xì)胞培養(yǎng)上清,置于-80 ℃保存,ELISA試劑盒檢測細(xì)胞因子。
1.11 攻擊感染實(shí)驗(yàn) 末次免疫兩周后,每只小鼠經(jīng)腹腔接種Wh3速殖子103個(gè),記錄一般狀況和存活時(shí)間。
1.12 統(tǒng)計(jì)學(xué)方法 實(shí)驗(yàn)數(shù)據(jù)用SPSS13.0軟件進(jìn)行統(tǒng)計(jì)分析,用T-test、ONE-WAY ANOVA分析,以α=0.05為水準(zhǔn),P<0.05具有統(tǒng)計(jì)學(xué)意義。
2 結(jié) 果
2.1 pEGFP-ROP16I/III表達(dá)載體的構(gòu)建與鑒定 以提取的重組質(zhì)粒pEGFP-ROP16I/III為模板進(jìn)行PCR擴(kuò)增,電泳后顯示一條分子量約2 124 bp的條帶,說明重組表達(dá)質(zhì)粒pEGFP-ROP16I/III初步構(gòu)建成功(圖1)。
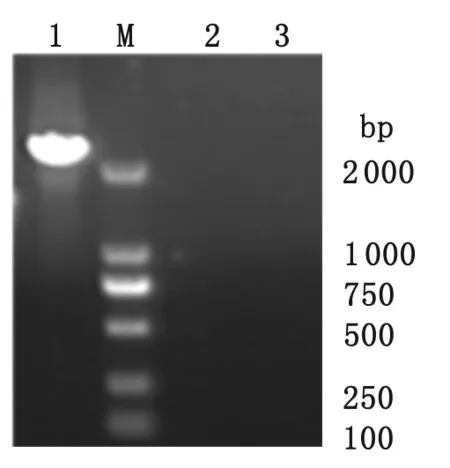
M: DNA分子量標(biāo)準(zhǔn)(bp);1 轉(zhuǎn)染重組質(zhì)粒pEGFP-ROP16I/III細(xì)胞組;2 轉(zhuǎn)染空質(zhì)粒pEGFP-C2組;3 空白對照組
M: molecular weight (bp); 1: cells transfected with recombinant pEGFP-ROP16I/III; 2: cells transfected with control plasmids; 3: cells without plasmids transfection.
圖1 重組載體pEGFP-ROP16I/III經(jīng)PCR擴(kuò)增的ROP16I/III片段
Fig.1 PCR amplification ofROP16I/IIIfrom template recombinant pEGFP-ROP16I/III.
2.2 pEGFP-ROP16I/III細(xì)胞內(nèi)表達(dá)的檢測 質(zhì)粒pEGFP-ROP16I/III體外轉(zhuǎn)染HEK293T細(xì)胞,培養(yǎng)24 h后,在熒光顯微鏡下見到綠色熒光,同時(shí)未轉(zhuǎn)染質(zhì)粒的HEK293T細(xì)胞觀察不到綠色熒光。雖然pEGFP-ROP16I/III轉(zhuǎn)染效率不及空白質(zhì)粒,但是與對照組相比,表明重組質(zhì)粒在真核細(xì)胞內(nèi)能有效表達(dá)(圖2)。
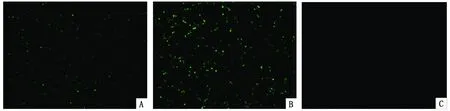
A:轉(zhuǎn)染重組質(zhì)粒pEGFP-ROP16I/III;B:轉(zhuǎn)染空質(zhì)粒pEGFP-C2;C,空白對照
A: cells transfected with recombinant pEGFP-ROP16I/III;B: cells transfected with control plasmids; C: cells without plasmids transfection.
圖2 重組質(zhì)粒pEGFP-ROP16I/III轉(zhuǎn)染HEK293T后目的基因表達(dá)的檢測
Fig.2 Fluorescent detection of theROP16I/IIIgene expression in transfected HEK293T cells
2.3 重組質(zhì)粒在免疫小鼠體內(nèi)的表達(dá) 提取小鼠后腿肌肉組織總RNA,逆轉(zhuǎn)錄成cDNA后PCR擴(kuò)增ROP16I/III目的片段。結(jié)果可見與目的片段大小(2 124 bp)位置一致的條帶(圖3),證明體內(nèi)有ROP16I/III的mRNA水平轉(zhuǎn)錄。
2.4ROP16I/III的生物信息學(xué)分析 Chinese 1 基因型蟲株的ROP16氨基酸序列不同于II型蟲株(ROP16II)而與I型/III型蟲株一致(ROP16I/III)。亦即,其ROP16I/III第503位氨基酸為亮氨酸(L)而非絲氨酸(S),見圖4 A。ROP16I/III的編碼序列采用Bepipred Server (http://www.cbs.dtu.dk/services/BepiPred/) 軟件進(jìn)行B細(xì)胞表位分析,結(jié)果見到,ROP16I/III缺乏線性B細(xì)胞表位(見圖4 B)。ROP16I/III的三維空間結(jié)構(gòu)模型見圖5。
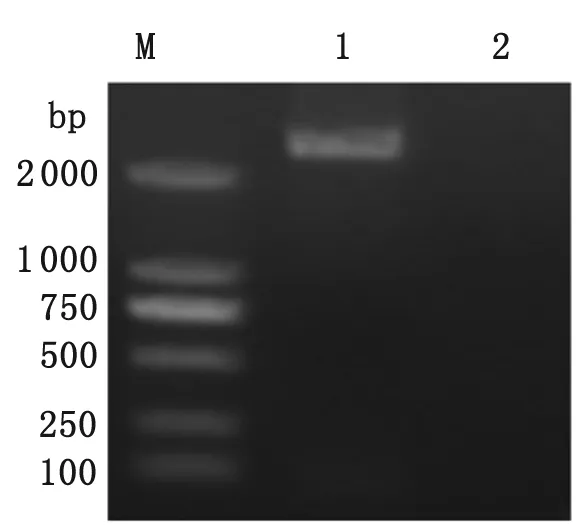
M:DNA分子量標(biāo)準(zhǔn)(bp);1:小鼠接種局部組織;2:遠(yuǎn)離接種局部的組織
M: DNA molecular weight(bp); 1: local tissues of vaccinated mouse; 2: adjacent tissues of the location of vaccination.
圖3 RT-PCR鑒定重組質(zhì)粒在小鼠肌肉組織表達(dá)
Fig.3 Identification ofROP16I/IIIexpression in the muscles of mouse by RT-PCR
2.5 免疫小鼠血清特異性抗ROP16I/IIIIgG抗體的檢測 最后一次免疫小鼠后,ELISA檢測小鼠血清IgG,如圖6所示:與對照組相比,實(shí)驗(yàn)組小鼠未能產(chǎn)生特異性抗體,在450nm處OD值未見明顯差異(P>0.05)。
檢測小鼠血清中IgG1和IgG2a的水平,如圖7所示,實(shí)驗(yàn)組和對照組的IgG1和IgG2a含量亦無顯著變化(P>0.05)。
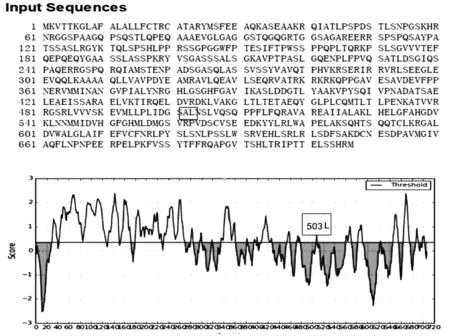
A:Chinese 1基因型弓形蟲ROP16多態(tài)性特征為ROP16I/III503L(第503位氨基酸為亮氨酸);B:ROP166I/III線性B細(xì)胞表位預(yù)測(示缺乏B細(xì)胞表位)
A: polymorphicROP16I/III,showing leucine (L) at amino acid 503; B: B epitope prediction ofROP16I/III, showing lack of B epitope in ROPI/III.
圖4ROP16I/III的生物信息分析
Fig.4 Bioinformatic analyses ofROP16I/III
2.6 免疫小鼠細(xì)胞免疫的檢測 免疫小鼠在末次免疫后取小鼠脾細(xì)胞,制備脾細(xì)胞懸液,加入相對應(yīng)的刺激物培養(yǎng),收集培養(yǎng)上清,經(jīng)ELISA檢測,發(fā)現(xiàn)試驗(yàn)組IL-2、IL-4、IL-10和IFN-γ與對照組之間無差異(表1),P>0.05,無統(tǒng)計(jì)學(xué)意義。
2.7 攻擊感染免疫保護(hù)效果 3組小鼠經(jīng)腹腔接種感染103個(gè)Wh3速殖子,均未能耐受攻擊感染,于第3 d開始,所有小鼠出現(xiàn)活動(dòng)減少、豎毛、弓背、飲食減少等表現(xiàn);攻擊后第8 d時(shí)3組小鼠均開始死亡;第10 d全部死亡。重組質(zhì)粒組未能延長小鼠壽命(P>0.05)(圖8)。說明ROP16I/III的DNA疫苗未能成功誘導(dǎo)機(jī)體產(chǎn)生抗攻擊感染的免疫保護(hù)作用。
圖5ROP16I/III的3D結(jié)構(gòu)(綠色箭頭所指為503L位點(diǎn))
Fig.5 Three dimensional structure ofROP16I/III, indicating leucine (L) at amino acid 503 (arrow)
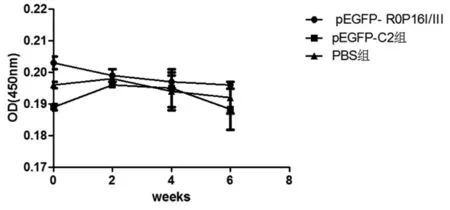
圖6 pEGFP-ROP16I/III質(zhì)粒免疫BALB/c小鼠血清抗ROP16I/III總IgG抗體
Fig.6 Total seral IgG antibodies againstROP16I/IIIin mice immunized with pEGFP-ROP16I/III
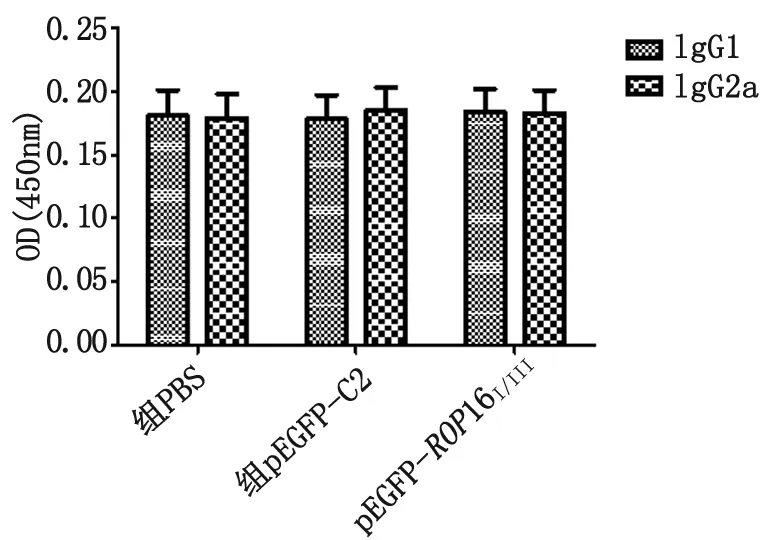
圖7 pEGFP-ROP16I/III質(zhì)粒免疫小鼠,血清抗體IgG1和IgG2a的檢測
Fig.7 IgG1 and IgG2 antibodies againstROP16I/IIIin mice immunized with pEGFP-ROP16I/III
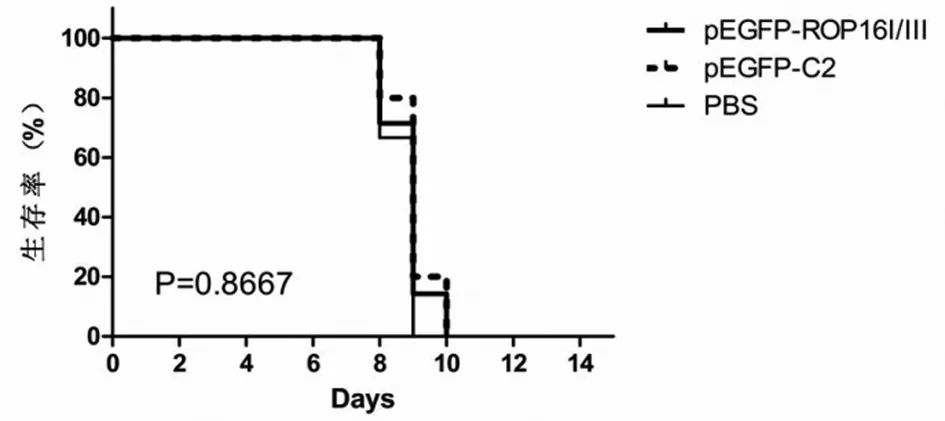
圖8ROP16I/IIIDNA免疫小鼠經(jīng)103個(gè)Wh3速殖子攻擊感染后的生存時(shí)間
Fig.8 Survival time of mice immunized with pEGFP-ROP16I/IIIfollowed by challenge with 103tachyzoites ofToxoplasmaWh3 virulent strain

表1 pEGFP-R0P16I/III質(zhì)粒免疫小鼠脾細(xì)胞分泌的細(xì)胞因子檢測(P>0.05)Tab.1 Cytokines detection in the splenocytes of mice immunized with pEGFP-ROP16I/III and control (P>0.05)
3 討 論
目前,唯一在歐洲和新西蘭注冊用于綿羊接種的弓形蟲疫苗為減毒活疫苗S48(熱敏蟲株)[13]。活蟲疫苗雖不能在體內(nèi)增殖,但是有轉(zhuǎn)變?yōu)榘业娘L(fēng)險(xiǎn)而不適用于人體。近年有關(guān)弓形蟲以及其他頂復(fù)原蟲(Apicomplexan)的表膜抗原和分泌抗原作為候選疫苗分子的研究較多。在蛋白質(zhì)疫苗中,有報(bào)告認(rèn)為[14],只有弓形蟲的全蟲裂解抗原(而非是若干重組抗原的混合物)才能誘導(dǎo)小鼠產(chǎn)生免疫保護(hù)力,表現(xiàn)為脾細(xì)胞高表達(dá)IFN-γ,巨噬細(xì)胞分泌高水平的iNOS和NO,小鼠腦內(nèi)包囊較少。
弓形蟲棒狀體(rhoptry)位于蟲體頂端復(fù)合器,一般含8~12個(gè)。蟲體入侵宿主細(xì)胞后,棒狀體分泌一系列具有絲氨酸/蘇氨酸蛋白激酶(serine-threonine kinase)活性的棒狀體蛋白(ROPs)。ROPs家族成員眾多,目前已經(jīng)鑒定出30多個(gè),其編碼基因位于弓形蟲第VIIa染色體上,在細(xì)胞入侵時(shí)蟲體分泌的ROPs與前端表膜融合,有助于蟲體入侵宿主細(xì)胞[15]。在宿主細(xì)胞內(nèi),ROPs主要定位于納蟲泡膜(parasitophorous vacuole membrane, PVM)[16],是弓形蟲調(diào)控宿主細(xì)胞的重要效應(yīng)分子,也是蟲體逃避巨噬細(xì)胞清除的重要機(jī)制。因此ROPs家族的鑒定和功能描述被認(rèn)為是弓形蟲生物學(xué)研究的一個(gè)重大突破[17]。Garcia 等曾試用弓形蟲棒狀體復(fù)合粗抗原接種家豬,雖然見到抗原可誘導(dǎo)豬對III型蟲株VEG卵囊感染的部分免疫保護(hù)(腦內(nèi)包囊減少),但是對急性期感染無保護(hù)作用。其原因可能是皮下接種抗原不能提供腸粘膜的免疫保護(hù)[18]。用相同抗原接種貓,結(jié)果3只貓中的2只排卵囊數(shù)目減少。但是由于動(dòng)物數(shù)量少同樣難以判斷疫苗的保護(hù)力[19]。棒狀體蛋白中的ROP2的DNA疫苗可在C3H小鼠提供部分免疫保護(hù),但在C57BL/6 和BALB/c 小鼠則未見有任何作用[20]。同樣的結(jié)果見于Leyva 的報(bào)道[21]。作者采用ROP2的DNA疫苗在3個(gè)小鼠品系(CBA/J, C57BL/6, BALB/c),進(jìn)行了測試。結(jié)果未見對于RH攻擊感染的任何保護(hù)力。
ROP16為ROPs家族的重要成員,被認(rèn)為是弓形蟲重要的毒力相關(guān)因子。弓形蟲接觸細(xì)胞后10分鐘,在受染細(xì)胞的細(xì)胞核內(nèi)便可以檢測到ROP16。弓形蟲在入侵宿主細(xì)胞時(shí),會(huì)分泌并釋放ROP16到宿主細(xì)胞質(zhì)中,然后ROP16被核定位信號(nuclear localization signals, NLS)運(yùn)輸至細(xì)胞核內(nèi),完成細(xì)胞核內(nèi)的定位并發(fā)揮功能,這在弓形蟲特異性蛋白中非常罕見[16-17]。
本研究克隆表達(dá)了流行我國的優(yōu)勢基因型 Chinese 1 弓形蟲Wh3株ROP16I/III編碼序列。以此構(gòu)建pEGFP-ROP16I/III重組質(zhì)粒,體外轉(zhuǎn)染HEK293T細(xì)胞見到目的蛋白的表達(dá);質(zhì)粒接種BALB/c后,檢測到ROP16I/III的mRNA在體內(nèi)的轉(zhuǎn)錄。給與免疫小鼠經(jīng)腹腔攻擊感染W(wǎng)h3株速殖子103個(gè)。結(jié)果見到,雖然接種和表達(dá)成功,但是免疫小鼠未出現(xiàn)明顯的體液和細(xì)胞介導(dǎo)的免疫應(yīng)答,且對攻擊感染缺乏有效的免疫保護(hù)。感染鼠的臨床表現(xiàn)與存活時(shí)間與免疫組相比未見顯著差異。
國內(nèi)有報(bào)告,用pVAX-ROP16質(zhì)粒免疫昆明小鼠,可以誘導(dǎo)出淋巴細(xì)胞的增殖、高滴度抗體、IL-2、IFN-γ、IL-4和IL-10水平升高,小鼠攻擊感染后存活時(shí)間延長,提示ROP16具有疫苗研究價(jià)值[8];用ROP16和GRA7構(gòu)建融合表達(dá)質(zhì)粒肌注免疫昆明小鼠,可誘導(dǎo)高滴度的IgG抗體和IFN-γ, CD8+細(xì)胞比例升高,小鼠存活時(shí)間延長[9]。然而有趣的是,Jensen 等應(yīng)用II型蟲株的ROP16轉(zhuǎn)基因弓形蟲(Strain II+ROP16I)免疫C57BL/6J小鼠,結(jié)果見到,只有內(nèi)源性的GRA15II和插入的ROP16I共同表達(dá)的基礎(chǔ)上,才能誘導(dǎo)小鼠對于經(jīng)口感染的免疫保護(hù)力,表現(xiàn)為小鼠產(chǎn)生了對1 000個(gè)PRU包囊和100個(gè)ME49株包囊攻擊感染后的100%的保護(hù)力[22]。最近, Alvarez 等[17]發(fā)現(xiàn),人體與動(dòng)物感染(肉類)的弓形蟲的ROP16具有顯著的多態(tài)性差異。前者多為強(qiáng)毒的ROP16(ROP16I);而后者全部為弱毒的ROP16(ROP16II)。強(qiáng)毒的ROP16I因?yàn)槿狈€性B細(xì)胞表位而失去免疫原性,難以(僅10.5%)誘導(dǎo)小鼠產(chǎn)生特異性抗體。本研究與該作者結(jié)果相一致。
本實(shí)驗(yàn)采用我國Chinese 1 型弓形蟲的多態(tài)性ROP16I/III的DNA疫苗免疫小鼠,雖然在體內(nèi)獲得了真核表達(dá)產(chǎn)物,但未見誘導(dǎo)小鼠的顯著性免疫保護(hù)力。其原因可能是:①ROP16主要是蟲體入侵后分泌在宿主細(xì)胞質(zhì)內(nèi),構(gòu)成納蟲泡膜的成分參與蟲體的免疫逃避和宿主的免疫調(diào)節(jié),難以暴露并有效刺激宿主產(chǎn)生抗蟲免疫;②毒力因子ROP16I/III缺乏線性B細(xì)胞表位,這一結(jié)構(gòu)特點(diǎn)難以誘導(dǎo)出高水平的特異性抗體;③相同疫苗在不同小鼠品系的結(jié)果差異較大,本實(shí)驗(yàn)采用了對弓形蟲高度敏感的BALB/c小鼠,結(jié)果與Vercammen的報(bào)告一致。為了真實(shí)地反應(yīng)出ROP16I/III的疫苗效果,本研究的質(zhì)粒構(gòu)建未采用細(xì)胞因子佐劑的融合表達(dá),攻擊感染采用了毒力較強(qiáng)的Wh3株而非是II型成囊株。由于我國流行的這一優(yōu)勢基因型弓形蟲Wh3蟲株兼有II型和I型的主要效應(yīng)分子多態(tài)性特點(diǎn),深入探討其他效應(yīng)分子的潛在疫苗價(jià)值仍然具有十分重要的意義。
4 結(jié) 論
我國流行的優(yōu)勢基因型Chinese 1的多態(tài)性ROP16I/III難以誘導(dǎo)實(shí)驗(yàn)小鼠產(chǎn)生高滴度抗體和Th1應(yīng)答,不能為BALB/c小鼠提供有效的免疫保護(hù)力。
[1]Katzer F,Canton G, Burrells A, et al. Immunization of lambs with the S48 strain ofToxoplasmagondiireduces tissue cyst burden following oral challenge with a complete strain of the parasite[J]. Vet Parasitol, 2014, 205(1/2): 46-56.
[2]Wang Y, Yin H. Research progress on surface antigen 1 (SAG1) ofToxoplasmagondii[J]. Parasit Vectors, 2014, 13(7): 180. DOI: 10. 1186/1756-3305-7-180
[3]Lu G, Wang L, Zhou A,et al. Epitope analysis, expression and protection of SAG5A vaccine againstToxoplasmagondii[J]. Acta Trop, 2015, 146: 66-72. DOI: 10.1016/j.actatropica.2015.03.013
[4]Hiszczynska-Sawicka E, Oledzka G, Holec-Gasior L, et al. Evaluation of immune responses in sheep induced by DNA immunization with genes encoding GRA1, GRA4, GRA6 and GRA7 antigens ofToxoplasmagondii[J]. Vet Parasitol, 2011, 177(3/4): 281-289.
[5]Lourenco EV, Bemardes ES, Silva NM, et al. Immunization with MIC1 and MIC4 induces protective immunity againstToxoplasmagondii[J]. Microbes Infect, 2006, 8(5): 1244-1251.
[6]Qiu W, Wernimont A, Tang K, et al. Novel structural and regulatory features of rhoptry secretory kinases inToxoplasmagondii[J]. EMBO J, 2009, 28 (7): 969-979.
[7]Xu Y, Gao Q, Xu QM, et al. New advances in studies ofToxoplasmagondiirhoptry neck proteins and rhoptry proteins[J]. Chin J Zoonoses, 2013, 12: 1217-1221. (in Chinese) 徐穎, 高琦, 徐前明, 等. 弓形蟲棒狀體頸部蛋白及棒狀體蛋白研究的新進(jìn)展[J]. 中國人獸共患病學(xué)報(bào), 2013, 12: 1217-1221.
[8]Yuan ZG, Zhang XX, He XH, et al. Protective immunity induced byToxoplasmagondiirhoptry protein 16 against toxoplasmosis in mice[J]. Clin Vaccine Immunol, 2011, 18(1): 119-124.
[9]Liu Q, Wang FW, Wang G, et al.Toxoplasmagondii: immune response and protective efficacy induced by ROP16/GRA7 multicomponent DNA vaccine with a genetic adjuvant B7-2[J]. Human Vaccines Immunotherapeutics, 2014, 10(1): 184-1912.
[10]Wang L, Chen H, Liu DH, et al. Genotypes and mouse virulence ofToxoplasmagondiiisolates from animals and humans in China[J]. PLoS ONE, 2013, 8(1): e53483.
[11]Wang L, Cheng HW, Huang KW, et al.Toxoplasmagondiiprevalence in food animals and rodents in different regions of China: isolation, genotyping and mouse pathogenicity[J]. Parasites Vectors, 2013, 6: 273.
[12]Li M, Mo XW, Wang L, et al. Phylogeny and virulence divergency analyses ofToxoplasmagondiiisolates from China[J]. Parasites Vectors, 2014, 7: 133.
[13]Burrells A, Benavides J, Canton G, et al. Vaccination of pigs with the S48 strain ofToxoplasmagondii-safer meat for human consumption[J]. Vet Res, 2015, 46(1): 47. DOI: 10.1186/s13567-015-0177-0
[14]Wagner A, Schabussova I, Ruttkowski B. Prime-boost vaccination withToxoplasmalysate antigen, but not with a mixture of recombinant protein antigens, leads to reduction of brain cyst formation in BALB/c mice[J]. PLoS One, 2015, 26(5): e0126334. DOI: 10.1371/journal.pone.0126334
[15]Taylor S, Barragan A, Su C, et al, A secreted serine-threonine Kinase determines virulence in the eukaryotic pathogenToxoplasmagondii[J]. Science, 2006, 314: 1776-1780. DOI: 10. 1126/science.1133643
[16]Gilbert LA, Ravindran S, Turetzky JM, et al.Toxoplasmagondiitargets a protein phosphatase 2C to the nuclei of infected host cells[J]. Eukaryotic Cell, 2007, 6(1): 73-83.
[17]Catalina A,De-la-Torre A,Vargas M, et al. Striking divergence inToxoplasmaROP16 nucleotide sequences from human and meat samples[J]. J Infect Dis, 2015, 211: 2006-2013.
[18]Garcia JL, Gennari SM, Navarro IT, et al. Partial protection against tissue cysts formation in pigs vaccinated with crude rhoptry proteins ofToxoplasmagondii[J]. Vet Parasitol, 2005, 129(3/4): 209-217.
[19]Garcia JL, Navarro IT, Biazzono L, et al. Protective activity against oocyst shedding in cats vaccinated with crude rhoptry proteins of theToxoplasmagondiiby the intranasal route[J]. Vet Parasitol, 2007, 145(3、4): 197-206.
[20]Vercammen M, Scorza T, Huygen K, et al. DNA vaccination with genes encodingToxoplasmagondiiantigens GRA1, GRA7, and ROP2 induces partially protective immunity against lethal challenge in mice[J]. Infect Immun, 2000, 68(1): 38-45.
[21]Leyva R, Herion P, Saavedra R, et al. Genetic immunization with plasmid DNA coding for the ROP2 protein ofToxoplasmagondii[J]. Parasitol Res, 2001, 87(1): 70-79.
[22]Jensen KD, Hu K, Hitmarsh RJ, et al.Toxoplasmagondiirhoptry 16 kinase promotes host resistance to oral infection and intestinal inflammation only in the context of the dense granule protein GRA15[J]. Infect Immun, 2013, 81(6): 2156-2167.
Recombinant eukaryotic plasmid ofROP16I/IIIfailed to provide efficient protection against toxoplasmosis in BALB/c mice
ZHOU Qin-zhi1,2,LI Man1,CHEN He3,DU Jian1,LUO Qing-li1,SHEN Ji-long1
(1.DepartmentofParasitology,AnhuiMedicalUniversity,Hefei230032,China;2.DepartmentofClinicalLaboratory,theCentralHospitalofCTCEGROUP,Hefei230023,China;3.DepartmentofBiochemistry,AnhuiMedicalUniversity,Hefei230032,China)
To construct the recombinant eukaryotic expression plasmids ofToxoplasmaROP16I/III, and to assess the immune protection in BALB/c mice induced byROP16I/IIIDNA vaccine,T.gondiiROP16I/IIIcoding gene obtained by PCR amplification was inserted into the eukaryotic expression vector pEGFP-C2 to generate recombinant pEGFP-ROP16I/III. T293 cells were transfected by Liposome methodinvitroand the fusion protein expression was identified by Western blotting. Thirty-six mice were randomly divided into 3 groups with 12 in each: PBS control group; empty plasmid group; and pEGFP-ROP16I/IIIgroup, respectively. Immunization was completed in each mouse by intramuscular injection once 2 weeks for 3 times. Sera at each time point were collected for detection of antibodies againstToxoplasmaby indirect ELISA. After two weeks of the last immunization, mouse splenocytes were harvested and cultured for tests of cytokines in the supernatant. The remaining mice were injected with 1 000 Wh3 tachyzoites for each, followed by observation of survival time and mortality of the animals. The recombinant eukaryotic expression vector of pEGFP-ROP16I/IIIwas successfully generated and efficient expressions were noted both in the transfected T293 cells and in the muscles in which the plasmid vaccine was given. All vaccinated mice, however, did not present the increased IgG antibody titers and cytokine levels after pEGFP-ROP16I/IIIimmunization. Correspondingly, no difference was seen of either survival or mortality of the animals compared with the control. Our conclusion is thatToxoplasmaROP16I/IIIDNA vaccine failed to provide BALB/c mice with efficient immune protection against virulent tachyzoites challenge due to its inaccessibility to host immune components and its lack of linear B epitope.
Toxoplasmagondii;ROP16I/III; DNA vaccine
Shen Ji-long, Email: shenjilong53@126.com
國家自然科學(xué)基金(No.81471983)資助;國家“973”計(jì)劃項(xiàng)目(No. 2010CB530001)資助
沈繼龍,Email: shenjilong53@126.com
1.安徽醫(yī)科大學(xué)病原生物學(xué)教研室,安徽省人獸共患病重點(diǎn)實(shí)驗(yàn)室,安徽病原生物學(xué)省級實(shí)驗(yàn)室,合肥 230032; 2.中鐵四局集團(tuán)中心醫(yī)院檢驗(yàn)科,合肥 230000; 3.安徽醫(yī)科大學(xué)第一附屬醫(yī)院,合肥 230032
10.3969/j.issn.1002-2694.2015.11.005
R382
A
1002-2694(2015)11-1010-07
2015-06-17;
2015-08-12

Supported by the Funded by the National Natural Science Foundation of China ( No. 81471983); the National Basic Research Program of China (973 Project, No.2010CB530001)
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫(yī)科大學(xué)學(xué)報(bào)(2021年1期)2021-02-07 01:06:36
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
中華養(yǎng)生保健(2020年4期)2020-11-16 01:31:40
中西醫(yī)結(jié)合肝病雜志(2020年2期)2020-10-27 02:18:50
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年4期)2014-02-02 02:44:59
